Глава II. Происхождение, родственные связи и эволюция водорослей
Палеонтологические данные о водорослях настолько скудны, что на основании их совершенно невозможно нарисовать сколько-нибудь стройную картину эволюции этих растений и для этого приходится пользоваться почти исключительно методом сравнения и сопоставления ныне живущих форм. При этом принимаются в расчет разнообразные признаки - как морфологические, так и физиологические. Так, большое значение в систематике водорослей имеют их пигменты, особенности строения и цикла развития, наличие или отсутствие подвижных жгутиковых стадий и т. п. Следует также напомнить, что водоросли - сборное понятие, объединяющее несколько самостоятельных типов растительного мира, и поэтому вопрос о происхождении водорослей должен ставиться раздельно по отношению к различным типам и далеко не для всех из них он может быть решен с одинаковой степенью достоверности.
Большинство исследователей полагает, что жизнь впервые возникла в воде и лишь впоследствии перешла на сушу. Примитивность строения многих водорослей и обитание их в водной среде дает поэтому основание утверждать, что водоросли - наиболее древние зеленые растения, от которых произошла вся наземная растительность. Иными словами, мы можем сказать, что первыми зелеными растениями на земле были водоросли. Поэтому, если ставить вопрос, от каких растений произошли водоросли в давно прошедшие эпохи, необходимо сразу же отметить, что это могли быть только какие-то бесцветные организмы, более простые по своему строению, чем современные водоросли. Однако никаких ископаемых остатков их не сохранилось.
Каковы же были эти первичные организмы и какая форма обмена веществ была им свойственна? По этому вопросу существуют две основные точки зрения. Первая по времени точка зрения сводится к тому, что первичные организмы должны были обладать способностью строить вещества своего тела из неорганических веществ, получаемых из окружающей среды (автотрофное питание). Необходимую для такого синтеза энергию они могли черпать из производимых ими химических реакций (хемоавтотрофное питание), так как световая энергия не могла быть используема из-за отсутствия соответствующих пигментов, как это имеет место и у современных хемосинтезирующих бактерий. Такое воззрение основывается на предположении, что в самом начале жизни на земле не было изобилия органических веществ, которыми могли бы питаться первые бесцветные организмы.
Согласно другой точке зрения, разработанной главным образом акад. А. И. Опариным, появлению первых организмов на земле, наоборот, должно было предшествовать образование массы органических веществ, за счет которых они и питались (гетеротрофное питание). Последнее воззрение является во многих отношениях более обоснованным и правильным. Можно полагать, что от таких первичных бесцветных гетеротрофов произошли, с одной стороны, многие бактерии* - в преобладающем большинстве также бесцветные и не способные к самостоятельному синтезу органических веществ, - а, с другой - зеленые растения, в первую очередь, водоросли.
* (Тип бактерий в целом, вероятно, представляет собой разнородную по своему происхождению группу. Некоторые бактерии (тиобактерии) скорее всего произошли от синезеленых водорослей в результате вторичной утери ими пигмента.)
Не подлежит сомнению, что хлорофилл и сопровождающие его пигменты, равно как и процесс фотосинтеза в той форме, в которой мы находим его у современных водорослей, настолько сложны, что их появлению должен был предшествовать длительный этап эволюции. О возможных путях этой эволюции известное представление может дать знакомство с некоторыми бактериями.
Как известно, среди ныне живущих бактерий, наряду с бесцветными, есть и окрашенные формы. У большинства из них вырабатываемые ими пигменты выделяются из клетки наружу и не участвуют в отправлении основных физиологических функций. Многие из этих пигментных бактерий питаются гетеротрофно. Однако существуют и такие, у которых пигмент остается внутри клеток, будучи связан с их протоплазмой. Очевидно, что в последнем случае отношение этих бактерий к свету должно быть иным, чем у бесцветных форм, так как свет, поглощаемый пигментами, может вызывать в клетке различные фотохимические реакции. И в настоящее время установлено, что у пурпурных и зеленых бактерий имеет место процесс фотосинтеза, который протекает несколько иначе чем у содержащих хлорофилл зеленых растений.
Хлорофилл, характерный для подавляющего большинства водорослей разных типов, представляет собой пигмент, переводящий световую энергию в химическую. На основании сказанного выше, можно думать, что формы, переходные между первичными бесцветными гетеротрофами и первичными зелеными фотосинтетиками, первоначально питались готовыми органическими веществами. Лишь впоследствии, благодаря выработке пигмента, остающегося в клетке, и связанной с этим возможности использовать световую энергию для синтетических процессов, гетеротрофное питание могло заместиться фототрофным - питанием органическими веществами, синтезируемыми из минеральных за счет световой энергии солнца. Так возникла и в дальнейшем все более совершенствовалась та форма питания, которая определила собой основное направление эволюции растительного мира на основе принципиально нового отношения к факторам внешней среды.
Происшедшие от первичных бесцветных гетеротрофов бактерии оказались слепой ветвью в развитии, не приведшей к появлению сколько-нибудь сложно построенных форм, но отличающейся, однако, большим разнообразием физиологических отправлений*. Напротив, первичные водоросли - как могут быть названы первые зеленые фотосинтетики - дали начало всему многообразию водорослей, явившихся, в свою очередь, родоначальниками высших растений. Как же могли произойти от них современные тины водорослей и какова связь между ними?
* (Способность некоторых бактерий к хемоавтотрофному питанию представляет собой скорее всего вторичное явление и характеризует боковое ответвление главного пути эволюции органического мира (акад. А. И. Опарин).)
Мы уже указывали, что в ряду признаков, кладущихся в основу системы водорослей, большое значение имеет наличие или отсутствие жгутиковых стадий. Большинство водорослей в цикле своего развития имеет подобные стадии или явно имело их в прошлом и утратило вторично. Однако известны два типа водорослей - синезеленые (Cyanophyta) и красные или багряные (Rhodophyta) - в отношении которых есть все основания полагать, что подобные стадии отсутствуют у них первично. У красных водорослей эта их особенность сочетается с весьма своеобразным половым процессом и - у многих форм - со сложной дифференциацией их талломов. С другой стороны, у синезеленых водорослей мы находим также и ряд других признаков, указывающих на их примитивность: просто построенное тело и - что особенно важно - полное отсутствие полового процесса и морфологически оформленных ядер и хроматофоров. Все эти особенности, несомненно, являются здесь первичными и свидетельствуют о значительной древности этого типа, не давшего, так же как и бактерии, сколько-нибудь сложных форм*. Древность происхождения синезеленых водорослей подтверждается и палеонтологическими данными (достоверные ископаемые остатки их обнаружены в верхнегуронских пластах протерозойской эры). Можно думать, что синезеленые и являются сохранившимися до наших дней сравнительно мало изменившимися потомками первичных водорослей.
* (Объединение синезеленых водорослей с бактериями в один тип дробянок (Schizophyta) на основании, главным образом, отсутствия ясной дифференциации протопласта на цитоплазму и ядро, не может считаться обоснованным, так как между ними существуют важные различия. Строение клетки тех и других далеко не одинаково. Типичные бактерии имеют жгутиковые стадии. Есть и ряд других существенных различий. Сказанное не исключает того, что некоторые бактерии могли произойти непосредственно от сине зеленых.)
Далеко не ясен вопрос о происхождении красных водорослей. Кроме отсутствия в цикле их развития жгутиковой стадии, с синезелеными водорослями их сближает сходство набора пигментов, сходное строение плазмодесм и некоторые другие, главным образом, биохимические признаки (запасный углевод гликоген синезеленых, повидимому, родственен "крахмалу багрянок" красных; слизь, образуемая некоторыми синезелеными, кажется химически иногда довольно близкой к слизи красных). С другой стороны, наличие в их клетках типичных ядер и хроматофоров*, сложный оогамный половой процесс и морфологическое расчленение слоевища у высших представителей говорит как будто против непосредственной связи этого типа с синезелеными водорослями, хотя исследования последнего времени показали наличие значительного морфологического сходства между некоторыми примитивными представителями порядка гониотриховых из красных водорослей (Asterocytis, Goniotrichum) и порядка тубиелловых из синезеленых водорослей (Tubiella, Cyanothrix).
* (Интересно, однако, что запасные углеводы образуются здесь не в хроматофорах (как это обычно характерно для зеленых водорослей), а в цитоплазме, обыкновенно у краев хроматофоров, что, может быть, указывает на известную примитивность этих органоидов у красных водорослей.)
В настоящее время, повидимому, правильнее всего считать, что красные водоросли имели когда-то общего предка с синезелеными, строение которого в настоящее время обрисовать невозможно, но с тех пор проделали длинный путь эволюции, являясь весьма древней группой (ископаемые остатки известны с кембрийского периода палеозоя), не давшей переходов к другим растениям.
Возвращаясь к нашей исходной группе первичных водорослей, мы должны, следовательно, отметить, что, дав боковую ветвь к синезеленым и красным водорослям, она свое основное развитие проделала по другому пути. Очевидно, что в боковой ветви, несмотря на все морфологическое разнообразие составляющих ее водорослей, не появилось таких форм, которые в эволюционном отношении были бы прогрессивными. Такой эволюционно-прогрессивной формой оказалось строение растения в виде хлорофиллоносного подвижного жгутикового организма (монадная структура), возникновение которого знаменовало собой весьма важный этап в эволюции растительного мира, так как все другие типы водорослей (кроме синезеленых и красных) прямо или косвенно произошли от жгутиконосных предков. Последние, таким образом, явились как бы центром расхождения для многообразных водорослей. При этом нужно только помнить, что современные организмы монадного строения сами являются продуктом длительной эволюции и, несомненно, построены много сложнее чем те первичные жгутиконосцы, которые явились предками большинства водорослей.
Для некоторых типов (эвгленовые) до сих пор, за немногими исключениями, известна только монадная организация - за пределы ее в своей эволюции они не вышли. Однако другие типы выработали и иные структуры, основные формы которых были рассмотрены нами в главе I под именем ступеней морфологической дифференциации тела водорослей.
Усложнение строения и переход к многоклеточности, сначала в его простейшей форме нитчатого слоевища, а потом и в более сложных проявлениях, - были связаны с утерей подвижности в вегетативном состоянии. Исходная монадная структура во многих случаях сохранилась только у репродуктивных клеток (зооспоры, гаметы), а иногда исчезла и здесь (замена зооспор апданоспорами; переход от копуляции подвижных гамет к конъюгации протопластов вегетативных клеток). Однако именно наличие подвижных стадий у многих неподвижных в вегетативном состоянии водорослей и служит доказательством их происхождения от флагеллатообразных предков.
В высшей степени замечателен тот факт, что подвижные репродуктивные клетки водорослей, относящихся к разным типам, воспроизводят основные признаки, присущие одноклеточным их представителям, нормально характеризующимся монадной структурой. Так, зооспоры нитчатых зеленых крайне схожи с одноклеточными подвижными зелеными водорослями, зооспоры различных желтозеленых водорослей своими двумя неравными жгутами и окраской имитируют монадных одноклеточных представителей этого типа, зооспоры нитчатых золотистых водорослей весьма напоминают монадных Chrysophyta (рис. 65) и т. п.
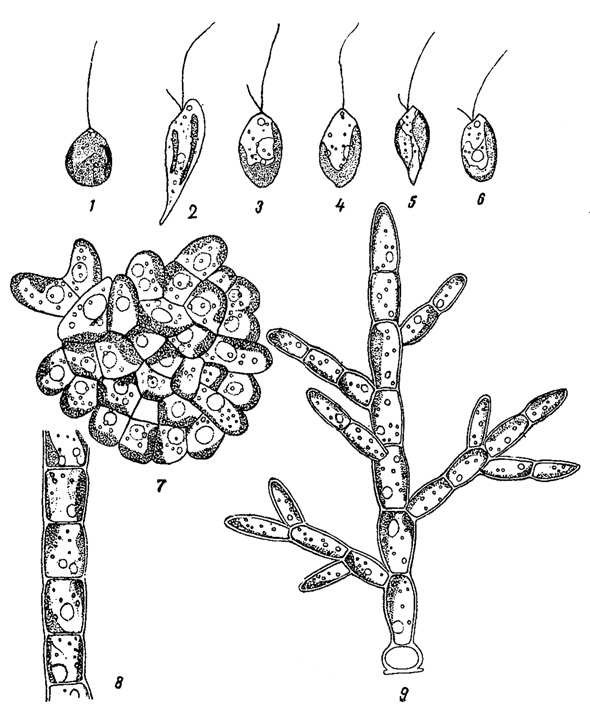
Рис. 65. Сходство одноклеточных водорослей монадной структуры с зооспорами водорослей нитчатой структуры в типе золотистых водорослей. 1-3 - водоросли монадной структуры: 1 - Chromulina, 2 - Ochromonas, 3 - Ochromonas. 4-6 - зооспоры водорослей нитчатой структуры: 4 - Phaeodermatium, 5 - Nematochrysis, 6 - Phaeothamnion. 7-9 - нитчатые слоевища этих водорослей: 7 - Phaeodermatium, 8 - Nematochrysis, 9 - Phaeothamnion
Это указывает на то, что эти типы произошли от разных жгутиконосцев, характерные особенности которых и до сих пор сохранились у репродуктивных клеток водорослей, утративших в вегетативном состоянии свою подвижность. В процессе их эволюции выработались разнообразные новые структуры, в то время как подвижная стадия все более сокращалась, сохраняя, однако, черты родоначальной формы.
Интересно далее, что сопоставление различных типов показывает известный параллелизм в их эволюции, проявляющийся в наличии у них схожих ступеней морфологической дифференциации. Так, у зеленых водорослей мы встречаем все отмеченные выше основные ступени кроме ризоподиальной. У желтозеленых водорослей они также налицо, включая и последнюю, но отсутствует пластинчатое и гетеротрихальное строение. У золотистых водорослей известны формы структуры амебоидной, монадной, пальмеллоидной и нитчатой. У пирофитовых водорослей - ризоподиальной, монадной, пальмеллоидной, коккоидной и нитчатой. Следует только иметь в виду, что простое перечисление наличных структур еще не характеризует в достаточной мере морфологические особенности данного типа, так как для разных типов преимущественно характерным оказывается различное строение. Так, нитчатая ступень особенно отчетливо выражена в типах зеленых и бурых, слабее у желтозеленых и еще более слабо у золотистых и пирофитовых водорослей, большинство представителей которых характеризуется монадной структурой.
А. Пашер (A. Pascher, 1931), работами которого, главным образом, и обосновано представление о происхождении типов водорослей от разных групп окрашенных жгутиковых, предложил ряд наименований систематических единиц (классов, подклассов, порядков), из которых сразу ясно, какая ступень морфологической дифференциации им свойственна. Для этого при обозначении этих категорий в пределах различных типов применяются одни и те же окончания или приставки, а именно: окончание - "monadineae" ("monadeae", "monadales") выражает монадную организацию, "capsineae" ("capsales") - пальмеллоидную, "coccineae" ("coccales") - коккоидную, Urichineae" ("trichales") - нитчатую, "siphoneae" ("siphonales") - сифональную; амебоидная структура передается приставкой "Rhizo". Так, например, в типе золотистых водорослей имеются классы Chrysomonadineaey Rhizochrysidineae, Chrysocapsineae, Chrysotrichi- neae; в типе желтозелепых водорослей - классы Rhizochloridineae, Heterocapsineae,Heterococcineaef Heterotrichineae, Heterosiphoneae и т. п.
Происхождение разных типов водорослей от различных жгутиконосных предков означает, что родоначальные формы у них неодинаковы. Это не исключает, конечно, того, что последние, в свою очередь, могли произойти от одного общего корня, но никакими сведениями об этом мы не располагаем. Однако есть основания считать, что некоторые типы находятся друг с другом и в прямой связи. Среди водорослей мы встречаем ряд типов, происхождение которых, несомненно, независимо, но, наряду с этим, некоторые типы, повидимому, филогенетически связаны между собой. Для того, чтобы выяснить это, следует остановиться на происхождении каждого типа в отдельности.
Происхождение золотистых водорослей (Chrysophyta) не вызывает сомнений. У них, как мы уже указывали, преобладает монадное строение, и несомненно, что подобные золотистые первичные монады и явились родоначальником всего этого типа в целом.
К золотистым водорослям близки диатомовые (Bacillariophyta) и желтозеленые или разножгутиковые водоросли (Xanthophyta или Heterocontae). Сходство между ними настолько значительно, что А. Пашер (1914, 1921) объединяет даже все эти три группы в одну единицу более высокого ранга, которой и присваивает наименование Chrysophyta, трактуемое им, следовательно, шире, чем это принято в настоящем "Определителе". Следующие главнейшие признаки являются для них общими. 1) Сходство в строении оболочек. Последние всегда (диатомовые) или очень часто (желтозеленые; цисты золотистых водорослей) состоят из двух равных или неравных половинок; а иногда слагаются из множества вдвинутых друг в друга отдельных частей или слоев (Tribonema, Ophiocytium желтозеленых; панцыри некоторых диатомовых, в которых при росте образуются вставочные ободки; домики ряда хризомонад, рис. 66). Кроме того, в оболочках у представителей всех трех типов откладывается кремнезем. 2) Близость пигментов. 3) Отсутствие в клетках крахмала и наличие, в качестве продуктов ассимиляции, жира и лейкозина (особенно характерного для золотистых, но найденного и у некоторых диатомовых и желтозеленых водорослей). 4) Сходство цист хризомонад с так называемыми покоящимися спорами (гипноспорами) некоторых диатомовых (Chaetoceros) и с цистами некоторых желтозеленых водорослей (Chloromeson, рис. 67). Важно также, что типичное для желтозеленых водорослей наличие у подвижных клеток двух неравных жгутов наблюдается и у многих хризомонад (порядок охромонадовых).
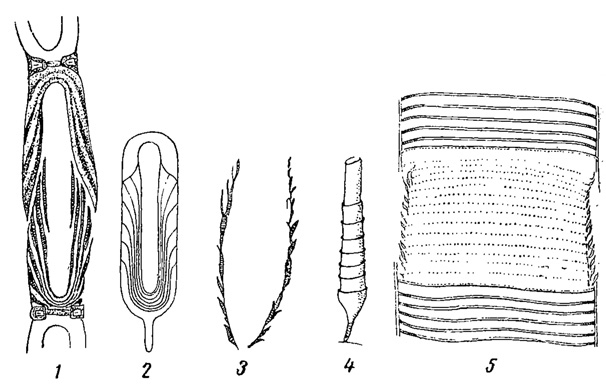
Рис. 66. Сходство в строении клеточных оболочек у желтозеленых, золотистых и диатомовых водорослей. 1 - клетка желтозеленой водоросли Tribonema. 2 - схема строения оболочки желтозеленой водоросли Ophiocytium. 3 - домик золотистой водоросли Dinobryon utriculus. 4 - строение домика золотистой водоросли Hyalobryon Voigtii. 5 - часть панцыря диатомовой водоросли Rhabdonema с вставочными ободками
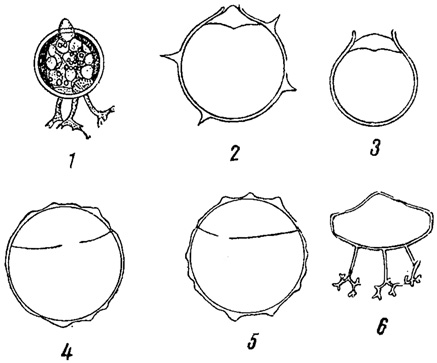
Рис. 67. Сходство покоящихся стадий желтозеленых, золотистых и диатомовых водорослей. 1-3 - цисты хризомонад (из золотистых водорослей). 4, 5 - цисты желтозеленой водоросли Chloromeson. 6 - покоящаяся спора диатомовой водоросли Chaetoceros
Для диатомовых характерна коккоидная структура. Однако у некоторых представителей класса Centricae известны зооспоры*, а в вегетативном состоянии их протопласты иногда имеют пульсирующие вакуоли (Attheya, Rhizosolenia). Это говорит в пользу происхождения диатомовых водорослей от предков монадного строения, каковыми, вероятно, и являлись формы, близкие к хризомонадам, так что палеонтологически сравнительно молодой тип Bacillariophyta (достоверные ископаемые остатки известны лишь начиная с юрского периода) представляет собой скорее всего боковую ветвь золотистых водорослей. Что же касается желтозеленых, то их происхождение от жгутиконосных предков не вызывает сомнений, а отмеченная только что близость к диатомовым и золотистым водорослям указывает на возможность наличия у всех этих трех типов общих родоначальных форм.
* (См. подстрочное примечание выше.)
У пирофитовых водорослей (Pyrrophyta), как отмечено выше, преобладает монадная структура. Этот тип занимает среди водорослей довольно изолированное положение и имеет или независимое происхождение или представляет собой группу, некогда обособившуюся от золотистых водорослей.
Не вполне ясно происхождение бурых водорослей (Phaeophyta), к которым относятся исключительно только многоклеточные формы, имеющие в цикле развития жгутиковую стадию. Филогенетически они, повидимому, связаны с какими-то организмами монадного строения, окрашенными в бурый цвет и, вероятно, близкими к хризомонадам. Возможно, что начало им дали более сложные колонильно-нитчатые хризомонады, уже потерявшие подвижность в вегетативном состоянии. Значительная сложность структуры, присущая многим представителям этого типа, указывает на то, что они являются продуктом длительной и своеобразной эволюции и представляют собой довольно древнюю группу (окаменелые остатки бурых водорослей известны, повидимому, со времен силурийского периода палеозойской эры).
Совершенно независимы в своем происхождении от рассмотренных типов зеленые водоросли (Chlorophyta), ведущие свое начало от зеленых жгутиконосных предков и дающие особенно ясную и полную картину постеленного перехода к более сложным формам, Этот тип относится к числу древнейших, так как уже в силурийских пластах обнаружены сложно построенные его представители (сифональной структуры), тогда как окаменелые остатки более простых форм неизвестны. Зеленые водоросли явились, повидимому, родоначальниками высшей наземной растительности. В пределах этого типа эволюция протекала по разным направлениям и привела к формам различной структуры. В подтипе Conjugatae имела место полная утеря подвижных репродуктивных клеток и переход к конъюгации. Этот подтип представляет собой боковую ветвь эволюционного ствола зеленых водорослей, подобно тому, как диатомовые являются боковой и еще более обособившейся ветвью золотистых водорослей.
Так же, как и пирофитовые, изолированное место среди водорослей занимает тип эвгленовых водорослей (Euglenophyta) - морфологически сравнительно однообразный, но характеризующийся довольно сложным и специализированным строением клеток. Возможно, что эвгленовые некогда произошли от примитивных зеленых водорослей монадной структуры, причем промежуточные формы вымерли.
Трудно сказать что-либо определенное о происхождении харовых водорослей (Charophyta), представляющих собой резко очерченную группу, не обнаруживающую отчетливых филогенетических связей с другими типами. Очевидно, это - боковой ствол зеленых водорослей, давно уже обособившийся от них (окаменелые остатки харовых известны с девонского периода палеозойской эры), причем связующие звенья не сохранились.
Мы видим, таким образом, что вопрос о происхождении водорослей выяснен далеко не полностью и в решении его существует значительное разнообразие точек зрения. То же относится и к системе водорослей. В настоящее время нет единства взглядов на систему водорослей и разные авторы по-разному делят их на таксономические единицы. Принимаемые нами типы водорослей перечислены и кратко охарактеризованы выше. Дальнейшие систематические подразделения этих типов будут приведены в соответствующих выпусках "Определителя".
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'