5. Размножение и цикл развития водорослей
Все разнообразные формы размножения водорослей могут быть сведены к трем основным типам - вегетативному, бесполому и половому. Вегетативное размножение - размножение отделившимися частями тела (многоклеточными или одноклеточными), обычно не специализированными в связи с репродуктивной функцией. Бесполое размножение - размножение с помощью выделения особых клеток (спор), специальной функцией которых является именно этот процесс. Наконец, половое размножение - размножение, осуществляемое также специализированными клетками (гаметами), но обычно сопровождаемое половым процессом (оплодотворением). Таким образом, при двух последних типах (бесполом и половом) размножение всегда происходит посредством отдельных клеток, тогда как при первом (вегетативном) оно часто осуществляется с помощью многоклеточных образований.
Во всех трех случаях размножение может быть связано с выработкой особых покоящихся стадий, приспособленных к перенесению неблагоприятных условий. При половом размножении эта связь в большинстве случаев налицо, при вегетативном и бесполом - она наблюдается далеко не всегда. С другой стороны, образование подобных стадий может иметь место и вне прямой зависимости от размножения. Так, отдельные клетки некоторых нитчатых водорослей (например, Ulothrix из зеленых водорослей) иногда одеваются толстыми оболочками, часто более, или менее округляются, в их протопластах откладывается большое количество запасных веществ (масло, крахмал), а иногда и пигмент, и в таком виде они переносят условия, в которых обычные вегетативные части погибают (рис. 40). Подобные клетки, называемые акинетами, представляют собой, таким образом, покоящиеся стадии. В дальнейшем, при благоприятных условиях, они могут снова, без распадения нити, начать нормально развиваться (например, у Cladophora из зеленых водорослей) и в этом случае образование акинет не связано с размножением. Однако часто нить распадается на отдельные акинеты и этот последний процесс является уже вегетативным размножением. Акинетам вполне подобны так называемые споры (покоящиеся клетки) у многих синезеленых водорослей (рис. 41).
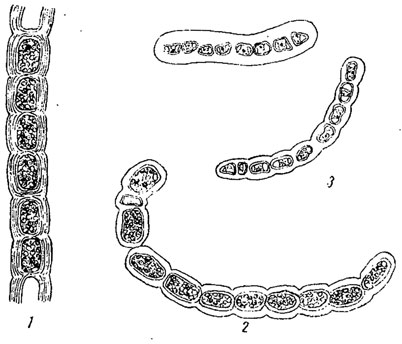
Рис. 40. Акинеты у разных видов Ulothrix. 1 - Ulothrix sp. 2 - U. tenerrima. 3 - U. subtilissima
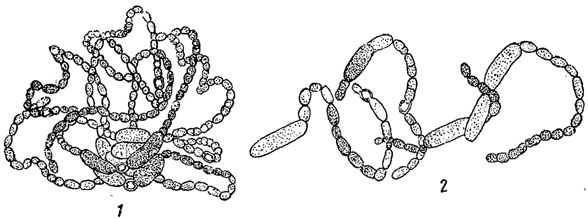
Рис. 41. Споры у синезеленых водорослей. 1 - Anabaena Lemmermannii. 2 - A. flos-aquae
Другой формой приспособления к перенесению неблагоприятных условий, обычно не связанной здесь непосредственно с размножением, является процесс инцистирования, наблюдаемый у многих водорослей монадной структуры. При подсыхании окружающей среды многие из них окружаются особой твердой оболочкой - цистой, внутри которой сохраняют свою жизнеспособность; при наступлении благоприятных условий существования оболочка сбрасывается и организм снова переходит в деятельное состояние (рис. 10).
Наконец, у некоторых как подвижных, так и неподвижных водорослей одной из форм покоящейся стадии является пальмеллевидное состояние, т. е. временный переход к пальмеллоидной структуре. Этот процесс часто сочетается с вегетативным размножением, так как окруженные слизью клетки могут интенсивно делиться (рис. 28). На других покоящихся стадиях у водорослей, образование которых связано с размножением, мы остановимся ниже, при характеристике последнего.
Вегетативное размножение у одноклеточных водорослей, естественно, осуществляется с помощью, отдельных клеток; у многоклеточных форм, оно в большинстве случаев происходит посредством отделения многоклеточных участков их тела. Простейшей формой вегетативного размножения, присущей одноклеточным водорослям амебоидной, монадной, пальмеллоидной и коккоидной структуры, является простое деление надвое. У форм монадного строения оно совершается или в подвижном или в неподвижном состоянии. В результате из одной клетки получаются две, подобные друг другу, которые или сразу расходятся или сосуществуют вместе, образуя колонию - временную или постоянную (рис. 42). Деление клеток внутри колонии приводит или к росту колонии или к образованию новой колонии, отделяющейся от материнской. В протекании всех этих процессов в разных группах водорослей существует большое разнообразие.
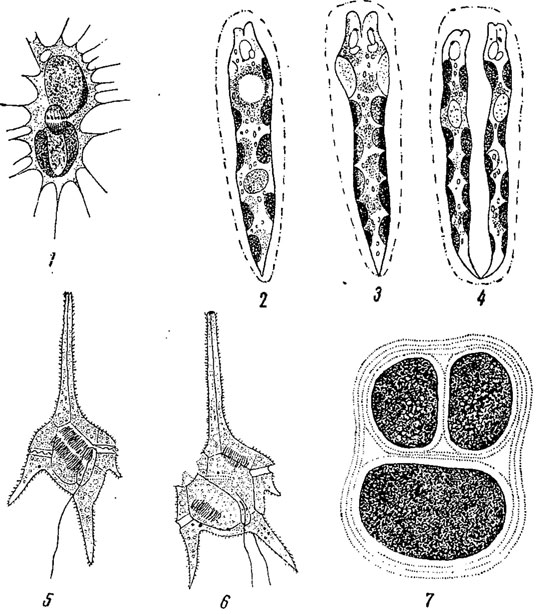
Рис. 42. Деление на-двое у разных водорослей. 1 - золотистая водоросль Rhizochrysis Scherfelii. 2-4 - эвгленовая водоросль Euglena deses. 5-6 - пирофитовая водоросль Ceratium hirundinella. 7 - синезеленая водоросль Gloeocapsa gigantea
У многоклеточных форм простейшим способом вегетативного размножения является фрагментация - распадение тела на отдельные части, из которых каждая впоследствии дорастает до нормальных размеров. Фрагментация наблюдается у многих нитчатых водорослей как зеленых (например, в порядке зигнемовых), так и, особенно, синезеленых (класс Hormogonecie). У последних отдельные фрагменты носят название гормогониев (рис. 43). Если нить, образующая гормогонии, имеет слизистый футляр, то гормогонии выскальзывают из него. Если слизистого футляра нет, нить просто распадается на гормогонии. Последние, таким образом, всегда лишены слизистого футляра. Гормогонии обладают способностью к активному движению в воде или во влажной среде, обусловленному выделением из клеток слизи.
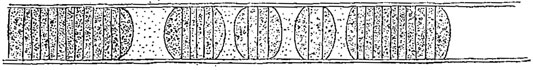
Рис. 43. Образование гормогониев у синезеленой водоросли Lyngbya Birgei
Сочетание функции размножения с приспособлением к перенесению неблагоприятных условий характерно для гормоспор или гормоцист, встречающихся также у некоторых синезеленых водорослей (в порядках мастигокладовых, диплонемовых и стигонемовых) и, в отличие от гормогониев, одетых толстой оболочкой, представляющей собой разросшийся слизистый футляр (рис. 44).
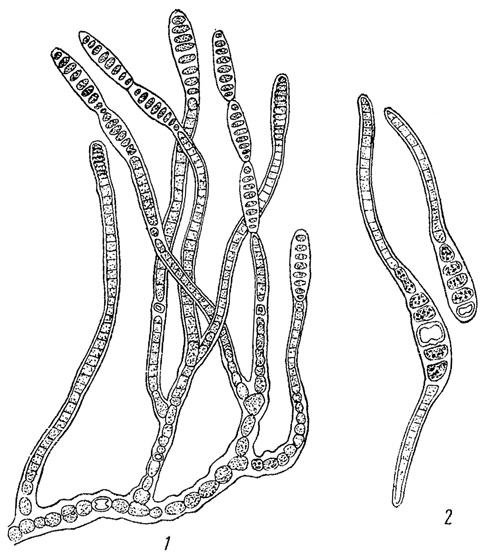
Рис. 44. Гормоспоры (гормоцисты) у синезеленой водоросли Leptopogori intricatus. 1 - образование гормоспор. 2 - две гормоспоры, прорастающие в нити
Отдельные фрагменты, на которые при вегетативном размножении распадается нитчатый таллом, в большинстве случаев состоят из нескольких клеток. Реже они одноклеточны, как это, например, имеет место у некоторых синезеленых водорослей, образующих гонидии и планококки (рис. 45) - одноклеточные фрагменты, сохраняющие слизистую оболочку (гонидии) или лишенные ее (планококки). Фрагментация нитей может вызываться механическими воздействиями, отмиранием отдельных клеток или быть независимой от того и другого и представлять собой в определенные периоды нормальное явление в жизни водоросли (часто образование гормогониев у синезеленых).
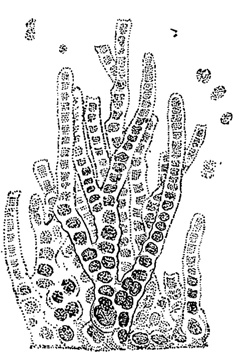
Рис. 45. Образование планококков у синезеленой водоросли Desmosiphon maculans
Наиболее специализированная форма вегетативного размножения наблюдается у некоторых видов харовых водорослей, образующих особые одноклеточные и многоклеточные клубеньки, богатые крахмалом (рис. 46). Одноклеточные клубеньки возникают из ризоидов, многоклеточные - представляют собой результат разрастания узлов на погруженных в ил частях "стебля". Клубеньки перезимовывают и весной дают начало новым растениям. Вегетативное размножение харовых происходит также с помощью специальных отростков - "побегов", вырастающих из нижних узлов и развивающихся впоследствии в новые растения.
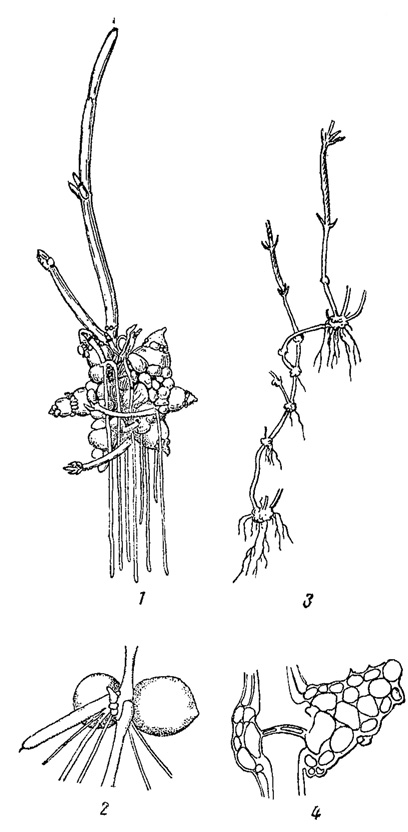
Рис. 46. Размножение клубеньками у харовых водорослей. 1 - прорастающий клубенек Nitellopsis obtusa. 2 - корневые клубеньки Chara aspera. 3 - нижняя часть побега Ch. baltiea с клубеньками. 4 - стеблевой клубенек Ch. baltica в продольном разрезе
При бесполом размножении клетки, посредством которых оно совершается, - споры - всегда покидают оболочку материнской клетки, что отличает эту форму размножения от простой фрагментации. Эти споры могут иметь разное строение, в соответствии с чем получают и различное наименование. Клетки, в которых они образуются, в большинстве случаев ничем не отличаются от других вегетативных клеток; реже они имеют иную форму и более крупные размеры и называются тогда спорангиями (бурые, красные водоросли, а из зеленых водорослей - Trentepohlia и некоторые другие).
Большинство водорослей, стоящих на разных ступенях морфологической дифференциации, размножается бесполым путем с помощью подвижных спор - зооспор (рис. 47, 48). Зооспоры имеют монадную структуру, в огромном большинстве случаев они голые и развиваются в клетках чаще в числе нескольких, так что процесс их образования связан с делением протопласта (одним или многими). Впрочем, количество их, в зависимости от вида, колеблется от одной (Oedogonium из зеленых водорослей) до нескольких сот (Cladophora из зеленых водорослей).
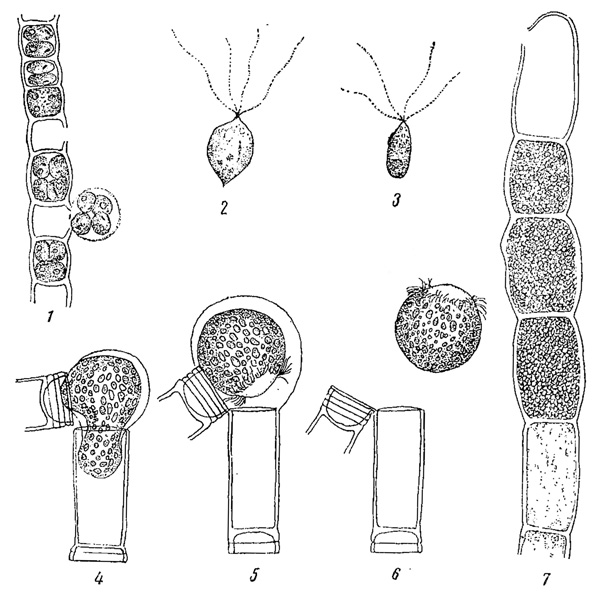
Рис. 47. Образование зооспор у зеленых водорослей. 1 - выход зооспор из клеток Ulothrix zonata. 2, 3 - две зооспоры U. zonata. 4-6 - последовательные стадии выхода зооспоры у Oedogonium concatenatum. 7 - образование зооспор в клетках Cladophora
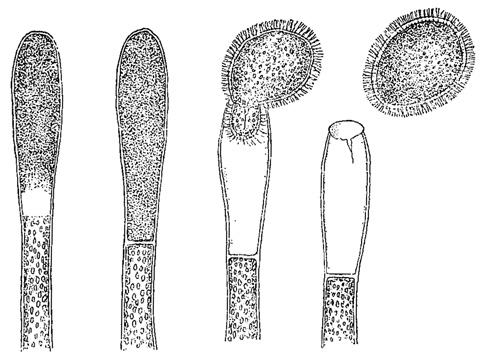
Рис. 48. Последовательные стадии образования зооспоры у зеленой водоросли Vauclieria repens
Зооспоры выходят наружу через отверстие, образующееся в оболочке материнской клетки (круглое или щелевидное), или благодаря ее общему ослизнению. Иногда в момент выхождения они окружены тонким слизистым пузырем. Каждая зооспора при благоприятных условиях может дать начало новому организму. При этом, если зооспора принадлежит форме, неподвижной в вегетативном состоянии, она, поплавав некоторое время, прикрепляется к какому-либо подводному предмету и теряет жгутики, после него из нее развивается водоросль, подобная той, из которой она произошла. Подвижность зооспор обеспечивает быстрое распространение производящих их водорослей на большие расстояния.
Наличие или отсутствие зооспор в цикле развития водоросли - важный систематический признак. То же относится и к их форме и строению, которые в разных группах далеко неодинаковы. Особенно большое значение имеет число жгутов, место их отхождения (от переднего конца или сбоку), а также - при наличии двух жгутов - их взаимная длина (одинаковая или неодинаковая). По числу жгутов зооспоры бывают одножгутиковыми, двужгутиковыми, четырехжгутиковыми и многожгутиковыми.
Своеобразное строение зооспор имеют представители порядка эдогониевых (из зеленых водорослей); здесь, несколько отступя от переднего (бесцветного) конца зооспоры, располагается целый венец жгутиков (рис. 47, 4-6).
Многожгутиковые зооспоры зеленой водоросли Vaucheria (характеризующейся сифональной структурой) правильнее рассматривать как агрегат многих двужгутиковых зооспор, не обособившихся друг от друга (рис. 48).
У некоторых водорослей (многие хлорококковые из зеленых водорослей, многие желтозеленые и др.) споры, посредством которых происходит бесполое размножение, в отличие от зооспор, лишены жгутиков и одеты оболочкой. Они развиваются в клетке так же, как зооспоры и их считают вторичным образованием - зооспорами, утратившими свою подвижность (рис. 49). Подобные неподвижные споры называют вообще апланоспорами, но, в зависимости от их строения, они получают и другие наименования. Иногда, находясь еще в материнской клетке, апланоспоры приобретают форму, вполне подобную ей, являясь как бы ее уменьшенной копией. В этих случаях говорят об автоспорах (рис. 50). У колониальных хлорококковых автоспоры уже в материнской клетке слагаются в колонии характерной для данного вида формы, так что после разрыва оболочки наружу выходит целая маленькая колония. Апланоспоры могут принимать на себя функцию покоящейся стадии, в связи с чем их оболочка утолщается; это - так называемые гипноспоры (например, Ulolhrix из зеленых водорослей, рис. 51). Гипноспоры отличаются от акинет тем, что они всегда развивают свою собственную оболочку, в образовании которой оболочка материнской клетки не участвует.
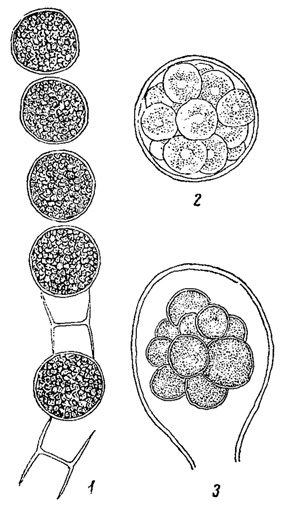
Рис. 49. Образование апланоспор у зеленых водорослей. 1 - Microspora Willeana. 2 - Trebouxia cladoniae. 3 - Prolosiphon botryoides
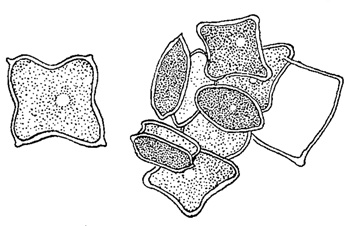
Рис. 50. Образование автоспор у зеленой водоросли Tetraedron minimum
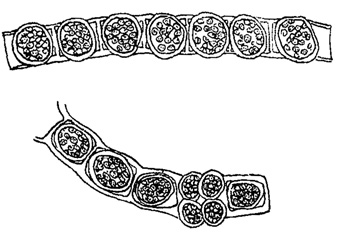
Рис. 51. Образование гипноспор у Ulothrix
Своеобразны неподвижные споры некоторых синезеленых, а также красных и бурых водорослей. У первых (в классе Chamaesiphoneae) встречается два рода таких спор - эндоспоры и экзоспоры. Эндоспоры (рис. 52) развиваются в клетках в большом числе (до 128) и, в сущности, отличаются от апланоспор только отсутствием оболочки, хотя у некоторых форм (например, Chlorococcopsis) она имеется. Экзоспоры, в отличие от эндоспор, по мере своего образования, отчленяются и выходят наружу (рис. 53). У встречающейся в пресных водах красной водоросли Batrachospermum неподвижные споры развиваются в спорангиях в единственном числе, почему их и называют моноспорами (рис. 54). У других представителей этого типа чаще встречается образование четырех спор в одном спорангии (тетраспоры). Тетраспоры характерны и для некоторых бурых водорослей (порядок диктиотовых).
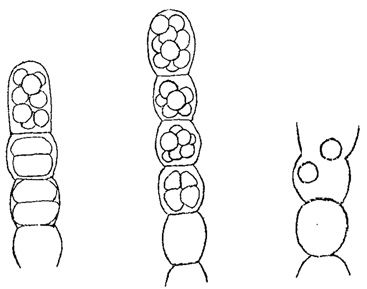
Рис. 52. Образование эндоспор у синезеленой водоросли Endonema moniliforme
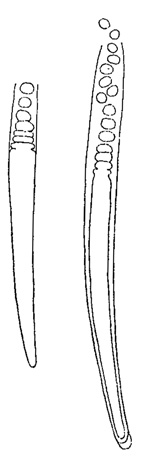
Рис. 53. Образование экзоспор у синезеленой водоросли Chamaesiphon curvatus
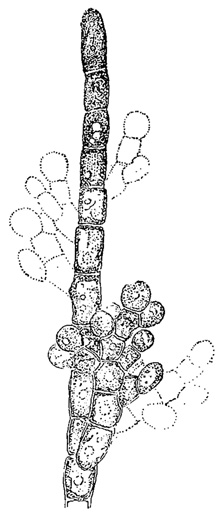
Рис. 54. Образование спор у Batrachospermum anatinum
Половое размножение осуществляется посредством половых клеток, имеющих у разных водорослей различное строение, но носящих общее наименование гамет. Половой процесс (оплодотворение), представляющий собой характерную особенность полового размножения, заключается в слиянии (копуляции) двух гамет в одну клетку - зиготу, причем сливается не только цитоплазма, но и ядра. Зиготы в зрелом состоянии обычно имеют толстую, нередко покрытую скульптурой оболочку, содержат большое количество запасных питательных веществ и, после периода покоя, прорастают, давая начало новым организмам*. Таким образом, зигота является покоящейся стадией.
* (Период покоя наблюдается не у всех зигот; он особенно характерен для зигот пресноводных водорослей.)
В редких случаях гаметы дают начало новому организму без всякого оплодотворения, т. е. без образования зиготы. Такое упрощенное половое размножение называется партеногенезом. Партеногенез встречается у некоторых зеленых, а также бурых водорослей как сравнительно редкое явление. Однако у некоторых видов он представляет собой обычную форму размножения, как это имеет место у Chara canescens (из харовых водорослей) и Spirogyra mirabilis (из зеленых водорослей). У обоих этих водорослей споры, подобные зиготам, развиваются из одной гаметы; их называют а зиготами или партеноспорами. С другой стороны, ниже мы увидим, что процесс оплодотворения иногда может быть и не связан с размножением.
Половое размножение водорослей отличается большим разнообразием, однако, основные его формы могут быть сведены к сравнительно немногим типичным случаям.
Строение гамет в разных группах водорослей, так же как и строение зооспор, различно, но чаще всего мужские гаметы (а нередко и женские) подвижны и имеют монадную структуру. Такие гаметы крайне напоминают зооспоры, часто отличаясь от них только меньшими размерами. Они образуются в результате деления протопласта в клетках, нередко вполне подобных другим клеткам, не участвующим в размножении. В более сложных случаях клетки, дающие начало гаметам, более или менее отличны; их называют тогда гамотангиями.
Большой интерес представляет дифференциация гамет на мужские и женские. У некоторых водорослей обе сливающиеся между собой гаметы подвижны и морфологически неотличимы друг от друга. Подобные гаметы носят название изогамет, а самый половой процесс обозначают как изогамию (рис. 55). Далее замечено, что при изогамии нередко копулируют гаметы, развивающиеся из одной особи или колонии; в этом случае говорят, что водоросль гомоталлична. С другой стороны, часто копуляция возможна только между гаметами, произошедшими от разных особей или колоний; это значит, что водоросль гетероталлична. При гетероталлизме нельзя еще сказать, какая гамета является мужской, а какая женской, так как морфологически они вполне подобны друг другу, однако различия физиологического порядка здесь, несомненно, налицо, и поэтому принято различать + гаметы и - гаметы. Изогамия широко распространена среди водорослей. Мы находим ее у многих зеленых водорослей монадной, пальмеллоидной, коккоидной, нитчатой и сифональной структур, а также у бурых водорослей. В одних случаях имеет место гомоталлизм (например, у некоторых Chlctmydomonas), в других - гетероталлизм (например, Gonium, Pandorina из порядка вольвоксовых, некоторые Chlamydomonas).
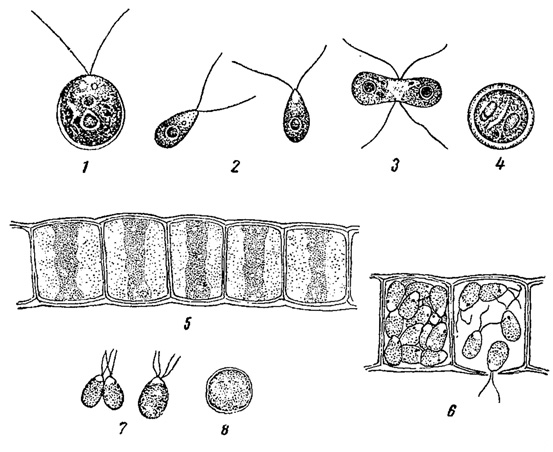
Рис. 55. Изогамия у зеленых водорослей. 1-4 - Chlamydomonas Reinhardii: 1 - отдельная особь, 2 - изогаметы, 3 - копуляция изогамет, 4 - зигота. 5-8 - Ulothrix zonata: 5 - часть нити, 6 - образование изогамет, 7 - копуляция изогамет, 8 - зигота
Изогамия связана переходами с гетерогамией. Этим термином обозначают такой половой процесс, при котором копулирующие гаметы (гетерогаметы), хотя и являются обе подвижными, но различаются между собой размерами, а иногда и формой. Более крупная гамета (макрогамета) является женской, более мелкая (микрогамета) - мужской (рис. 56, 57). Гетерогамия также встречается у многих водорослей, хотя все же реже, чем изогамия. Интересно, что у форм с гетерогамным половым процессом наблюдается разная степень дифференцировки мужских и женских гамет. Иногда (например, у Phyllobium, Draparnaldia и других зеленых водорослей) различия между макро- и микрогаметами крайне невелики, иногда же (например, у Eudorina, Aphanochaete и др.) - весьма значительны, так что мужские гаметы напоминают сперматозоиды, а женские - яйцеклетку.
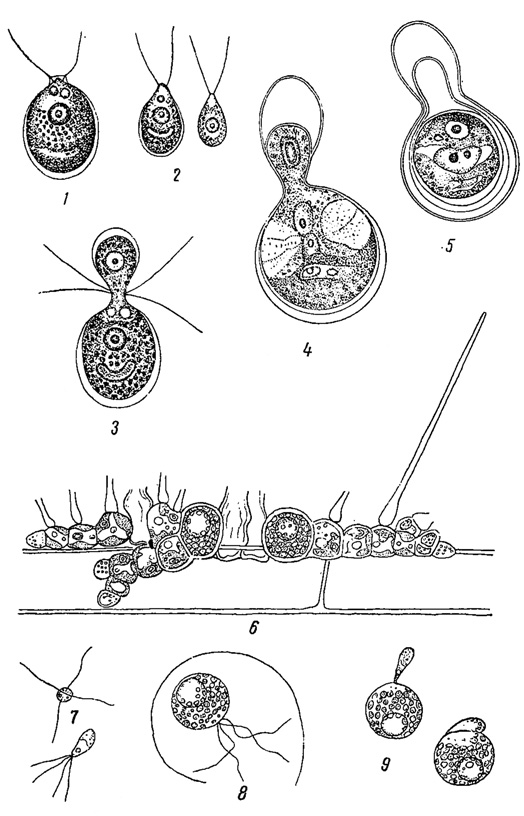
Рис. 56. Гетерогамия у зеленых водорослей. 1-5 - Chlamydomonas Braunii: 1 - отдельная особь, 2 - гетерогаметы, 3-5 - последовательные стадии копуляции гетерогамет. 6-9 - Aphanochaete repens: 6 - слоевище Aph. repens на нити водоросли, 7 - микрогаметы, 8 - макрогамета, 9 - их копуляция
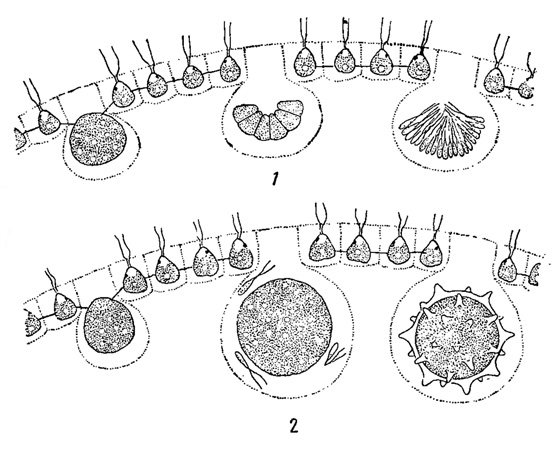
Рис. 57. Гетерогамия у зеленых водорослей. 1, 2 - Phyllobium dimorphum: 1 - выход гетерогамет, 2 - их копуляция. 3 - Eudorina elegans (микрогаметы, окружающие колонию с макрогаметами)
Наибольшая дифференциация гамет наблюдается при оогамии, встречающейся у различных водорослей, имеющих разнообразную морфологическую структуру и относящихся к разным типам. Оогамия характеризуется тем, что женские гаметы имеют вид яйцеклеток, а мужские - сперматозоидов (антерозоидов). Яйцеклетки - крупные, неподвижные, развиваются в малом количестве; сперматозоиды - мелкие, весьма подвижные, развиваются в очень большом числе (рис. 58, 59). Гаметангии, образующие яйцеклетку, носят общее название оогониев; гаметангии, дающие начало сперматозоидам, называются антеридиями. Сперматозоиды по созревании всегда выходят наружу из антеридия. Яйцеклетки же у одних форм (например, некоторые виды Chlorogonium из зеленых водорослей) тоже выходят наружу, так что оплодотворение происходит вне тела растения, а у других (например, Volvox и мн. др.) - остаются внутри оогония, где и происходит копуляция гамет (рис. 59). Зигота, получающаяся в результате оплодотворения яйцеклетки сперматозоидом, носит название ооспоры. Оогонии и антеридии развиваются то на одной особи или колонии (обоеполость или однодомность), то на разных (раздельнополость или двудомность). У многих видов Oedogonium (из зеленых водорослей) клетки нитчатого слоевища дают начало не антеридиям, а особым спорангиям - андроспорангиям. В них развиваются подвижные клетки, напоминающие зооспоры, но меньших размеров (андроспоры). Последние подплывают к оогонию и прорастают здесь в короткую (двуклеточную) нить, на которой образуется антеридий. Этот антеридий дает уже начало сперматозоидам (рис. 60).
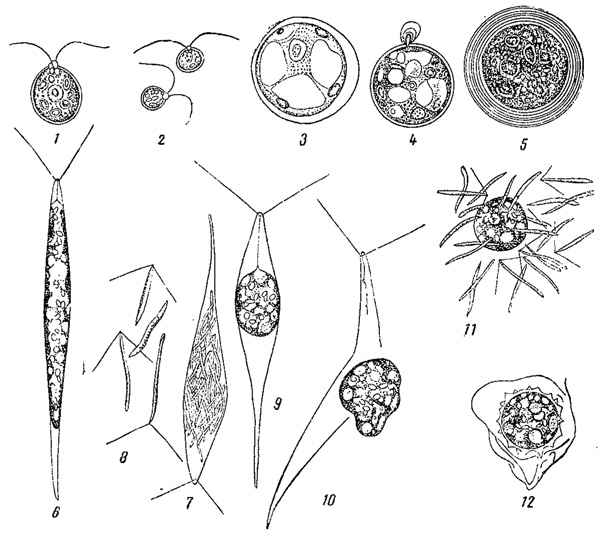
Рис. 58. Оогамия у зеленых водорослей. 1-5 - Chlamydomonas coccifera: 1 - отдельная особь, 2 - сперматозоиды, 3 - яйцеклетка, 4 - оплодотворение, 5 - зигота. 6-12 - Chlorogomum oogamum: 6 - отдельная особь, 7 - образование сперматозоидов, 8 - вышедшие нарушу сперматозоиды, 9, 10 - образование яйцеклетки, 11 - оплодотворение, 12 - зигота
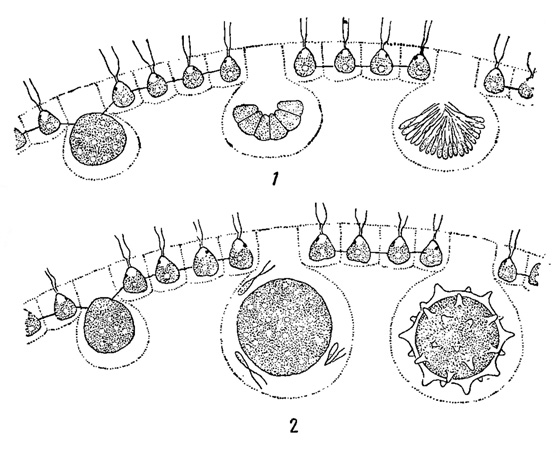
Рис. 59. Оогамия у Volvox. 1 - часть мужской колонии в разрезе (видны последовательные стадии образования сперматозоидов). 2 - часть женской колонии в разрезе (видна молодая яйцеклетка, оплодотворение, зигота) (схематизировано)
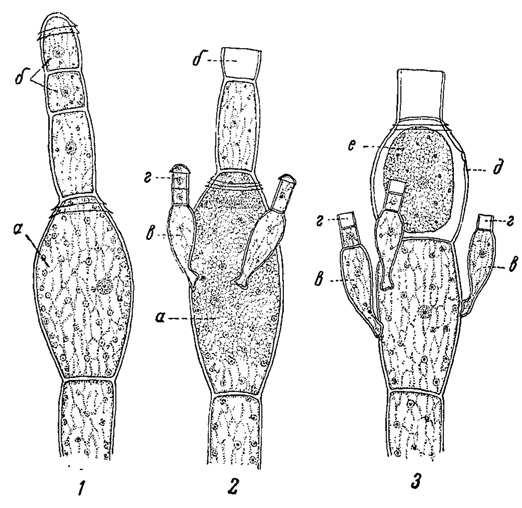
Рис. 60. Последовательные стадии размножения Oedogonium concatenatum. а - материнская клетка оогония; б - андроспорангий; в - нить, развившаяся из андроспоры; г - антеридий; д - оогоний; е - яйцеклетка
Та или иная форма полового процесса (изогамия, гетерогамия, оогамия) является характерной особенностью различных видов, но в пределах более крупных систематических единиц нередко встречаются разные формы. Так, среди зеленых водорослей в порядке вольвоксовых известны изо-, гетеро- и оогамия, в порядке хлорококковых - изо- и, изредка, гетерогамия, в порядке хетофоровых - изо-, гетеро- и оогамия и т. д. У Chlamydomonas (порядок хламидомонадовых) все три формы полового процесса наблюдаются в пределах рода. С другой стороны, существуют группы, все представители которых характеризуются однотипным (в смысле дифференциации гамет) половым процессом. Так, все эдогониевые, сфероплейные и вошериевые оогамны. Резко выраженная оогамия характерна для типа харовых водорослей. У последних половые органы (мужской - антеридий и женский - оогоний или споропочка) имеют сложное строение и отличаются от других водорослей своей многоклеточностью (рис. 61). Оогамный половой процесс наблюдается также у многих бурых и всех красных водорослей. У последних он протекает крайне своеобразно. Сперматозоиды здесь всегда лишены жгутиков и не способны к активному движению; их называют спермациями. Мужской половой орган - антеридий - дает начало одному спермацию. Женский половой орган, называемый карпогоном, обычно имеет бутылкообразную форму (рис. 62). Из оплодотворенной яйцеклетки развиваются особые неподвижные клетки - карпоспоры, дающие впоследствии начало новым растениям. У разных красных водорослей образование карпоспор происходит различно, и процесс этот иногда весьма сложен.
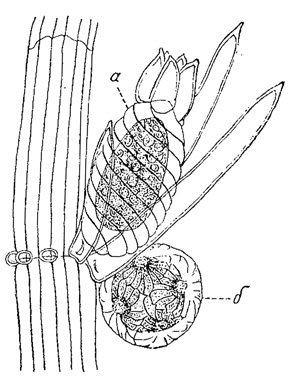
Рис. 61. Половые органы Chara fragilis. а - споропочка; б - антеридий
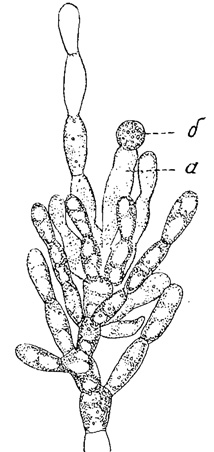
Рис. 62. Часть слоевища красной водоросли Batrachospermum ectocarpum. а - карпогон; б - спермаций
Отсутствие гамет, обладающих органоидами движения - жгутиками, - характерно также для зеленых водорослей подтипа Conjugatae и, повидимому, для диатомовых*. Здесь половой процесс заключается в слиянии протопластов двух обычных вегетативных клеток, которые и выполняют функцию гамет; специальные дифференцированные гаметы отсутствуют. Подобный половой процесс часто называют конъюгацией. Так, в порядке зигнемовых (из Conjugatae) между клетками двух разных нитей (или одной нити) образуется сквозной канал (конъюгационный или копуляционный канал), через который протопласт одной клетки переливается в другую и сливается с ее протопластом, давая зиготу. Иногда зигота развивается в самом конъюгационном канале (рис. 63). В одной нити обычно образуется много зигот, из которых каждая при благоприятных условиях может дать начало новому нитчатому слоевищу. Таким образом, половой процесс здесь, как и в выше рассмотренных случаях, сочетается с размножением. У одноклеточных Conjugatae порядка десмидиевых при конъюгации двух особей развивается одна зигота, дающая затем обычно два проростка, так что умножение особей не имеет места (рис. 64). У диатомовых водорослей (в классе Pennatae) конъюгация по внешности напоминает аналогичный процесс у Conjugatae, хотя цитологически резко от него отличается. Зиготы здесь превращаются в ауксоспоры - особые растущие стадии, дающие начало новому организму. Наконец, у некоторых других диатомовых (класс Centricae) имеет место редкая форма полового процесса, называемая автогамией (самооплодотворением) и заключающаяся в слиянии двух ядер, образующихся внутри одной клетки.
* (Некоторые (преимущественно морские) диатомовые, относящиеся к классу Centricae, способны образовывать подвижные, имеющие жгутики, клетки, которые иногда рассматриваются как гаметы. Однако копуляция их не наблюдалась, и весьма вероятно, что это зооспоры.)
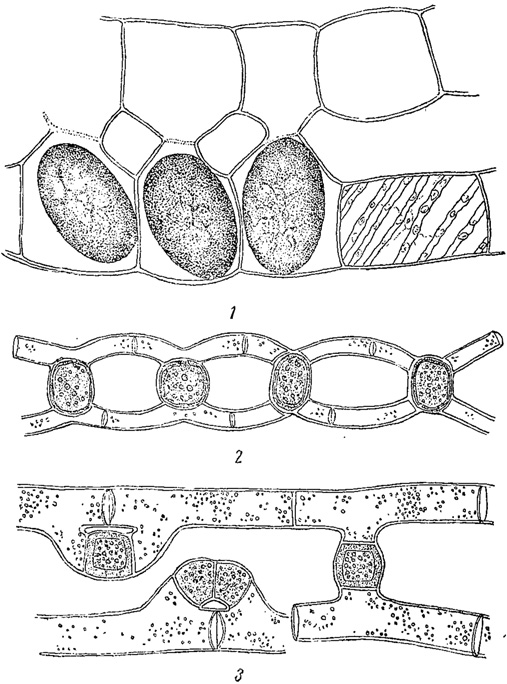
Рис. 63. Конъюгация у зигнемовых водорослей. 1 - Spirogyra Reinhardii. 2 - Mougeotia calcarea. 3 - M. mirabilis
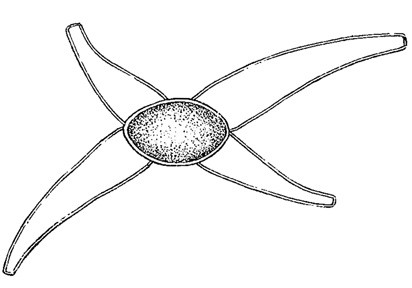
Рис. 64. Конъюгация у десмидиевой водоросли Closterium Elenkinii
Разные формы размножения часто наблюдаются у одного и того же вида, причем смена форм размножения иногда сочетается со сменой форм развития*. При смене форм развития происходит смена особей, размножающихся то только бесполым, то только половым путем. В этих случаях жизненный цикл водоросли включает в себя, следовательно, как половую форму развития, так и бесполую, и ни одна из них, взятая раздельно, не может охарактеризовать его в полной мере. При таких усложненных циклах развития те особи, которые могут образовывать только половые клетки (гаметы), получили название гаметофитов, другие же, образующие только клетки бесполого размножения (споры), называются спорофитами.
* (Сменой форм развития мы называем здесь то, что в специальной и учебной литературе до сих пор обычно называли "чередованием поколений".)
Смена форм развития у водорослей отличается большим разнообразием, которое сводится к следующим двум типам: иногда обе формы (половая и бесполая) морфологически вполне подобны друг другу (изоморфная смена), иногда же различны (гетероморфная смена). В пределах второго типа соотносительная степень развитости гаметофита и спорофита может быть весьма неодинаковой, причем, как правило, преобладает спорофит, вплоть до того, что в некоторых случаях гаметофит имеет лишь микроскопические размеры, тогда как спорофит достигает весьма мощного развития (например, у морской бурой водоросли Laminaria).
Смена форм развития, проявляясь обычно с большой правильностью, может все же резко нарушаться в зависимости от тех или иных условий существования.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'