19.2.7. Порядок 7. Эдогониевые - Oedogoniales (В. П. Юнгер)
Порядок Oedogoniales объединяет зеленые нитчатые водоросли, для которых характерно деление вегетативных клеток с образованием колпачков, особое строение монад - с венчиком жгутиков на переднем конце и прогрессивная редукция межских растений до одной клетки - антеридия. В мировой флоре известно 655 видов этих водорослей, во флоре Украины - немногим более 180.
Индивиды эдогониевых водорослей имеют нитчатый таллом. Нити простые (Oedogonium Link) или разветвленные (Bulbochaete Ag., Oedocladium Stachl), преимущественно прикрепленные, заканчиваются тупой или заостренной клеткой (Oedogonium, Oedocladium) или волоском (Bulbochaete, Oedogonium), могут достигать в длину до 50 см, но обычно не превышают нескольких миллиметров (рис. 19.30). Прикрепляются к субстрату базальной клеткой с ризоидальными выростами, дисковидным дермоидом, полушаровидной или сферической стопой (Bulbochaete, Oedogonium), либо посредством разветвленных, слабоокрашенных ризоидных нитей (Oedocladium). Рост апикальный (Oedocladium), базальный и интеркалярный (Bulbochaete) или диффузный (Oedogonium).
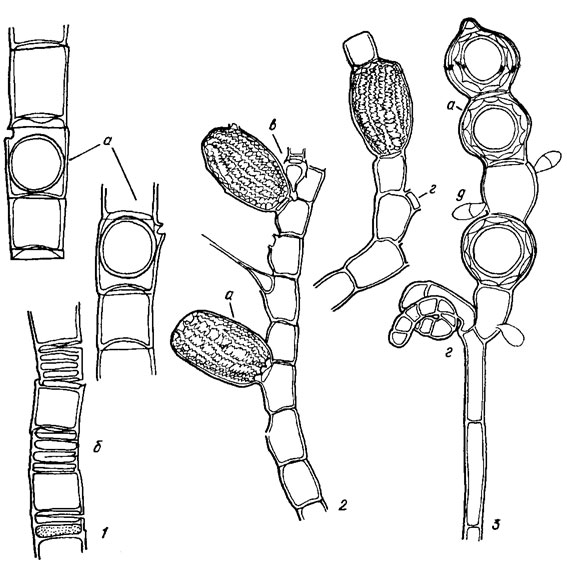
Рис. 19.30. Oedogoniales: 1 - Oedogonium capillare (L.) Kütz. (a - оогонии, б - антеридии); 2 - Bulbochaete rectangularis Wittr. (a - оогонии, в - наннандрий с антеридием, г - андроспорангиальные клетки); 3 - Oedocladium caroiinianum Beaney et Hoffmann (а - оогонии, г - андроспорангиальные клетки, д - наннандрий - антеридий)
Клетки преимущественно цилиндрические (Oedogonium, Oedocladium) или обратноконические (Bulbochaete). Оболочки клеток трехслойные; внутренний слой - целлюлозный, средний - пектиновый, наружный (кутикулярный) - предположительно хитиновый. Однако исследования Б. К. Паркера [574] наличия хитина не подтвердили. Целлюлозный слой согласно данным электронной микроскопии состоит из нескольких слоев микротрубочек, ориентированных вдоль оси нитей [574]. Наружная поверхность клеточной стенки гладкая (Oedocladium) или иногда со спиральной пунктацией (Oedogoniurn, Bulbochaete), природа которой не ясна. Нередко наружные оболочки клеток инкрустированы солями кальция или оксидами железа, что затрудняет микроскопирование эдогониевых водорослей.
Для представителей порядка Oedogoniales характерно наличие колпачков, образующихся при вегетативном делении клеток - т. н. линейном почковании [305]. В начале деления в верхней части клетки, ближе к ее вершине, на внутренней стороне оболочки образуется складка, которая утолщается и постепенно превращается в кольцо (рис. 19.31). В это время ядро переходит из периферической части клетки в центральную и делится митотически. По окончании кариокинеза и расхождения ядер стенка материнской клетки в районе кольца разрывается, кольцо раскрывается и разрастается, обеспечивая интенсивный рост дочерней клетки. Остатки оболочки материнской клетки остаются на дочерней в виде колпачка. Число колпачков соответствует количеству делений данной клетки. Деление вегетативных клеток эдогониевых водорослей впервые описал Н. Прингсхейм [597]. Он же выяснил роль кольца в нем. Дальнейшее изучение деления клеток эдогониевых в основном сводилось к изучению строения кольца и путей его возникновения. Большинство исследователей [342, 485 и др.] считали, что кольцо представляет собой впячивание внутреннего слоя оболочки материнской клетки. В настоящее время электронно-микроскопическими исследованиями показано, что кольцо является протоплазматическим новообразованием, возникающим вследствие деятельности аппарата Гольджи, и на ранних стадиях развития состоит из системы диктиосом [480].
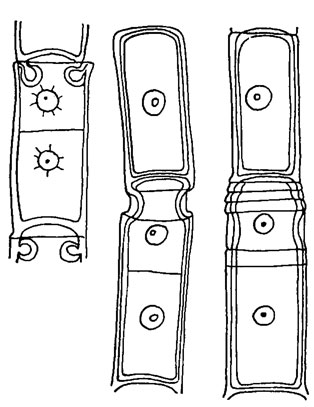
Рис. 19.31. Деление клеток у Oedogonium borisianum Wittr
Ядро обычно крупное, сферическое или несколько продолговатое, в интерфазе четко очерчено и ограничено ядерной оболочкой, содержит нуклеоплазму, хроматин, одно или несколько ядрышек. Митоз у эдогониевых водорослей изучен слабо, однако немногочисленные наблюдения в роде Oedogonium свидетельствуют о том, что он сходен с таковым у высших растений. В хромосомах Oedogoniales обнаружены центромеры, занимающие субтерминальное или медиальное положение. Количество хромосом колеблется от 13 до 38, известны полиплоиды. Хлоропласт один, пристенный, крупный, сетчато продырявленный, иногда до лентовидного, рассеченного на узкие полосы (рис. 19.32). Ультраструктурное строение хлоропласта эдогониевых такое же, как и у большинства зеленых водорослей. Пиреноиды размещены в строме хлоропласта. Их количество непостоянно - от 1 до 20 и более. При наблюдении в электронный микроскоп виден зернистый матрикс пиреноида, окруженный двойной цитоплазматической мембраной; матрикс пиреноида нередко пронизан цитоплазматическими канальцами [488]. Фотосинтезирующие пигменты у эдогониевых такие же, как и у остальных зеленых водорослей: хлорофиллы а и b, каротины, лютеин и ксантофиллы (виолоксантин и зеаксантин). Основной продукт ассимиляции - крахмал, накапливающийся вокруг пиреноидов в особых крахмальных влагалищах.
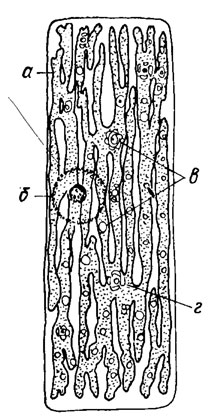
Рис. 19.32. Схема строения вегетативной клетки Oedogonium: а - хлоропласт; б - ядро; в - пиреноиды; г - зернышко крахмала
У эдогониевых водорослей наблюдается половое и бесполое размножение. Вегетативное размножение происходит с помощью специальных клеток - акинет, или фрагментацией таллома. Фрагментация нитей в роде Oedogonium встречается часто, в родах Bulbochaete и Oedocladium - значительно реже. Сведения о прорастании фрагментов в полноценные талломы отсутствуют. Акинеты - тонкостенные красноватые или оранжевые клетки, образующиеся из любой клетки таллома, расположенные по одной (Oedogonium) или по несколько (3-4) подряд (Oedocladium). У видов рода Oedocladium кроме того в подземной части таллома образуются геммы, отличающиеся от акинет только местом возникновения, а также короткие боковые веточки, состоящие из нескольких оранжевых клеток.
Бесполое размножение осуществляется зооспорами, развивающимися по одной в любой клетке таллома, за исключением базальной. Зооспоры овалоидные, на переднем конце несут венчик жгутиков (100-120 жгутиков у Oedogonium и 20-30 у Bulbochaete). Жгутики расположены парами, каждый из них окружен цитоплазматической мембраной, являющейся продолжением мембраны протопласта, поверхность их гладкая, по строению они подобны жгутикам других подвижных клеток растений (структура "9 + 2"). Основания каждой пары жгутиков соединяются с базальными телами, связанными между собой фиброзным кольцом, состоящим из микрофибриллярных волокон (рис. 19.33). Между каждой парой трубочек от фиброзного кольца отходят по два радиально расположенных жгутиковых корешка, один из которых микрофибриллярный, направлен наружу от кольца, второй, внутренний, состоящий из 3 микротрубочек, направлен к ядру клетки, но не соединяется с ним. Столь сложно устроенный жгутиковый аппарат, по-видимому, уникален для зеленых водорослей. Зооспоры эдогониевых водорослей, кроме того, обязательно содержат ядро, хлоропласт, цитоплазму, сократительную вакуоль, стигму и другие клеточные компоненты. После выхода из зооспорангия зооспоры свободно плавают в течение некоторого времени (до 2 ч), затем передним концом прикрепляются к субстрату и прорастают в новое растение, причем у видов Oedogonium и Bulbochaete тело зооспоры непосредственно превращается в базальную клетку, а ее бесцветный конец - в ризоидальный вырост.

Рис. 19.33. Строение жгутикового аппарата у Oedogonium cardiacum (Hass.) Wittr
Половое размножение у эдогониевых водорослей всегда оогамное, причем образование женских и мужских половых клеток может происходить на одном и том же растении (однодомные виды) или на различных растениях (двудомные виды). По строению мужских растений двудомные виды можно разделить на две группы: 1) с мужскими талломами, аналогичными построению женским; 2) с карликовыми мужскими одноклеточными и многоклеточными растениями - наннандриями. Наннандрии, как одноклеточные, так и многоклеточные развиваются из андрозооспор. Андрозооспоры морфологически подобны остальным монадам эдогониевых. У одних видов они образуются на тех же талломах, что и оогонии (гинандроспоровые виды), у других - на особых андроспорангиальных растениях (идиоондроспоровые виды). У видов с одноклеточными наннандриями андрозооспоры, прикрепившись, или непосредственно превращаются в антеридии, или предварительно делятся без образования кольца. У видов с многоклеточными наннандриями андрозооспора, прикрепившись, превращается в базальную клетку, от которой путем нормального для Oedogoniales деления с образованием кольца отшнуровывается несколько вегетативных, а затем и антеридиальных клеток. Антеридиальные клетки как у однодомных, так и у двудомных видов по форме отличны от вегетативных (см. рис. 19.30). Они значительно короче вегетативных клеток, встречаются одиночно или группами. В каждом из антеридиев образуется по две, реже по одной мужской гамете - сперматозоиду. Сперматозоиды эдогониевых водорослей очень похожи на зооспоры, но имеют меньшие размеры и меньшее количество жгутиков. Оогонии обычно крупнее вегетативных клеток, в каждом из них развивается по одной ооспоре. Оогонии могут располагаться одиночно или группами, их оболочка иногда имеет крыловидные выросты (Oedogonium). Оогонии открываются овальным отверстием, щелью или крышечкой. Способ раскрытия оогония является важным таксономическим признаком для видов рода Oedogonium.
Оплодотворение у эдогониевых водорослей - сложный многоступенчатый процесс. Например, у Oedogonium borisianum Wittr., андрозооспоры хемотаксически привлекаются к инициальной клетке оогония, развитие которого начинается только после завершения развития наннандрия; уже сформировавшийся оогоний выделяет массивные слизистые оболочки, перекрывающие и наннандриевые растения, т. е. оплодотворяющий сперматозоид движется в желатиновом пузыре. Оплодотворенная яйцеклетка превращается в зиготу (оослору), которая обычно окружена толстой трехслойной оболочкой. Наружная и средняя оболочки зиготы могут быть гладкими или скульптурированными (шиповатыми, бородавчатыми, выемчатыми, ребристыми и т. д.). Строение оболочки - важный таксономический признак на видовом уровне в каждом из родов Oedogoniales. После периода покоя ооспора делится мейотически и прорастает, образуя четыре мейозоспоры, которые, проплавав некоторое время, прикрепляются к субстрату и прорастают в новые нити. Следовательно, цикл развития эдогониевых водорослей гаплофазный, диплоидна лишь зигота, которая мейотически делится только при образовании зооспор, т. е. у эдогониевых водорослей наблюдается спорическая редукция.
Серьезной трудностью в изучении эдогониевых водорослей является то, что их систематика на видовом уровне основывается на признаках, связанных со структурами, возникающими при половом воспроизведении - строении мужских растений, антеридиев, оогониев и ооспор. В то же время период полового воспроизведения у некоторых видов этих водорослей может быть очень недолгим*.
* (Для возможно более полного выявления видового состава эдогониевых водорослей необходимы продолжительные стационарные исследования. Кратковременные экспедиционные выезды рациональнее всего планировать на период, когда можно встретить наибольшее количество видов со сформированными оогониями и антеридиями (на Украине - июнь, июль).)
Сомнений в принадлежности эдогониевых к зеленым водорослям не возникает. В пользу этого безусловно свидетельствует набор фотосинтезирующих пигментов, характер продуктов ассимиляции и тонкое строение хлоропластов. Чаще всего эдогониевые водоросли рассматривают как самостоятельный порядок класса Chlorophyceae. Однако среди альгологов, изучающих эдогониевые водоросли, все большее распространение приобретает мнение о необходимости выделения их в самостоятельный класс Oedogoniophyceae отдела Chlorophyta, принимая во внимание своеобразие вегетативного деления и строения жгутикового аппарата монад [446, 679].
Эдогониевые водоросли в целом широко распространены в природе. Виды родов Oedogonium и Bulbochaete - типично водные организмы. Они встречаются во всех частях света за исключением Антарктиды. Представители рода Oedocladium, исключая один вид (Oedocladium hazenii Lewis ex Tiffany - водный организм), наземные водоросли. Они обнаружены в Юго-Восточной Азии, южных штатах США и в Австралии, единственная находка известна также в Европе. Виды родов Oedogonium и Bulbochaete распространены в разнообразнейших континентальных водоемах, от олиготрофных болот до лиманов, от гляциальных озер до термальных источников. Обычно они обрастают различные погруженные в воду предметы, выступая как эпилиты, эпиксилы и эпифиты, причем среди них явно преобладают эпифиты.
В некоторых случаях виды рода Oedogonium могут развиваться массово, и их нити, оторвавшись от субстрата, в этом случае обычно образуют мощные ватообразные скопления, засоряющие водоемы. Массовое развитие этих водорослей чаще происходит в пойменных водоемах, прудах и в эвтрофных озерах. Массовое развитие видов рода Bulbochaete наблюдается значительно реже. На Украине наибольшее видовое богатство эдогониевых водорослей (видов родов Oedogonium и Bulbochaete) выявлено в эвтрофных болотах и мелких, периодически пересыхающих водоемах, наименьшее - в реках и олиготрофных болотах.
Практическое значение эдогониевых водорослей обусловлено той ролью, которую они играют в континентальных водоемах. Эти водоросли представляют и значительный теоретический интерес в связи с их значительной геологической древностью (наиболее древние находки относятся к среднедевонским отложениям) и своеобразным положением среди зеленых водорослей.
Порядок Oedogoniales включает 1 семейство Oedogoniaceae De Bary, содержащее 3 рода: Oedogonium Link (530 видов), Bulbochaete Ag. (110 видов), Oedocladium Stachl (15 видов), отличающихся морфологией таллома (см. рис. 19.30). Наиболее распространенными в водоемах Украинской ССР являются Oedogonium capiHare (L.) Kütz., Oe. capilliforme Kütz., Oe. pringsheimii Cramer, Bulbochaete rectangularis Wittr. (см. рис. 19.30).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'