19.2.3. Порядок 3. Хлорококковые - Chlorococcales (Protococcales) (П. М. Царенко)
Хлорококковые водоросли довольно многочисленный и разнообразный порядок зеленых водорослей, насчитывающий свыше 1200 видов [510], из них около 450 видов известны во флоре УССР.
Преобладающее большинство видов являются хлорофилл содержащими организмами и имеют зеленую окраску. В основном это микроскопические формы, размеры которых колеблются от 0,5 до 100 мкм, обычно 5-20 мкм, и только виды рода Hydrodictyon Roth и отдельные слизеобразующие представители, способные формировать макроскопические колонии-разрастания, достигают 1,5 см.
К хлорококковым водорослям относятся одноклеточные, колониальные или ценобиальные, неподвижные в вегетативной фазе жизненного цикла, индивиды коккоидной структуры (как исключение, отдельные из них обладают сократительными вакуолями или стигмой), снабженные прочной целлюлозной или целлюлозо-спорополлениновой оболочкой, объединяющие моно- и ценоцитные формы (рис. 19.16). Одноклеточные формы образуют иногда скопления в виде однослойных пластинок или бесформенных конгломератов.
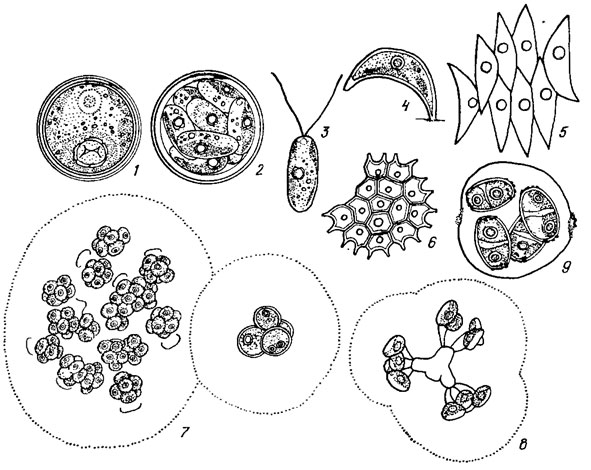
Рис. 19.16. Одноклеточные (1-4), ценобиальные (5, 6) и колониальные (7-9) Chlorococcales: 1-3 - Chlorococcum infusionum (Schrank) Menegh.; 4 - Characlum ornithoce phalum var. pringsheimii (A. Br.) Kom.; 5 - Scenedesmus acutus Meyen; 6 - Pediastrum boryanum (Turp.) Menegh.; 7 - Coenochloris pyrenoidosa Korsch.; 8 - Dictyosphaerium tetrachotomum Printz; 9 - Granulocystopsls pseudocoronata (Korsch.) Hind
Колонии чаще всего с более или менее развитым слизистым покровом, в котором отдельные клетки расположены свободно (не связаны друг с другом) или соединены с помощью слизистых нитей, стеблей или остатков оболочки материнских клеток, в то время как в ценобии клетки соединены непосредственно своими боковыми поверхностями или с помощью выростов клеточной оболочки. Широко распространенной формой колонии является сферическая или овальная, только иногда встречается древовидная, гроздевидная или сетевидная (подсемейство Ecballocystoideae). Иногда наблюдается заметный рост клеточной оболочки, которая становится ограничивающим барьером колонии из нескольких генераций и определяет ее форму (Oocystасеае).
Форма клеток хлорококковых также довольно разнообразна - от шаровидной до цилиндрически-веретеновидной, спиралевидной или трапециевидной. Однако преобладающей формой клетки является шаровидная или овальная до яйцевидной, нередко с разнообразными отростками. Прикрепленные формы развивают специальные органы крепления - ножку, стопу или только слизистую подушечку.
Клеточная оболочка большинства видов сплошная, цельная, гладкая, в то время как у других - состоит из двух или нескольких сегментов (Treubariaceae). Обычно она имеет несколько слоев: внутренний широкий слой из целлюлозы и один или несколько внешних. Тонкие внешние слои состоят из пектина, спорополленина, иногда полимерных полисахаридов. В некоторых случаях оболочка многослойная, состоит только из целлюлозы с характерными утолщениями в определенных местах и способна сильно ослизняться или заметно расширяться.
Внешний слой оболочки имеет различного рода ультраструктуру - гладкую, скробикулированную, сетчатую, складчатую и т. п., которая иногда различима при наблюдении в световой микроскоп как пунктированная, морщинистая или ребристая. Составными элементами этого слоя являются разнообразные шиповидные образования - иглы, шипы, щетинки, зубчики (рис. 19.17), которые различаются между собой как формой, относительными размерами, временем возникновения, так и химическим составом [461]. Щетинки, состоящие из пучков целлюлозных микрофибрилл, характерны для Golenkinia Chod., Acanthosphaera Lemm., Echinosphaeridium Lemm., а шипы, содержащие нити протеина, установлены для Scenedesmis Meyen, Polyedriopsis Schmidle, Pediastrum Meyen, Micractinium Fres., Lagerheimia Chod., Siderocystopsis Swale [461]. Для видов родов Scenedesmus и Tetrastrum Chod. известны также шипы (зубчики), состоящие из спорополленина и являющиеся частью клеточной оболочки [461]. У этих двух родов внешний спорополлениновый слой имеет своеобразную структуру и особые поверхностные образования - розетки, сетки, бородавки, сосочки, ребра, шипики [460]. Довольно разнообразна ультраструктура щетинок у планктонных видов родов Scenedesmus, Pediastrum и Coelastrum Näg., которые выполняют функцию плавательного аппарата. Развитие щетинок зависит от экологических условий. Среди других поверхностных структур оболочки ясно выделяются отдельные бородавки, валики, бугорки, локализованные в определенных местах на поверхности клетки или образующие общую орнаментацию. Выделение липкой слизи и образование слизистых головок на одном из полюсов клетки у отдельных видов Ankistrodesmus Corda, Pediastrum, Scenedesmus служат для прикрепления к субстрату или как временные приспособления при смене биотопа. У прикрепленных форм (Characium A. Br., Korschikoviella Silva) подушечки или пластинки слизи нередко пропитаны гидроксидом железа и окрашены в бурый цвет.
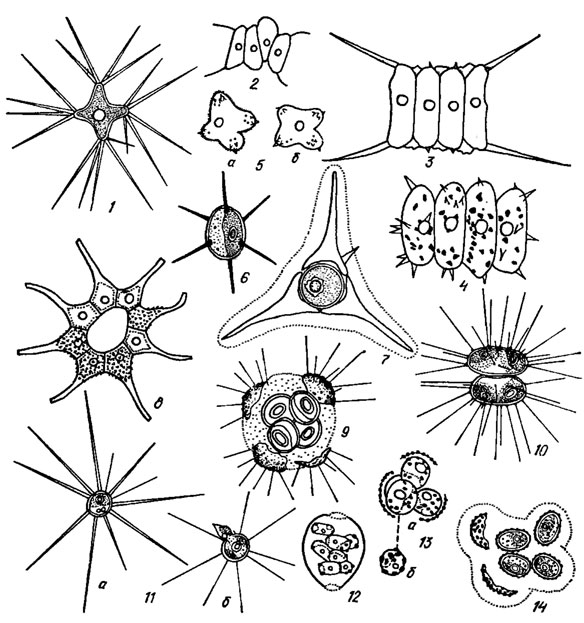
Рис. 19.17. Chlorococcales с разнообразными выростами, шипами, щетинками, зубчиками, бородавками на оболочке: 1 - Polyedriopsis spinulosa (Schmidle) Schmidle; 2 - Scenedesmus intermedius Chod.; 3 - S. communis Hegew.; 4 - S. pseudogranulatus var. hystricoides Mass.; 5 - Tetraedron minimum var. apiculato-scrobiculatum f. elegans Hortob. (а - с бородавчатой оболочкой, б - с гладкой оболочкой); 6 - Lagerheimia marssonii Lemm.; 7 - Treubaria planctonica Korsch.; 8 - Pediastrum simplex var. echinulatum Wittr. et Nordst.; 9 - Siderocystopsis fusca (Korsch.) Swale; 10 - Dicellula geminata (Printz) Korsch.; 11 - Golenkiniopsis solitaria (Korsch.) Korsch. (а - вегетативная клетка, б - яйцеклетка со сперматозоидом); 12 - Granulocystopsis coronata (Lemm.) Hind.; 13 - Siderocelis sphaerica Hind, (а - дочерние клетки с остатками материнской оболочки, б - вегетативная клетка); 14 - Siderocelis ornata (Fott) Fott
Слизь, окружающая клетки хлорококковых водорослей, либо продуцируется этими клетками (Radiococcaceae), либо образуется вследствие ослизнения остатков оболочки материнской клетки (Botryococcaceae, Sphaerocystidaceae). Остатки оболочки могут ослизняться довольно быстро, и тогда они практически не заметны в общей колониальной слизи (Coenococcus Korsch., Coenocystis Korsch. и др.), или сохраняться продолжительное время без изменения первоначальной формы клетки, или деформироваться (Coenochloris Korsch., Schizochlamydella Korsch.). В других случаях (например, Botryococcus Kütz.) остатки оболочки ослизняются в центре колонии, но отчетливо заметны на ее периферии или из них формируются слизистые прямые или разветвленные соединительные тяжи.
Ядро обычно занимает периферическое положение. Вегетативные клетки большинства видов содержат только одно ядро, реже несколько (например, 2-4 у Scenedesmus incrassatulus Bohl. и Nephrochlamys subsolitaria (G. S. West) Korsch.) или иногда являются многоядерными - т. н. ценоцитная структура (Hydrodictyon, Foilicularia Miller и т. п.). Ряд видов характеризуется непостоянным количеством ядер, которое зависит от возраста клетки или определенных отклонений в процессе размножения. Тонкая структура ядра хлорококковых полностью идентична таковой других эукариотических водорослей. В процессе митоза установлено наличие 4-32-80 (обычно 8-12) хромосом.
Хлоропластов - от одного или нескольких до многих; форма их различна. Наиболее часто встречается пристенный (периферический) тип хлоропласта в виде блюдца, корыта или чаши. Довольно редки спонгиоморфный (губчатый) и центральный, или осевой (звездчатый и лопастной) типы хлоропластов [510, 656].
Ультраструктура хлоропластов хлорококковых обычная для зеленых водорослей, так же как и состав основных фотосинтетических пигментов (хлорофиллы а и b). Однако существует группа бесцветных Chlorococcales, лишенных пигментов (Hyaloraphidium Pasch. et Korsch., Prototheca Kruger, Hyalocharacium Pasch.), которые являются, в принципе, аналогами хлорофиллсодержащих Ankistrodesmus, Monoraphidium Kom.-Legn., Chlorella, Characium.
Пиреноид расположен обычно в базальной утолщенной части хлоропласта, реже сбоку и только иногда строго в центре. Форма пиреноида может изменяться от сферической, овальной до почковидной или многоугольной, слегка неправильной. Для большинства видов характерно наличие одного пиреноида, реже нескольких (некоторые виды Characium, Korschikoviella, Eremosphaera Miller) или многих (Hydrodictyon, Coenococcus polycoccus (Korsch.) Hind.), но у некоторых они неразличимы в световом микроскопе и, возможно, отсутствуют (Heleochloris Korsch., Actidesmium Lagerh., Palmella Lyngb.). В целом пиреноид Chlorococcales состоит из основного вещества (стромы), нередко погруженной в него ламеллярной системы и часто окружающей его сплошной или фрагментарной обкладки, благодаря которой эта органелла хорошо заметна даже в живых клетках. Обкладка имеет крахмальную природу и характеризуется сильной преломляемостью света. Такая структура пиреноида характерна для большинства представителей порядка Chlorococcales, хотя существуют некоторые роды с т. н. открытыми пиреноидами - лишенными крахмальной обкладки с одной, обычно внутренней, обращенной к центру, стороны (Acanthosphaera, Golenkinia). Известны виды с голыми, трудноразличимыми в световом микроскопе пиреноидами, лишенными крахмальной обкладки (Kirchneriella Schmidle, Monoraphidium, Selenastrum Printz и др.), существование которых подтверждено в настоящее время методами электронной микроскопии.
Стигма, функционально связанная с хлоропластом, имеется у репродуктивных клеток Chlorococcales и, как исключение, у взрослых вегетативных клеток отдельных родов (Ankyra Fott, Characium, Bicuspidella Pasch.). Она расположена на переднем конце клетки в периферической части хлоропласта около клеточной оболочки. Размеры стигмы могут заметно колебаться даже в пределах одного рода (например, у Chlorococcum Meneghini).
Стигма функционально связана не только с хлоропластом, но и со жгутиковым аппаратом репродуктивных клеток Chlorococcales - зооспор, гамет. Для большинства зооспорообразующих форм характерны клетки с двумя жгутиками равной длины и только для некоторых (например, Pseudocharacium Korsch.) - с четырьмя. Тонкая структура жгутика полностью соответствует таковой других зеленых водорослей.
В периферической части короткого (до 2 мкм) базального тела жгутика располагается 9 триплетов микротрубочек (центральная пара отсутствует). Эти образования у хлорококковых ориентированы по часовой стрелке [547]. В подвижных клетках Pediastrum sp., Hydrodictyon reticulatum (L.) Lagerh. базальные тела соединены между собой перемычкой или (у зооспор Н. reticulatum) они направлены друг к другу под углом 180°. От базальных тел отходят несколько жгутиковых корней - сложная система собранных в пучки (по 5-8 у Hydrodictyon) микротрубочек [279]. Микротрубочки, при выходе из базальных тел, располагаются симметрично и образуют характерную для зеленых водорослей крестообразную фигуру. Пучки микротрубочек направлены к периферии клетки, к плазмалемме и выполняют роль цитоскелетных образований.
Сходны с базальными телами по внешней форме и тонкой структуре центриоли, которые обнаружены даже у родов, полностью утративших в жизненном цикле подвижную стадию (например, Kirchneriella, Chlorella Beijer., Scenedesmus, Tetrasdron Kütz.).
Размер и количество цистерн в диктиосоме (аппарат Гольджи) зависит как от функционального состояния клетки, так и от воздействия внешних факторов, однако колеблется в определенных пределах (например, 12-16 для Oocystis Näg.). Хлорококковые водоросли содержат обычно несколько митохондрий, хотя у некоторых наблюдается только одна (виды рода Chlorella). Число, размеры, форма и их расположение в клетке строго не фиксированы и колеблются в широких пределах в зависимости от возраста и функционального состояния клетки.
Цитоплазма пронизана системой канальцев эндоплазматического ретикулюма, которые местами расширяются, образуя цистерны или пузырьки с гладкой и шероховатой, несущей рибосомы мембраной. Разветвленная система канальцев наблюдается на периферии клетки вдоль плазмалеммы (например у Chlorella), либо вокруг ядра, образуя дополнительную перинуклеарную оболочку, которая сохраняется длительное время в процессе митоза у Hydrodictyon, Kirchneriella и Tetraedron. Помимо канальцев эндоплазматического ретикулюма клетки хлорококковых водорослей содержат микротрубочки, число и местоположение которых в клетке довольно изменчивы и зависят от ее функционального состояния. У Chlorella они возникают перед самым делением ядра и исчезают сразу же после завершения цитокинеза, а у Kirchneriella они сосредоточиваются на периферии клетки, непосредственно под плазмалеммой, в то время как у Hydrodictyon - около хлоропласта и располагаются поодиночке или собраны в группы по несколько в каждой. Из цитоплазматических телец (частиц) обнаружены пероксисомы (например, у видов рода Chlorella). Они состоят из мелкозернистого материала и содержат фермент гликолатдегидрогеназу.
Продукты ассимиляции - крахмал и масло, а у некоторых видов и вторичные каротиноиды. Крахмал концентрируется в основном вокруг пиреноидов, хотя отдельные зерна могут возникать также в строме хлоропласта между тилакоидами. Капельки масла иногда рассеяны в плазме клетки (Botryococcus, Trebouxia Puym., Spongiochloris Starr и некоторых видов Chlorococcum).
Деление клетки осуществляется с помощью клеточной пластинки, которая обычно появляется в плоскости клеточного деления в поздней анафазе или ранней телофазе между возникающими в ходе митоза дочерними ядрами. Она растет по направлению к периферии, разделяя протопласт на две части. У хлорококковых водорослей известно последовательное (например, Chlorococcum, Neochloris Star., Neospongiococcum Deas.), одновременное, которое нередко называют также делением распада (Characium,. Sorastrum Kütz.) и сегментационное деление (Spongiochloris, Planktosphaeria G. M. Sm., Dictyochloris Visch. et Star и др.). В результате цитокинеза] возникают совершенно обособленные одноклеточные особи, а оболочка материнской клетки, лишенная связи с протопластом, теряет способность к росту и под воздействием увеличивающихся в размере дочерних клеток разрывается. Разрушение оболочек происходит не спонтанно, а носит закономерный характер и зависит от количества и активности фермента целлюлазы.
Размножение происходит бесполым, реже половым путем, вегетативное деление отсутствует. Бесполое размножение осуществляется эндогенно образующимися бесполыми специализированными клетками - зооспорами, синзооспорами, гемизооспорами, апланоспорами и автоспорами. У некоторых родов установлена способность к размножению бесполыми клетками различных типов (при этом часто наблюдается значительное преобладание одного из типов), для других - только одного типа. Освобождение специализированных клеток происходит при полном или частичном местном ослизнении либо разрыве клеточной оболочки, а иногда ее распаде на две или несколько частей. У ценобиальных форм наблюдается формирование ценобиев в материнской клетке (Hydrodictyon, Scenedesmus) или после выхода из нее (Pediastrum, Euastropsis). Для некоторых родов или их отдельных видов (Chlorococcum, Tetraedron, Pediastrum и др.) характерно образование слизистого пузыря, в котором репродуктивные или сформированные дочерние клетки выходят из материнской.
Наиболее примитивные формы размножения Chlorococcales - активно подвижные зооспоры или ограниченно подвижные гемизооспоры. Количество этих клеток у отдельных родов и видов различно и иногда достигает нескольких десятков или сотен (например, у Chlorochytrium Cohn). Большинство зооспорообразующих родов характеризуются двужгутиковыми зооспорами и лишь некоторые (Pseudocharacium, Acanthosphaera) - четырехжгутиковыми. Зооспоры имеют различное морфологическое строение: одни из них лишены настоящей оболочки, т. е. голые - т. н. Dunaliella (-Carteria)-тип, что свойственно многим зооспорообразующим хлорококковым (Hydrodictyaceae, Golenkiniaceae, Sphaerocystidaceae и т. п.), другие - с прочной оболочкой, не метаболичны - т. н. Chlamydomonas-тип (Chlorococcum, Macrochloris Korsch., некоторые виды Characium) [405, 409, 510 и др.]. Гемизооспоры представляют собой в сущности недоразвитые зооспоры. Они лишены жгутиков, голые или покрытые оболочкой, прорастают на месте своего образования в материнской оболочке (Desmatractum W. et G. S. West, Trochiscia Kütz.) или выходят наружу в виде слегка амебоидновидных клеток и прорастают отдельно одна от другой (Treubaria Bern., Golenkinia, Acanthosphaera).
Неподвижные бесполые клетки размножения хлорококковых водорослей - апланоспоры и автоспоры - довольно сходны между собой и образуются в материнской клетке чаще всего в небольшом количестве, обычно по 4-8, значительно реже по 2, 16, 32 и 64, хотя апланоспор всегда формируется больше, чем автоспор [510]. Автоспоры у ценобиальных форм могут образовывать несколько или только один новый ценобий.
У колониальных и ценобиальных хлорококковых водорослей наблюдается иногда фрагментация или распад колоний или ценобия на отдельные части или одиночные клетки, что можно рассматривать как вегетативное размножение.
Половое размножение известно только у небольшого числа видов из различных семейств (Chlorococcaceae, Chlorochytriaceae, Dictyosphaeriaceae, Micractiniaceae и др.) и наблюдается довольно редко. Гаметы иногда сильно напоминают зооспоры. Среди хлорококковых водорослей, которым свойствен половой процесс, чаще наблюдается изогамия и значительно реже гетерогамия (Korschikoviella limnetica (Lemm.) Silva) и оогамия (Golenkiniopsis Korsch., Micractinium, Dietyosphaeriurn Näg.). Известны как гомоталличные (Chlorochytrium lemnae Cohn.), так и гетероталличные организмы (Hydrodictyon, Pediastrum, Micractinium, Golenkiniopsis, Dictyosphaerium).
В цикле развития хлорококковых зигота вскоре после слияния гамет переходит в состояние покоя и покрывается твердой оболочкой (гладкой, бородавчатой или шиповатой) и содержит большое количество запасных веществ. При прорастании зиготы происходит редукционное деление ядра и вегетативные клетки являются гаплоидными, реже оно отсутствует и тогда вегетативная стадия - диплоидная (Chlorochytrium lemnae). В последнем случае зигота прорастает сразу без слияния гамет, без периода покоя. У Golenkiniopsis, Micractinium наблюдались случаи партеногенетического преобразования гамет в вегетативные клетки.
При неблагоприятных условиях вегетативные клетки большинства хлорококковых способны превращаться в покоящиеся клетки - акинеты, цисты.
До настоящего времени нет единого мнения о систематическом статусе Chlorococcales и объеме этого порядка, не определена значимость отдельных признаков в систематике этих водорослей, существует большой разнобой в интерпретации видовых и внутривидовых таксонов, из-за чего многие из них трактуются разными авторами неоднозначно.
В зависимости от значимости, придаваемой коккоидной структуре таллома, хлорококковые водоросли рассматриваются в ранге класса, подкласса или порядка. В отечественной альгологии эта группа водорослей чаще всего представлялась как класс [65, 83, 205, 206, 233, 234, 330] или подкласс [161]. Зарубежные альгологи [355,356,359, 362, 417, 510 и др.] данную группу водорослей рассматривают в ранге порядка, что представляется и нам более обоснованным [318].
Тесные родственные связи Chlorococcales и Tetrasporales проявляются в наличии у некоторых видов рудиментарных признаков флагеллят - сократительных вакуолей и стигмы (Characiaceae, отдельные виды рода Chlorococcum) у вегетативных клеток. Существует также ряд родов, представляющих переходной тип между хлорококковыми и пакетовидными хлорсарциновыми, а также нитевидными улотриксовыми водорослями.
Систематика порядка Chlorococcales до сих пор остается искусственной, а разграничение на отдельные семейства - субъективным. За полтора столетия предложено свыше 70 классификационных систем, однако ни одна из них не является общепризнанной. Наиболее разработанной и обоснованной на данное время, с нашей точки зрения, следует признать систему И. Комарека, Б. Фотта [510], которая базируется на системах А. А. Коршикова [161] и Б. Фотта [417]. Данная система учитывает как традиционные морфологические признаки, так и современные данные по ультраструктуре и биохимии этих организмов. Принимая настоящую систему за рабочую, мы внесли в нее некоторые изменения с учетом новых данных по отдельным систематическим группам хлорококковых, а именно: 1) водоросли подсемейств Chlorosarcinoideae, Neochloridoideae р. р., Spongiococcoideae р. р., которым свойственно настоящее вегетативное деление клетки (десмосхизис) и своеобразный хлоросарциноидный тип структуры [198], относятся к порядку Chlorosarcinales, а подсемейство Disporoideae - к Ulotrichales; 2) оставшаяся часть видов подсемейства Neochloridoideae р. р. рассматривается нами в семействе Chlorococcaceae среди типовых представителей, а подсемейство Dictyochlorelloideae - в семействе Sphaerocystidaceae, в первом случае отказываясь от признака присутствия оболочки у зооспор, а во втором - признавая наличие зооспор, как указывал А. А. Коршиков [161]; 3) в соответствии с правилами "Международного кодекса ботанической номенклатуры" название семейства Palmellaceae изменено на Sphaerocystidaceae; 4) подсемейство Ankistrodesmoideae, на основании отличий в цитокинезе, в сравнении с другими представителями семейства Chlorellaceae, рассматривается в ранге самостоятельного семейства Selenastraceae, как ранее предлагалось [161, 543, 691]; 5) проведены соответствующие преобразования и перераспределение родов и видов в семействах Radiococcaceae, Oocystaceae, Chlorellaceae и Selenastraceae [462, 481, 543 и др.].
Хлорококковые водоросли распространены повсеместно. В экологическом отношении они довольно разнообразны. Преобладающее большинство видов - обитатели пресных или слабосолоноватоводных, мелких, хорошо прогреваемых, богатых биогенными веществами водоемов со стоячей или слабопроточной водой, лишь немногие встречаются в морях. Они заселяют водоемы различных типов - ручьи, реки, водохранилища, озера, пруды, в том числе биологические, болота, каналы, пойменные и эфемерные водоемы и различные их биотопы. Обнаружены они также на коре деревьев и среди обрастаний различных наземных материалов, в том числе и промышленных (капрон, пенопласт, стеклопластик или стекловолокно, сплавы алюминия, стали и т. п.).
Аэрофитные виды представлены в основном сферическими формами, из которых наиболее обычны представители родов Chlorococcum, Chlorella, иногда Trebouxia, Oocystis и др. Особое многообразие этих и подобных им родов наблюдается в почве, где они хорошо приспособились к неблагоприятным условиям (высушивание, промерзание, инсоляция и т. п.). Обычными представителями вневодных биотопов являются зооспорообразующие формы (например, виды родов Chlorococcum, Neochloris, Spongiochloris, Neospongiococcum). Наиболее обильно они населяют поверхностный слой почвы (0,2-1 см), но встречаются и на глубине до 2 м. С глубиной численность и видовое разнообразие хлорококковых резко уменьшается. Некоторые зооспорообразующие виды обитают на снегу (криофильные виды родов Chlorococcum, Chlorella, Oocystis Näg. em A. Br., Trochiscia, Scotiella Fritsch и др.) или в термальных источниках (отдельные виды рода Chlorella).
Большинство хлорококковых - свободноживущие формы и только некоторые - прикрепленные, имеющие или не имеющие специальных органов прикрепления (большинство видов Characiaceae, некоторые Selenastraceae, поселяющиеся на других водорослях, живых или отмерших частях высших растений, различных каменистых или бетонных субстратах), или эндофитные, либо эпизоитные, паразитические или симбиотические (например, Chlorochytrium lemnae развивается в клетках ряски, виды рода Chlorella - в клетках инфузорий, губок, мшанок, a Chlorochytrium, Kentrosphaera Borzi - внутри тела или слизи других водорослей). В последнем случае трудно установить четкую грань между симбиозом и паразитизмом, поскольку не всегда отчетливо проявляется повреждение клеток хозяина или другие явные признаки его угнетения. Виды родов Chlorella, Chlorococcum, Trebouxia, Myrmecia Printz обычно встречаются в теле лишайников, a Chlorella образует жизненное сообщество с плазмодием некоторых видов миксомицетов.
Обычно планктонные виды Chlorococcales обитают на глубине 0,5-1 м. Нередко они имеют специальные приспособления (щетинки, шипы, выросты и т. п.) для парения в толще воды (например, Micractinium, Golenkiniopsis, Golenkinia, Lagerheimia, Polyedriopsis). Этой же цели служат обширные массы слизи вокруг отдельных клеток или колоний (Coenochloris, Coenococcus Korsch., Coenocystis Korsch., Palmella и др.) или сочетание слизистой оболочки со щетинками или шипами, а также рыхлое строение ценобия, шаровидная, звездчатая его форма и т. п. Только обитатели поверхностной пленки воды (Ankyra ancora (G. М. Sm.) Fott, Chlorella neustonica Bourr., Emergosphaera superficialis Miller, Topaczevskiella nautococcoides Massjuk) снабжены специальными плавательными колпачковидными образованиями.
Наиболее благоприятны для развития хлорококковых водорослей пруды, особенно рыбоводные, где они являются одной из ведущих групп. Чаще всего в рыбоводных прудах наблюдается одновременная вегетация нескольких видов, создающих доминирующий ценоз конкретного водоема. "Цветение" воды могут вызывать виды родов Scenedesmus, Pediastrum, Crucigeniella Lemm. и другие, когда количество их клеток достигает 1-3 (до 5) млн/мл.
В целом, своеобразие экологических условий в каждом типе водоемов определяет характерный для него комплекс видов хлорококковых, однако свыше 40% видов этих водорослей встречаются на территории Украины практически в водоемах всех типов.
Хлорококковые водоросли играют большую роль в природе как продуценты органического вещества и кислорода. Они принимают активное участие в процессах самоочищения и формирования качества воды, а некоторые из них служат биоиндикаторами трофности и сапробности водоемов.
Хозяйственное значение многих видов хлорококковых определяется тем, что их поедают различные беспозвоночные и рыбы. Отдельные виды хлорококковых культивируют и используют в качестве стимулирующих кормовых и пищевых добавок, а также применяют в медицинской, парфюмерной и других отраслях промышленности.
Хлорококковые водоросли являются источником витаминов А, В, К, фолиевой и пантотеновой кислоты, холина и других витаминов (например, у видов рода Chlorella их насчитывают 15), характеризуются высоким содержанием полноценных белков (включая все необходимые для питания человека и животных аминокислоты, в том числе незаменимые) и других физиологически активных веществ. По содержанию белков, аминокислот и витаминов они не уступают другим источникам природных органических соединений. Например, пшеница содержит 13,8% белков, 66,6 углеводов и 1,8 жиров, мука гречневая - 8,5, 73,8 и 1,8 - соответственно, а суспензия хлореллы и сценедесмуса - 45; 35 и 12%. В качестве добавок к пищевому рациону сельскохозяйственных животных хлорококковые водоросли оказались особенно полезными в аридной зоне. Так, в Средней Азии их использование повышает продуктивность крупного и мелкого рогатого скота на 15-20% и снимает госсиполовое отравление; в птицеводстве - дает привес до 20%, увеличивает яйценоскость кур на 20-30% [224].
Лабораторные культуры этих водорослей используют в качестве удобного модельного объекта для выяснения механизмов дыхания и фотосинтеза, потенциальной продуктивности фотосинтетического аппарата, вопросов биологического саморегулирования и биосинтеза различных соединений, а также для решения некоторых других фундаментальных проблем физиологии, биохимии, генетики и т. п.
Согласно нашему пониманию порядка Chlorococcales, он подразделяется на 16 семейств; Chlorococcaceae (27 родов), Sphaerocystidaceae (20), Chlorochytriaceae (7), Dicranochaetaceae (1), Characiaceae (19), Treubariaceae (7), Hydrodictyaceae (4), Golenkiniaceae (4), Micractiniaceae (5), Botryococcaceae (13), Radiococcaceae (18), Chlorellaceae (24), Oocystaceae (30), Selenastraceae (17), Coelastraceae (6), Scenedesmaceae (29).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'