Морфология известковых сифоновых водорослей, терминология и методы изучения
Сифоновые водоросли, как и большинство других слоевцовых, сохраняются в осадках благодаря прижизненному обызвествлению. Степень обызвествления слоевища бывает самой различной, а от этого зависит полнота передачи морфологических признаков живой водоросли ископаемым формам. Обызвествление никогда не бывает полным и равномерным, поэтому какие-то детали всегда остаются неизвестными. Обычно минерализуются оболочки сифонов, периферических ответвлений, утрикул, спорангиев и т. д., а также пространство между этими морфологическими элементами. Как правило, наружные части слоевища минерализованы сильнее (за исключением дистальных ассимиляционных окончаний ветвей), а внутренние - слабее.
Палеоальголог, изучая сифоновые водоросли, имеет дело не со слоевищем, или талломом, как таковым, а всего лишь с его обызвествленной частью, минерализованным остатком, который несет то или иное количество информации о строении водоросли, но никогда не содержит всей информации. В этом принципиальное отличие сифоновых от синезеленых и красных водорослей, у которых обызвествление довольно полно передает детали строения слоевища.
Отмеченное обстоятельство требует серьезного внимания. Объект исследования, который мы видим в шлифе и считаем остатком сифоновой водоросли, может восприниматься и анализироваться двояко. С одной стороны, мы рассматриваем его просто как органический остаток с конкретными чертами строения, т. е. такой, каким мы его видим. С другой - зная систематическое положение этого остатка, можно мысленно как бы оживить саму водоросль и далее оперировать этим воображаемым объектом, отождествляя его с живым организмом.
На практике имеют место оба подхода. Какова доля реального и реконструктивного элементов в понимании ископаемых сифоновых водорослей и, следовательно, в их графической и словесной характеристике, зависит от индивидуальных особенностей исследователя, от биологического или палеонтологического (литологического) склада его мышления. Достаточно хорошо известно, что яркими представителями и убежденными сторонниками этих подходов (направлений) в изучении сифоновых водорослей являются крупнейшие отечественные палеоальгологи - К. Б. Кордэ и В. П. Маслов. Не имеет смысла обсуждать достоинства или недостатки разных подходов, поскольку они одинаково правомерны и должны гармонически дополнять друг друга. Однако следует подчеркнуть, что при палеонтологическом описании фактически наблюдаемое в шлифах должно быть четко отделено от предполагаемого.
До сих пор не выработана единая, схема описания ископаемых сифоновых водорослей. В. П. Маслов применял очень свободный стиль, называя обызвествленную часть слоевища " чехлом", "чехликом", "футляром", а в некоторых случаях вводил такие выражения, как "цилиндр", "палочка", "шарик", "колпачок", "трубка", "известковое тело" и т. д. Наиболее употребительным названием в его описаниях был "известковый чехол". Имеется в виду известковый чехол, облекающий слоевище. По сути дела, это выражение неточно, так как чехол включает только обызвествленную оболочку слоевища или эту оболочку и карбонатный материал, отлагавшийся на поверхности. Таким образом, чехол окружал не слоевище, а клеточное содержимое - протопласт. Ясно, что более "биологические" выражения типа "слоевище", "таллом", "обызвествленное слоевище", "известковое слоевище" ничем не лучше и в буквальном смысле могут употребляться относительно редко. Никаких возражений их использование не вызывает только тогда, когда речь идет о реконструкции растения.
Давно ощущается потребность в специальном обобщающем названии "обызвествленной части (остатка, реликта) слоевища". В качестве такового предлагается новый термин таллит, образованный от двух корней греческо-латинского происхождения: thallus - слоевище и lith(o) - камень. Синонимом может служить название таллоид. Выражением таллит (= таллоид) мы и будем пользоваться в дальнейшем изложении.
Форма таллитов сифоновых водорослей отличается значительным разнообразием (рис. III. 2). Цилиндрические или субцилиндрические таллиты могут быть простыми и разветвленными, массивными и трубчатыми, членистыми и нечленистыми. Субцилиндрические таллиты бывают сплюснутыми, а также монилиформными, т. е. снабженными регулярными внешними пережимами.
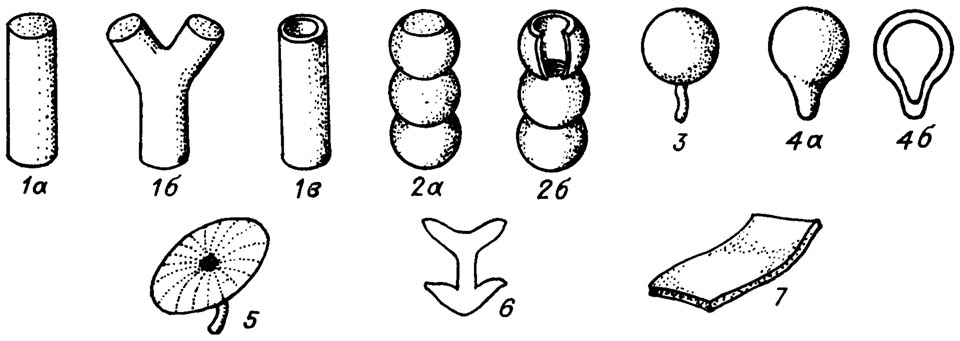
Рис. III. 2. Основные формы таллитов. 1 - цилиндрическая (а - простая стержневидная, б - разветвленная стержневидная, в - трубчатая); 2 - монилиформная (а - массивная, б - пустотелая); 3 - сферическая; 4 - грушевидная (клавиформная) (а - массивная, б - пустотелая); 5 - зонтиковидная (умбеллятная); 6 - ланцикулоидная; 7 - пластинчатая (ламеллярная, филлоидная)
Субсферические (циклокриноидные) таллиты бывают шаровидными грушевидными (клавиформными), яйце витыми (овоидными, овальными), массивными и пустотелыми. Относительно небольшую группу составляют зонтиковидные (умбеллятные) и сложнозонтиковидные многоярусные таллиты, характерные для ацетабуляриевых водорослей и образующиеся в результате особого обызвествления мутовок ответвлений. Столь же незначительно распространены пластинчатые, листовидные, или филлоидные, таллиты.
К особому типу относятся таллиты ланцикулоидного строения (род Lancicula Maslov). Они состоят из центрального стержня (рахиса) и серии расположенных друг над другом регулярных массивных расширений конической формы, которые мы именуем артикулами. Таллиты ланцикулоидного типа могут быть простыми, а также субдихотомически разветвленными.
В слоевищах сифоновых водорослей, а когда возможно, то и в таллитах, различают базальную и апикальную (вершинную) части. Во внутренней структуре слоевищ и таллитов в большинстве случаев четко выделяются осевая и периферическая зоны. Строение этих зон имеет важнейшее значение для систематики таксонов всех рангов - от вида до порядка.
Осевая зона (синоним - центральная, сердцевинная, медуллярная; медула) водорослей порядка Siphonales состоит из пучка трубчатых сифонов. В таллитах сифоны представляют собой цилиндрические продольные каналы, заполненные осадком (рис. III. 3); в пучке располагаются рыхло или тесно сомкнуты и имеют характер псевдопаренхимной структуры. Сифоны могут ветвиться и анастомозировать (срастаться), форма их может быть субцилиндрической, осложненной поперечными пережимами.
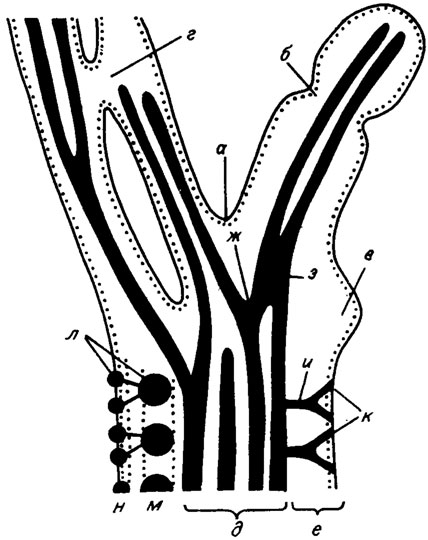
Рис. III. 3. Строение водорослей порядка Siphonales. а - ветвление слоевища; б - сегментация; в - пролификация; г - анастомоз слоевища; д - осевая (медуллярная) зона; е - периферическая зона; ж - ветвление сифонов; з - анастомоз сифонов; и, к - ветви первого и второго порядков; л - утрикул; м - коровый слой; н - вторичная кора (мантия)
Дазикладовые водоросли отличаются одноосевым, моносифонным строением. По оси таллита дазикладовой водоросли всегда проходит одна широкая или узкая трубка (рис. III. 4). У сифонокладовых центральная трубка снабжена поперечными перегородками типа днищ (рис. III. 5). Поперечные перегородки бывают сплошными, регулярными или нерегулярными, а также поясковыми, когда центральная часть перегородки не обызвествляется и на ее месте остается крупное отверстие.
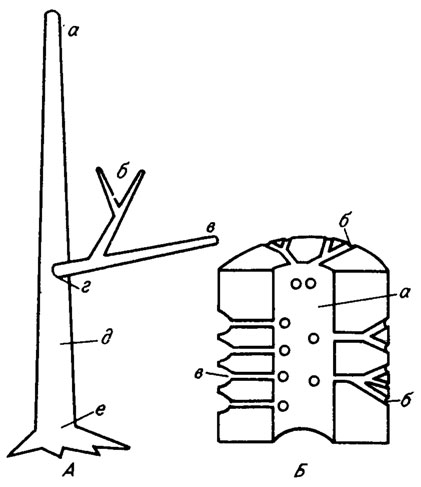
Рис. III. 4. Строение водорослей порядка Dasycladales. А - строение слоевища: а - апикальная часть слоевища, б - ветви 1-3 порядков, в - дистальная часть ветви, г - проксимальная часть ветви, д - центральная ось, е - базальная часть слоевища; Б - строение таллита: а - центральная трубка, б - периферические ответвления, в - коровый слой утрикул
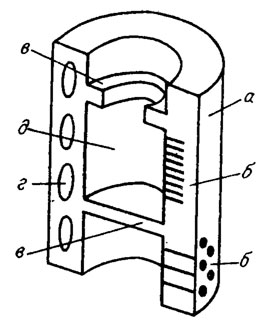
Рис. III. 5. Строение ископаемых сифонокладовых водорослей. а - стенка таллита; б - поры в стенках; в - поперечная перегородка; г - пометенные камеры; д - сегмент
Строение периферической зоны слоевища сифоновых водорослей отличается различной сложностью в зависимости от морфологии боковых ответвлений и характера их обызвествления. Дополнительный усложняющий элемент - образования типа утрикул и спорангиев.
Форма боковых ответвлений бесконечно разнообразна. Она может быть цилиндрической, гантеле- и каплевидной и т. д. Ветви бывают простыми или разветвленными. В этом случае различают ветви первого, второго, третьего порядка и т. д. (иногда до 5-7). У каждой ветви при изолированном рассмотрении выделяют начало - проксимальную - и окончание - дистальную части. В большинстве случаев минерализации подвергаются промежутки между ветвями. В результате на месте ветви остается простая или разветвленная пора в стенке таллита (рис. III. 6, III. 7). Пора обычно сквозная, поскольку в проксимальной части она сливается с осевой зоной слоевища, а в дистальной открыта наружу, так как ассимиляционные окончания не обызвествляются. Нередко каждая ветвь покрывается собственной известковой оболочкой по всей длине (см. рис. III. 6) или только в дистальной части (рис. III. 8). В этом случае ветви выступают на поверхность таллита в виде трубок и сосочковидных возвышений - папилл.
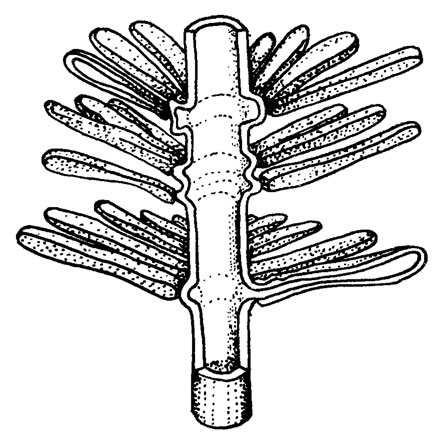
Рис. III. 6. Реконструкция ацетабуляриевой водоросли Masloviporella Kulik (по М. Р. Perret, D. Vachard /1977/)
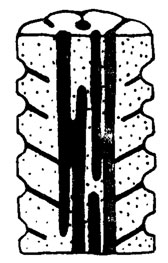
Рис. III. 7. Litanaia mira Masl. Принципиальная схема строения таллита
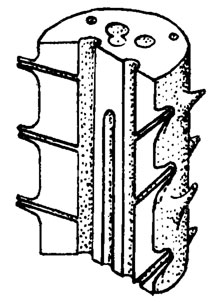
Рис. III. 8. Abacella Maslov. Реконструкция таллита
Распределение периферических ответвлений на поверхности слоевища бывает нескольких типов (рис. III. 9). Самый обычный - это беспорядочное (аспондильное) распределение. Оно наблюдается у многих собственно сифоновых и дазикладовых водорослей. Для дазикладовых водорослей характерно также мутовчатое (эуспондильное), поясково-мутовчатое и сложномутовчатое (метаспондильное) распределение. К сходным морфологическим типам относятся варианты распределения пор в трубчатых таллитах сифонокладовых водорослей. Поясково-мутовчатое распределение свойственно, например, для родов Beresella, Donezella, а метаспондильное - для рода Dvinella. Но в данном случае поры не открываются наружу, и поэтому трудно сказать, являются ли они слепками ответвлений или своеобразными структурами клеточных стенок.
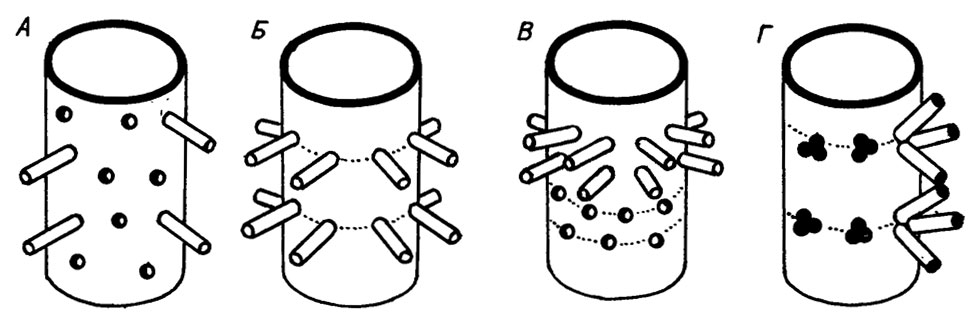
Рис. III. 9. Типы ветвления сифоновых водорослей. А - беспорядочное (аспондильное); Б - мутовчатое (эуспондильное); В - поясково-мутовчатое; Г - сложномутовчатое (метаспондильное)
Полисифонные формы ланцикулоидного строения обладают псевдомутовчатым характером развития ответвлений. Ветви в этом случае образуют периодические сгущения по длине слоевища. В результате их обызвествления образуются массивные конические артикулы. Внутри артикул располагаются спорангиеподобные камеры, поэтому их, по-видимому, можно считать спорангиофорами.
Большое систематическое значение имеет форма дистальных окончаний боковых ответвлений. Если ответвление было простым, волосовидным, то след его окончания сохраняется в виде цилиндрического отверстия в таллите или в форме известковой трубочки, возвышающейся над поверхностью (род Abacella).Довольно часто у сифоновых и дазикладовых ответвления заканчиваются шаровидными вздутиями - утрикулами. При тесном расположении образуется коровый (кортикальный) слой утрикул. В зависимости от степени обызвествления на их месте остаются сферические пустотки или полукруглые углубления на поверхности таллита (рис. III. 10) В тангенциальных сечениях кортикальный слой имеет ячеистое строение. От утрикул могут отходить волосовидные ответвления второго порядка. Если на дистальных окончаниях этих ответвлений вновь образуются утрикулы, то возникает вторичная кора, или коровая мантия (рис. III. 11).
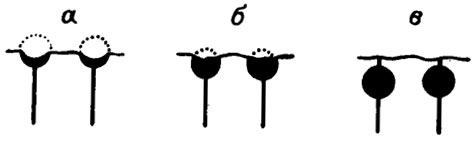
Рис. III. 10. Разная степень обызвествления и сохранности боковых ответвлений у водорослей Litanaia mira Maslov. а - обычная форма (точками показан внешний предполагаемый контур утрикула); б - форма обызвествления, встречающаяся относительно редко; в - встречается исключительно редко
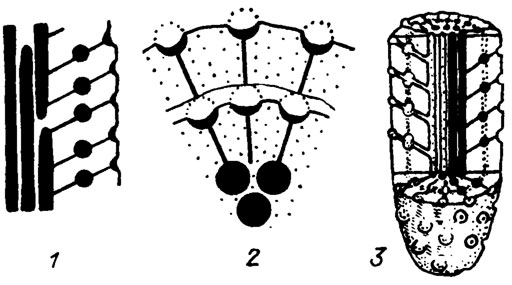
Рис. III. 11. Строение водорослей рода Circellet duplicata Schirschova (in press). 1 - строение таллита в продольном сечении (деталь); 2 - поперечное сечение; 3 - реконструкция одного членика таллита; нижний - средний девон, Урал
У типичных диклокриноидных водорослей боковые ответвления заканчиваются многоугольными пористыми пластинками, ориентированными перпендикулярно к ветвям. Смыкаясь краями, эти пластинки сливаются в "скорлупу" сферической, грушевидной формы и т. д.
Кроме описанного выше типа периферических мелких утрикул (см. рис. III. 10; III. 11), менее часто встречаются водоросли с крупными мешковидными утрикулами, расположенными перицентрально, т. е. непосредственно у медуллярной зоны (рис. III. 12). Такое строение характерно и для современного рода Codium (рис. III. 13).
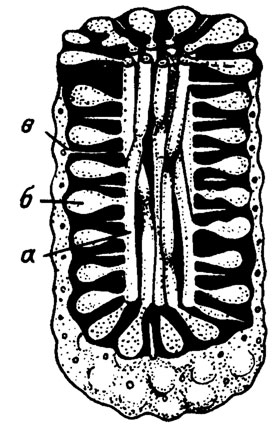
Рис. III. 12. Botrys compacta Schirschova (in press). Нижний - средний девон, Урал. Реконструкция таллита: а - сифоны; б - утрикулы; в - боковые ветвления
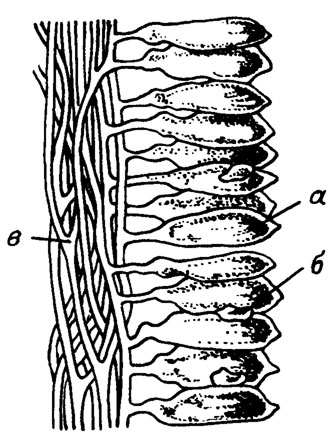
Рис. III. 13. Деталь строения современных водорослей рода Codium Sbackhouse. а - утрикулы; б - гаметангии; в - сифоны, образующие псевдопаренхиму (по К. Л. Виноградовой /Жизнь растений, 1977/)
В таллитах многих сифоновых водорослей встречаются сферические, грушевидные и другой формы пустотки, которые обычно интерпретируются как спорангии. По-видимому, правильнее рассматривать их как обычные морфологические элементы строения, поскольку генетическая их оценка всегда субъективна.
По всему комплексу различных морфологических признаков можно воссоздать внешний облик растения, наибольшую ценность имеют реконструкции таллитов. Операция реконструирования всегда неизбежна по той простой причине, что без этого морфологию таллита не описать. При достаточном количестве сечений в шлифах строение таллита восстанавливается однозначно и объективно.
Довольно часто пытаются воссоздать и само слоевище. Это, по-видимому, полезно, но использовать материал подобных реконструкций для целей систематики, как правило, нельзя, так как не исключены грубые ошибки. Поучителен в этом отношении пример с дазикладовыми водорослями рода Vermiporella Stolley. Таллиты вермипорел трубчатые, простые или ветвящиеся, иногда анастомозирующие с образованием трехмерных каркасов (V. fragilis Stolley). Стенки вермипорелл пронизаны многочисленными беспорядочно рассеянными порами. Предполагалось, что слоевище было очень простым и состояло из широкой центральной оси и коротких периферических ответвлений (рис. III. 14). Исследования Р. Козловского и Ю. Казьмерчака /Kozlowski, Kazmierczak, 1968 b/ показали ошибочность такой интерпретации. Оказалось, что слоевище вермипорелл состоит из относительно узкой центральной оси, от которой в виде мутовок отходят ветви трех порядков. Обызвествлению подвергались промежутки между короткими ответвлениями последнего порядка, в результате чего и образовывались известковые трубочки с пористыми стенками (рис. III. 15).
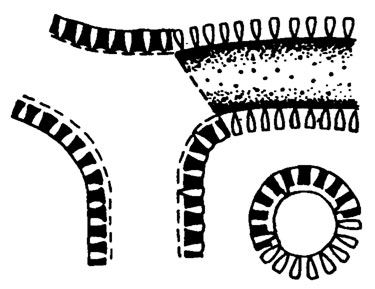
Рис. III. 14. Строение водорослей Vermiporella в интерпретации Дж. Пиа /Pia, 1920/
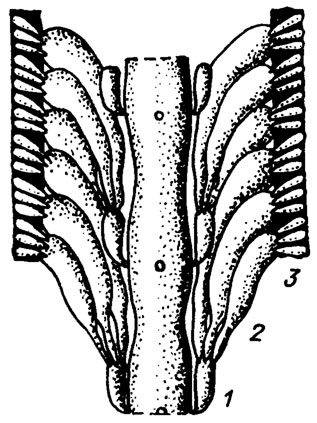
Рис. III. 15. Строение слоевища Vermiporella /Kozlowski, Kazmierczak, 1968/; видны центральная ось и ветви 1-3 порядков, расположенные в виде мутовок; черным цветом показаны обызвествленные промежутки между ответвлениями третьего порядка
В связи с этими уникальными данными приходится очень осторожно интерпретировать трубчатые таллиты других родов сходного строения (Rhabdoporella, Dasyporella, Anthracoporella). Как показали Р. Козловский и Ю. Казьмерчак, примитивное строение таллита еще не доказывает простоту строения слоевища.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'