Панцирь
Панцирь диатомей состоит из двух равных Половинок (подобно коробке, имеющей одинаковую с ней крышку). Одна половина панциря надвигается на другую, охватывая ее снаружи своими краями. Наружная половина панциря называется эпитекой, а половина, вдвинутая в эпитеку, - гипотекой. Эпитека и гипотека состоят из двух частей: створки и пояскового ободка. Створка обычно плоская (подобна дну коробки), края ее загнуты подобно бокам коробки и называются загибом створки, который бывает низким или высоким, занимающим до половины высоты боковой поверхности панциря. Поясковый ободок в виде широкой или узкой ленты окаймляет загиб створки и смыкается с ней, при этом свободным краем поясковый ободок эпитеки надвигается на свободный край пояскового ободка гипотеки и плотно охватывает его, но не срастается с ним. Границы поясковых ободков эпитеки и гипотеки панциря видны под микроскопом в виде двухконтурной полосы, называемой пояском панциря. В связи с таким строением панциря он имеет разный вид при рассматривании его со стороны створки и со стороны пояска. Известно незначительное количество родов, у которых поясковыс ободки отсутствуют, у них обе створки панциря плотно смыкаются непосредственно краями загиба створки (Melosira, Podosira, Hyalodiscus, Stephanopyxis, Annelus, Pyxilla, Endictya и др.). У этих диатомей поясок образуется во время деления клетки, которое протекает так же, как и у всех прочих диатомей. (Табл. II, 3а, 3в).
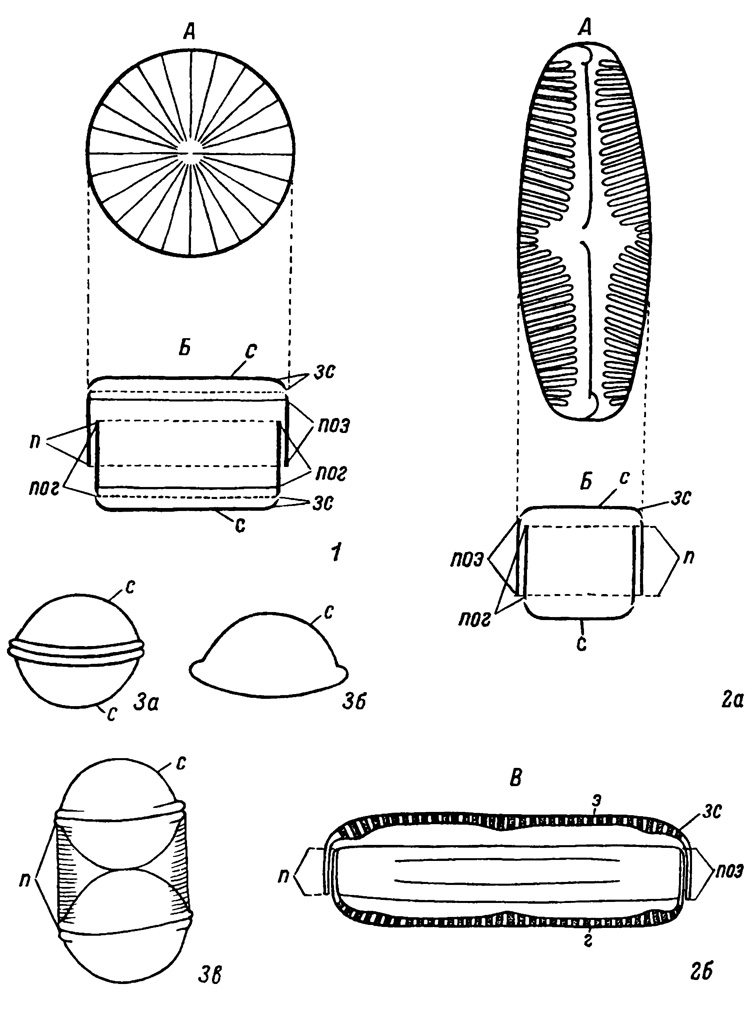
Таблица II. Морфология панциря. 1 - центричного типа, 2а, 2б - пеннатного типа, 3а-3в - центричного типа, не имеющего постоянного пояска. А - вид со створки, Б - поперечный разрез, В - вид с пояска, г - гипотека, зс - загиб створки, п - поясок, пог - поисковый ободок гипотеки, поэ - поясковый ободок эпитеки, с - створка, э - эпитека
У некоторых родов диатомей в процессе жизнедеятельности в панцире возникают дополнительные образования: вставочные ободки и септы. Вставочный ободок возникает между краями загиба створки и пояскового ободка, он обычно имеет вид узкой ленты, подобной поисковому ободку. Край вставочного ободка, граничащий с загибом створки, имеет бороздку, в которую вдвигается край загиба створки, а противоположный край ободка соединяется с поясковым ободком мельчайшими длинными зубчиками. Последующий вставочный ободок возникает на границе загиба и предыдущего вставочного ободка, иначе говоря, самый молодой ободок всегда граничит со створкой, а наиболее старый - с пояском. Биологическое значение вставочных ободков весьма существенно, они обеспечивают рост панциря, увеличивая объем его и протопласта, тогда как панцири, лишенные вставочных ободков, в течение своей вегетативной жизни имеют постоянный объем.*
* (Терминология деталей панциря и его структуры имеет важное значение при определении и описании диатомей, поэтому она помещена в виде терминологического словаря во втором томе этого издания, содержащего систематику диатомовых водорослей.)
Вставочные ободки по форме бывают различными, характерными для определенного рода. Кольцевидные ободки в виде ленты опоясывают загиб створки и своими заостренными концами, взаимно вклиниваясь, срастаются (роды Eucampia, Climacodium, некоторые виды Biddulphia, Corethron). Воротничковидные ободки подобны предыдущим, но они не смыкаются своими концами, а пространство между ними прикрывается выростом смежного, более молодого ободка (роды Guinardia, Lauderia, Detonula, Bacterosira, Schroederiella). Нередко вставочные ободки бывают не цельными, а состоящими из двух или нескольких члеников. Двучленистые, т. е. полу кольцевидные, ободки полуопойсывают панцирь и взаимно вклиниваются заостренными концами между собой (Leptocylindrus, Attheya, Dactyliosolen, Rhizosolenia longiseta, R. eriensis, Chaetoceros costatus). Вставочные ободки, состоящие из четырех и более члеников, имеют форму ромба, трапеции и чешуйки, они свойственны большинству видов рода Rhizosolenia. (Табл. III, 1-5). Вставочных ободков в эпитеке и гипотеке бывает от одного до очень многих. При делении клетки обе дочерние клетки получают по половине материнского панциря с принадлежащими ей вставочными ободками, а вновь образованные половины панцирей дочерних клеток не имеют ободков, они затем вырабатываются постепенно в течение жизни дочерних клеток.
У некоторых родов диатомей от внутренних стенок вставочных ободков в полость клетки вдается тонкая кремневая перепонка - септа, частично перегораживающая полость клетки. Септы образуются преимущественно у родов, имеющих удлиненные створки. У родов с гетеропольным панцирем септы обычно возникают на верхнем (широком) конце (Licmophora, Licmosphenia), а у изопольных - поочередно на обоих концах клетки (Striatella, Tetracyclics, Tabellaria). Септы бывают в виде очень узкой полоски на конце клетки или вдаются в полость клетки до ее половины. Реже септы возникают на вставочном ободке вдоль краев клетки, в этом случае края септ обычно смыкаются вдоль продольной оси панциря, оставляя одно или несколько отверстий (Rhabdonema, Grammatophora, Climacosphenia, Diatomella, Denticula, Epithemia). У рода Mastogloia боковые септы образуют камеры, которые трубчатыми порами связаны с внешней средой. (Табл. III, 6-12). Септы перегораживают клетку на полуизолированные отсеки, вследствие чего поверхность протопласта клетки сильно увеличивается, что ведет к увеличению поверхностного слоя цитоплазмы и количества мелких хлоропластов, расположенных в постепном ее слое, или увеличению изрезанности лопастей пластинчатых хлоропластов, проникающих в эти камеры. Жизненные преимущества клеток с септами совершенно очевидны.
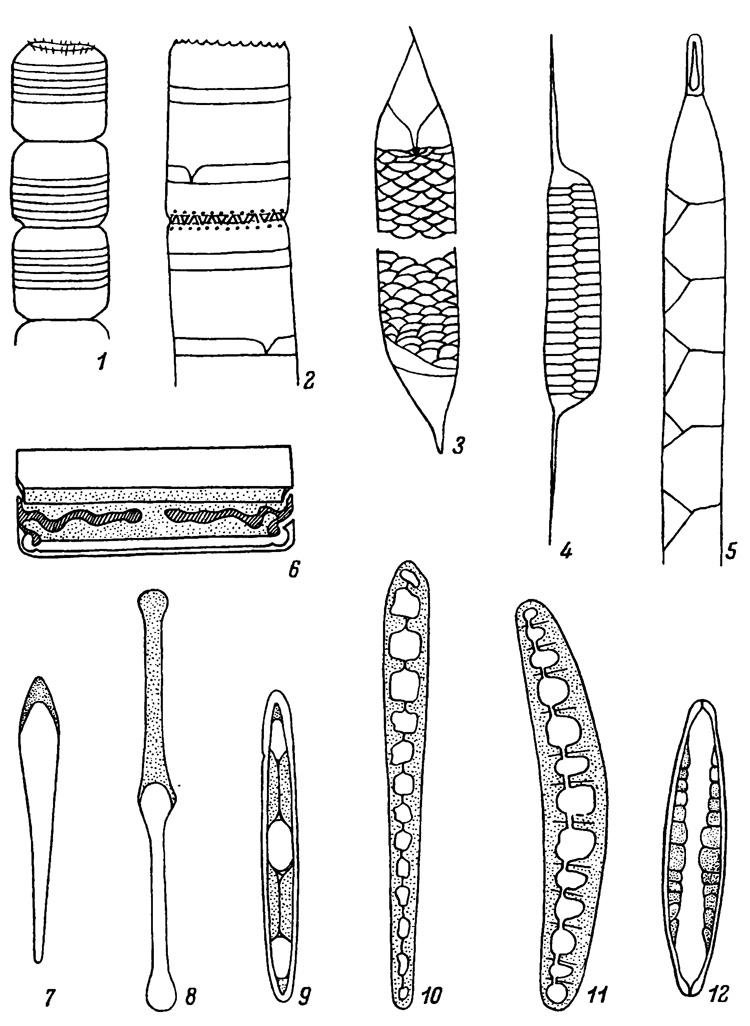
Таблица III. Вставочные ободки и септы (1 - по Gran, 1905; 7, 12 - по Прошкиной-Лавренко. 1963б; остальные - по Hustedt, 1927-1966, 1; 1930). 1-5 - формы вставочных ободков: 1 - кольцевидные у Lauderia borealis Gran, 2 - воротничковидные у Stephanodiscus binderanus (Kütz.) Krieger, 3 - чешуевидные у Rhizosolenia acuminata (Perag.) Gran, 4 - полукольцевидные у R. eriensis H. L. Smith, 5 - трапециевидно-чешуевидные у R. hebetata (Bright.) Gran; 6-12 - формы и положения септ: 6 - волнистая септа в продольном сечении у Grammatophora maxima Grun., 7 - полярная септа у Licmophora ehrenbergii Grun . 8 - септа, достигающая середины клетки у Tabellaria fenestrata (Lyngb.) Kütz., 9 - септа с тремя отверстиями и продольным швом у Rhabdonema adriaticum Kütz., 10 - септа со многими отверстиями и продольным швом у Climacosphenia moniligera Ehr., 11 - дорсовентральная септа с несколькими отверстиями и продольным швом у Epithemia argus Kütz., 12 - две постенные септы с камерами у Mastogloia pusilla Østr
Форма панцирей разнообразна, она тесно связана с формой створки. Панцири с круглыми створками имеют форму диска, высокого или низкого цилиндра. Панцири с продолговатыми створками: эллиптическими, ланцетными, линейной формы - имеют соответствующие очертания (табл. IV, 1-12). Концы створок также разнообразные (табл. IV, 13-18). Панцирь характеризуется преимущественно геометрической правильностью строения. Панцири с круглыми створками имеют радиальную (актиноморфную) симметрию. Панцири с эллиптическими, треугольными, ромбическими, линейными створками имеют билатеральную симметрию, реже панцири бывают асимметричными.
Стенки панциря тонкие, от 0.08 до 2.25 мкм. Они пронизаны мельчайшими отверстиями - перфорациями. Не существует панцирей, лишенных перфорированной структуры. Так называемые "гиалиновые", или "бесструктурные", панцири имеют очень тонкую пористую структуру, невидимую в оптические микроскопы, но обнаруженную электронным микроскопом (Chaetoceros, Leptocylindrus, Cerataulina, Detonula, Bacteriastrum, Attheya, Lauderia, Rhizosolenia и др.). На створках только некоторые участки с утолщенными стенками бывают лишены перфораций, это глазки, ставрос, осевое и центральное поле шва. К структуре панциря относятся также различного вида утолщения на панцире в виде складок, ребер и выростов.
Форма и структура панциря, в особенности его створок, имеет основное значение для систематики диатомовых водорослей. Структурные элементы на створке расположены различно: на створках центричного типа радиально, а на створках пеннатного типа перисто или поперечно, образуя штрихи. В зависимости от направления штрихов к краю створки они бывают параллельные, радиальные и конвергентные (табл. IV, 19-21), а по характеру штрихов - гладкие, точечные, линеолированные, редко в виде гладких ребер (табл. IV, 22-25). Характерными особенностями структуры являются постоянство ее расположения, правильная периодичность и количество структурных элементов у видовых и внутривидовых таксонов. Для диагностических целей количество элементов структуры исчисляют в линейной единице 10 мкм. В связи с правильностью расположения и периодичностью структурных элементов некоторые виды диатомей (Pleurosigma elongatum, P. angulatum, Amphipleura pellucida, Frustulia rhomboides) используются в качестве пробных объектов для определения апертуры микроскопических линз.
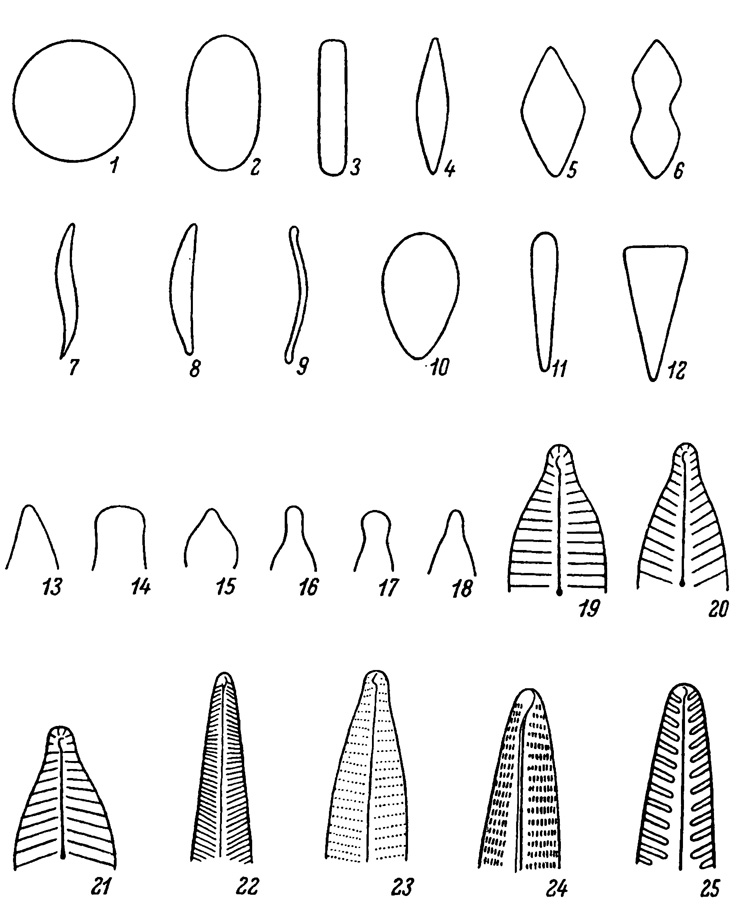
Таблица IV. Морфология створок. 1-9 - формы изопольных створок: 1 - круглая, 2 - эллиптическая, 3 - линейная, 4 - ланцетная, 5 - ромбическая, 6 - гитаровидная, 7 - сигмоидная, 8 - ладьевидная, 9 - серповидная; 10-12 - формы гетеропольных створок: 10 - яйцевидная (овальная), 11 - булавовидная, 12 - клиновидная; 13-18 - формы концов створок: 13 - острый, 14 - тупой, 15 - клиновидный, 16 - клювовидный, 17 - головчатый, 18 - оттянутый; 19-21 - расположение штрихов: 19 - параллельные, 20 - радиальные, 21 - конвергентные; 22-25 - характер штрихов: 22 - гладкие, 23 - точечные (пунктирные), 24 - линеолированные, 25 - ребровидные
Перфорированная структура панциря служит для сообщения протопласта клетки с внешней средой. Размеры перфораций структуры чрезвычайно малы, как показали вычисления, они обусловливаются осмотическим давлением в клетке. Характер структуры и расположения структурных элементов на створке, свойственный тем или иным видам, разновидностям и формам, находится, по-видимому, в прямой зависимости от физиологической деятельности клеток этих таксонов. Общая площадь перфораций на створке занимает от 10 до 75% ее площади.
Видимая в световой микроскоп структура створки представляется весьма разнообразной, но электронный микроскоп показал, что она состоит из небольшого количества основных элементов.
Использование электронного микроскопа для изучения тонкой структуры панциря и особенно его створки начато в 40-х годах (Kolbe, Gölz, 1943; Верцнер, 1944), и к 1959 г. было опубликовано свыше 100 работ, посвященных тонкой структуре панцирей и створок 300 с лишним видов, относящихся к 85 родам, причем многие виды были исследованы неоднократно (сводки: Desikachary, 1957; Okuno, Kurosawa, 1957; Hendey, 1959). К настоящему времени количество исследований сильно увеличилось. Помимо современных видов (Окидо, 1944-1968, 1951-1954 и др.), исследовались панцири ископаемых диатомей (Okuno, 1954-1959 и др.), появились специальные издания электронно-микроскопических микрографий, в том числе стереоскопических, посвященных структуре панциря и створки (Helmcke, Krieger, 1952-1970). В настоящее время известны детали структуры более 800 видов. Интерес к тонкой структуре створки возник после первых же ее исследований (Kolbe, Gölz, 1943; Kolbe, 1948- 1951, и др.) и последующих работ Дезикачари, Окуно и пр., показавших, что электронная микроскопия открыла детали структурных элементов, о которых при использовании оптического микроскопа нельзя было*и предполагать. Оказалось, что тонкое строение структурных элементов у многих генетически близких родов и даже у видов одного и того же рода часто было различным, и наоборот, у родов, относящихся к разным порядкам, встречались одинаковые структуры.
Это потрясло основы современной классической систематики - стало трудно принципиально обосновывать генетические соображения, казавшиеся ранее незыблемыми. В частности, с помощью оптического микроскопа различали структурные элементы в виде ареол у центричных диатомей и из точек (пор), образующих штрихи, - у пеннатных. Электронная микроскопия показала, например, что ребра Pinnularia - это те же ареолы, отличающиеся только формой. Ареолы свойственны, следовательно, не только центричным формам, но и пеннатным, поры также присутствуют у всех диатомей. Как поры, так и ареолы отличаются деталями своего строения, в связи с чем понадобилось их различать, поэтому были предприняты попытки классификации структурных элементов. Первоначально Дезикачари (Desikachary, 1952-1954, 1954) установил 6 типов ареол, позже на основании детального изучения их мембран он (Desikachary, 1957) добавил еще 4 типа. Окуно (Okuno, 1954-1959) различал 23 типа пор и ареол, а Хенди - 17 (Hendey, 1959). Было установлено существование двух основных типов строения кремневых стенок створки (Desikachary, 1957; Hendey, 1959): ламинарного типа - из одного кремневого слоя и локуларного - из двух слоев, между которыми находятся вертикальные перегородки, разделяющие пространство между двумя слоями на отдельные камеры - локулы, или ареолы. В стенках ламинарного типа перфорации представлены только простыми отверстиями - порами, а в локулярных стенках присутствуют ареолы и поры. (Табл. V).

Таблица V. Поры и ареолы (локулы) с мембранами разного типа (1, 5, 6, 9, 13, 15 - по Okuno, 1954-1959; остальные - по Helmcke, Krieger, 1952-1970). 1-4 - ареолы (локулы) круглые, шестигранные, линейные с круглыми порами, образующими ситовидную мембрану: 1 - Coscinodiscus lineatus Ehr., 2 - Actinopthychus undulatus (Bail.) Ralfs, 3 - Pinnularia major var. lacustris Meister, 4 - Achnanthes lanceolata Breb.; 5-8 - поры четырехугольные, округлые, продолговатые с выростами на их внутренней поверхности, образующими пересеченную отверстиями мембрану, которая рассматривается тоже как ситовидная: 5 - Cymbella mexicana (Ehr.) Cl. (миоцен), 6 - C. australica (A. Schmidt) Cl., 7 - Didymosphenia geminata (Lyngb.) M. Schmidt, 8 - Fragilaria brevistriata Grun. (миоцен). Масштаб 1 мкм; 9-12 поры круглые, полигональные, часто не вполне локуларные, с сетчатой мембраной разнообразного вида, у краев петли почковидной формы: 9 - Melosira granulata (Ehr.) Ralfs, 10 - Triceratium alternant Bail., 11 - Isthmia nervosa Kiitz. на загибе створки, 12 - то же на створке; 18-14 - ареолы (локулы) на калиптре четырехугольные и округло-четырехугольные с сетчатой мембраной (не в фокусе), с 2-4 круглыми сетчатыми порами по периферии: 13 - Rhizosolenia styliformis var. latissima Bright., 14 - Mastogloia pumila Grun.; 15 - Arachnoidiscus ornatus Ehr., ситовидная мембрана образована краевыми дихотомически разветвленными выростами; 16 - Navicula maculata var. inflata Okuno (миоцен-плиоцен), ареолы (локулы) разделены на ветви, основание широкоцилиндрическое, с мембраной из ситовидных пор. Масштаб 1 мкм, для 10-10 мкм
Поры - наиболее мелкие и простые структурные элементы, они представляют собой отверстие, пронизывающее стенку панциря, открытое наружу и внутрь клетки, диаметр пор 0.1-0.6 мкм. В толстых стенках панциря поры соответственно толщине стенок удлиняются в трубочки - поровые канальцы, которые иногда бывают сложными: начинаются снаружи стенки в виде простого канальца, а в толще стенки разделяются на 3 канальца, открывающихся в полость клетки. Часто наблюдаются полузамкнутые поры, у которых отверстие наружу открыто, а в полость клетки полузамкнуто. Среди них существует 5 типов. У первых трех типов отверстие в полость клетки закрыто тончайшей мембраной, пронизанной у первого типа круглыми отверстиями (ситовидная мембрана), у второго типа - квадратными (сетчатая мембрана) и у третьего - линейными (мембрана с насечками).Четвертый и пятый типы лишены мембраны, функции ее выполняют выросты на внутренней поверхности стенки поры: у четвертого типа древовидные, а у пятого-пластинчатые (Desikachary, 1957; Hendey, 1959; Drum, Pankratz, 1963b). (Табл. V).
Значительно сложнее устроены ареолы (локулы), образующиеся в локуларных стенках створок. Строение ареол независимо от их формы (круглой, многогранной, квадратной, удлиненной) можно свести к трем основным типахм: 1) ареолы, целиком открытые наружу створки и частично закрытые в полость клетки мембраной, пронизанной ситовидными порами или порами с насечкой (Actinoptychus undulatus, Melosira varians, Navicula viridula, Podosira stelliger и др.); 2) ареолы, открытые внутрь клетки и частично закрытые наружу мембраной с ситовидными порами, реже с пластинчатыми или древовидными выростами (Biddulphia tiiiana, Arachnoidiscus ehrenbergii, Isthmia nervosa); 3) ареолы, частично закрытые на обеих сторонах (наружной и внутренней) мембранами с порами различного типа (род Pinnularia, Rhizosolenia hebetata f. semispina, Stephanopyxis palmeriana, Coscinodiscus concinnus). (Табл. V).
Очень многие диатомеи имеют специализированные поры, выделяющие слизь, служащую преимущественно для скрепления клеток в колонии. Эти слизевые поры представляют собой канал, соединяющий полость клетки с наружной средой, размеры их крупные, форма разнообразная: круглая, эллиптическая, линейная. Располагаются слизевые поры у пеннатных форм на концах створки, у центричных - на середине створки (стр. 42).
Помимо перфорированной структуры существует структура, образованная местными утолщениями стенок створки на их внешней или внутренней поверхности в виде выпуклых ребер, высоких или низких складок, они придают прочность стенкам панциря. К структурным элементам относятся различного рода выросты на внешней поверхности створки: мелкие шипики, зубцы, длинные выросты в виде тонких щетинок или довольно толстых рогов. Они бывают сплошными или полыми внутри, иногда заполненными цитоплазмой или слизью, стенки их часто пронизаны порами, а некоторые снаружи имеют мельчайшие шипики. У некоторых видов через внутренний полый канал выделяется слизь, тогда этот канал по сути служит слизевой порой. Большинство выростов служит для соединения клеток в колонии и увеличения поверхности панциря. Эти структурные элементы присущи главным образом пелагическим морским диатомеям центричного типа.
Для многих диатомей пеннатного типа характерно присутствие на створке шва. Наиболее примитивный шов представлен двумя короткими прямыми, удаленными друг от друга щелями, расположенными по продольной оси створки (Rouxia, Amphipleura, Frustulia, Peronia) (табл. VI, 10-13). Такой шов произошел в результате удлинения щелевидной поры (Шешукова-Порецкая, 1956). Примитивный короткий щелевидный шов на обоих концах створки наблюдается у рода Eunotia, у многих видов он имеет хорошо выраженный конечный узелок, что ставит его генетически выше предыдущего шва. Кольбе (Kolbe, 1956) на обильном материале третичного вида E. eruca установил, что шов типа Eunotia произошел из ряда сближенных пор, слившихся вместе и образовавших короткую щель шва (табл. VI, 7-9). Несколько более развитый щелевидный шов по сравнению с предыдущими типами наблюдается у Cocconeis и Achnanthes, причем он находится только на одной из створок панциря: на эпитеке или гипотеке.
Наиболее высокоразвитый щелевидный шов присущ сем. Naviculaceae. Он одинаково развит на обеих створках панциря и тянется по продольной оси створки от одного ее конца до другого. Шов навикулоидного типа состоит из двух ветвей, т. е. двух длинных щелей, соединенных на середине створки центральным узелком и заканчивающихся близ концов створки конечным (полярным) узелком (табл. VI, 1). Щели ветвей шва коленчато изогнуты; щель, открывающаяся наружу, называется наружной щелью, а открывающаяся в полость клетки - внутренней щелью (табл. VI, 2-5). В центральном узелке обе ветви шва соединяются, наружная щель ветвей погружается в толщу створки, образуя воронковидный канал, которым соединяется с внутренней щелью своей ветви; обе ветви соединяются вместе с помощью горизонтального канала. В конечных узелках, представляющих собой Наружные и внутренние утолщения панциря, ветви шва оканчиваются. Наружная щель шва оканчивается в виде полулунной или слегка извитой терминальной (конечной) щели, открытой наружу, а внутренняя щель переходит в воронку, открытую в полость клетки. Конечные щели на одной створке панциря обычно изогнуты в одну сторону, а на второй створке - в противоположную сторону.
У некоторых семейств (Epithemiaceae, Nitzschiaceae, Surirellaceae) в складке стенки панциря имеется канал-шов, открывающийся наружу коленчато изогнутой щелью, а в полость клетки - рядом отверстий, называемых килевыми точками. Канал-шов также имеет центральный узелок, часто слабо выраженный или неразличимый (табл. VI, 14). Положение канала-шва на створке различно. Он бывает прямой и лежит более или менее эксцентрично к продольной оси створки (Denticula) или вдоль одного из краев створки (Nitzschia), иногда его ветви приближены к брюшному краю створки (Epithemia), или шов проходит по спинному краю створки (Rhopalodia), у родов Surirella и Campylodiscus шов тянется по краю крыла створки, поэтому он со стороны створки незаметен. Известен единственный род Cylindrotheca, у которого канал-шов спирально опоясывает панцирь.
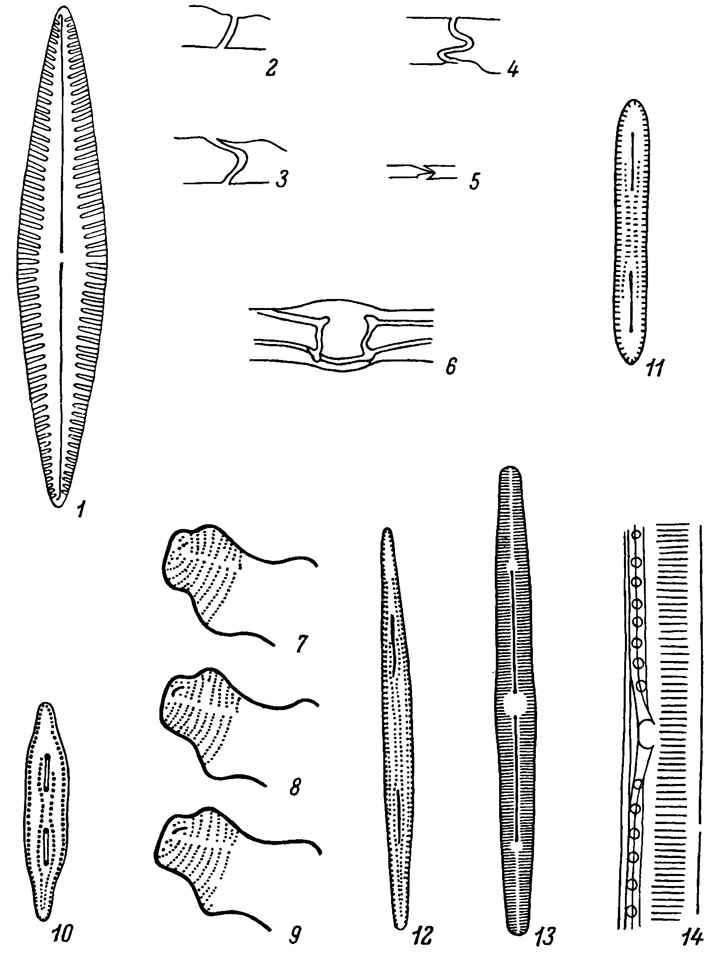
Таблица VI. Строение шва и его происхождение (1-6 - схемы; 7-9 - по Kolbe, 1956; 10-12 - по Шешуковой-Порецкой, 1956; 13-14 - по Hustedt., 1930). 1 - створка со щелевидным швом навикулоидного типа; 2-5 - щель шва в поперечном разрезе; 6 - центральный узелок в продольном разрезе; 7-9 - происхождение шва у Eunotia eruca Ehr. (третичный вид) путем слияния пор; 10-12 - происхождение шва навикулоидного типа путем удлинения щели у древних видов: 10 - Rouxia peragallii f. yabei (Hanna) Sneshuk., 11 - R. antarctica (Heid. et Kolbe) Hanna, 12 - R. peragallii f. califomica (M. Perag.) Sheshuk.; 13 - то же у современного вида Navicula brehhaensis Boye P.; 14 - канал обидный шов (средняя его часть) у Hantzschia amphyoxis (Ehr.) Grun
Шов свойствен более молодым филогенетически формам, виды со швом впервые появились в палеоцене и очень быстро прогрессировали. Вполне развитый щелевидный и каналовидный шов обеспечивает довольно быстрое движение клетки по субстрату и в толще воды, что, естественно, является большим жизненным преимуществом перед неподвижными формами.
Возникновению кремневой стенки панциря диатомей посвящен ряд электронно-микроскопических исследований.* Изучение серий ультратонких срезов (порядка 200-300 нм) делящихся клеток диатомей выяснило детали образования и роста кремневой створки и ее структуры. До настоящего времени в этом отношении детально исследовано 5 видов пеннатного типа: Gomphonema paruulum (Drum, Pankratz, 1964a), Amphipleura pellucida (Stoermer, Pankratz, 1964; Stoermer a. oth., 1965b), Cylindrotheca fusiformis, Navicula pelliculosa (Reimann a. oth., 1965-1966; Coombs a. oth., 1967, 1968) и Nitzschia alba (Lauritis a. oth., 1968).
* (Кремний (силиций, Si) - химический элемент, известный в виде трех устойчивых изотопов. Нами здесь употребляется наименование "кремневый", потому что химическое соединение кремния в природных водах и в панцире диатомей точно неизвестно, хотя часто называют кремневый панцирь диатомей "кремнеземным" (SiO2 - двуокись кремния).)
Эти исследования показали, что процесс образования створки происходит одинаково у всех вышеуказанных видов. В процессе деления диатомей, когда материнская клетка только что разделилась на две дочерние и протопласты дочерних клеток, еще будучи заключенными в материнском панцире, начинают слегка раздвигаться (в плоскости деления), в результате чего образуется узкое пространство между протопластами дочерних клеток, в этой стадии периферический слой цитоплазмы дочерних протопластов, уплотняясь, образует мембрану - плазмалемму (см. табл. I, 3, 4). Под плазма леммой, немедленно после ее образования, начинает формироваться новая створка, которая развивается синхронно с ростом и развитием молодой дочерней клетки, что свидетельствует о тесной связи образования структуры створки с физиологией клетки (Lewina. oth., 1966).
Процесс отложения кремния в возникающей створке детально прослежен у Amphipleura pellucida (Stoermer a. oth., 1965b) и у Navicula pelliculosa (Reimann a. oth., 1965-1966). Под плазмалеммой близ внутренней ее поверхности (на месте будущей створки) образуется трехслойная мембрана, в которой возникают пузырьки (вакуоли), где конденсируется кремний, поступающий из протопласта клетки. Трехслойную мембрану предложено называть силикалеммой (Stoermer a. oth., 1965b; Reimann a. oth., 1965-1966; Reimann, Volcani, 1967). Полагают, что силикалемма является активной избирательной перепонкой, проходя через которую кремний полимеризуется и отлагается в пузырьке (см. табл. I, 3, 4) в виде гомогенного тела, постепенно затвердевающего (Stoermer a. oth., 1965а, 1965b). Подобная трехслойная силикалемма наблюдалась у Cylindrotheca fusiformis, а осаждение кремния в пузырьках исследовано у Gomphonema parvulum и других видов; это лишь первые данные, и утверждать закономерность этого процесса для всех диатомей пока преждевременно. Под плазмалеммой сначала идет отложение кремния по продольной оси клетки в виде узкой полосы, посредине этой кремневой полосы формируется одновременно с ее возникновением щель будущего шва, щелевидного или каналовидного (см. табл. I, 3, 4). Край кремневой полосы постепенно расширяется вновь отлагающимся кремнием, при этом образуются структурные элементы створки, свойственные данному виду. Когда этот процесс достигнет стенок материнской половины панциря, края формирующейся дочерней створки загибаются, образуя загиб створки. Затем образуются стенки пояскового ободка и вновь образованная половина панциря дочерней клетки входит краями в материнскую створкуг которая всегда бывает эпитекой дочернего панциря (см. табл. I, 2). У большинства диатомей через 10-20 мин. после деления протопласта материнской клетки стенки створок дочерних клеток окончательно окремневают (Reimann, 1960, 1964).
Таков процесс возникновения кремневой створки диатомовых водорослей. Необходимо добавить, что в последние годы появились исследования, свидетельствующие о существовании у некоторых диатомей снаружи кремневой створки тончайшей органической пленки, названной кожицей (skin). Она плотно прирастает к створке, но может быть изолирована после растворения створки фтористым водородом в вакууме (Stoermer a. oth., 1965b). Предполагают, что эта кожица образуется при делении клетки из наружных остатков плазмалеммы и внешнего слоя силикалеммы, оставшихся снаружи створки после ее окремнения, а внутренний слой силикалеммы, оставшийся под кремневой створкой, по-видимому, в дальнейшем выполняет функцию мембраны протопласта, т. е. становится клеточной оболочкой. В стенках панциря цитоплазма отсутствует (Reimann a. oth., 1965-1966; Coombs, Volcani, 1968).
Кремний, из которого строится панцирь, находится в цитоплазме клетки в растворенном (неколлоидальном) состоянии, а вакуоли, в которых отлагается кремний, тесно связаны с клеточной цитоплазмой. Концентрация кремния в клетке диатомей колеблется от 4 до 50%, а вес панциря достигает 69% сухого веса клетки, толщина стенок створки колеблется от 0.08 до 2.25 мкм (Einsele, Grimm, 1938). Эти колебания зависят от вида диатомей (Jørgensen, 1953; Lewin, 1962, Lund, 1965). Но у одного и того же вида колебание содержания кремния в панцире связано с количественным содержанием его в воде водоема, в котором он обитает. Известно, что содержание кремния в природных водах сильно колеблется. В пресной воде большинства озер содержится растворенного кремния обычно меньше 5 мг/л, но бывает и выше: до 10 мг/л. Диатомеи могут переносить меньшее содержание кремния - 0.5 мг/л, но жизнедеятельность их при этом понижается. Опыты показали, что при недостатке кремния в культуральной среде образуются более тонкие панцири, уменьшаются темпы деления и активность ассимиляции в клетке, а при полном отсутствии кремния панцири совсем не образуются, однако беспанцирные клетки живут и даже делятся, а после внесения в среду кремния возникают клетки с нормальным панцирем (Bachrach, Lefevre, 1928, 1929, 1931; Bachrach, Pillet, 1930, 1931; Wiedling, 1941; Hendey, 1945, 1946). Йоргенсен экспериментально доказал растворимость кремния панциря при повышении в воде рН, так, например, панцирь Thalassiosira nana растворялся в воде при рН 10 в течение 37 дней на 97%. Он полагает, что кремний в панцире находится в форме, которая может гидролизоваться в различной степени. У одних диатомей кремний панциря легкорастворим, у других труднорастворим (Jørgensen, 1955b). Луин (Lewin, 1960) подтвердила способность кремневого панциря к растворению при рН 8-9 и повышенной температуре, указав особенности обмена веществ, происходящие при этом процессе. Вопросам усвоения диатомеями кремния и его растворению посвящено много исследований последнего времени.
Химический состав панциря изучался издавна. В прошлом столетии существовало мнение, что панцирь состоит из двуокиси кремния - SiO2. Позднее Либиш (Liebisch, 1929) указал, что его состав - гидрат окиси кремния, подобный опалу (SiO2+xH2O). Последующие исследователи установили в панцире около 2% примесей, состоящих из металлов: Аl, Fe, Mg (Metha a. oth., 1961). По поводу физического состояния кремния в панцире высказывались различные мнения. Одни исследователи полагали, что кремний в панцире находится в субколлоидальном и коллоидальном состоянии (Виноградов, 1939; Rogall, 1939), но другие указывали, что кремний панциря аморфный (Helmcke, 1954; Hendey, 1954), а у ископаемых панцирей он переходит в кристаллическое состояние. Исследования стенок панциря с помощью дифракции Х-лучей (рентгеновских) Доказали, что панцирь состоит из кристаллического кварца (одна из кристаллических модификаций SiO2) с добавлением в значительной пропорции органического компонента, который может быть протеином (Desikachary, Dweltz, 1961; Metha a. oth., 1961). Органический компонент (протеин) у ископаемых диатомей теряется,и кремний со временем все более и более кристаллизуется. Этим объясняется меньший удельный вес ископаемых панцирей (2.00) по сравнению с живыми диатомеями (2.07). Следует признать, что, несмотря на многочисленные исследования химического состава панциря, нет оснований считать его окончательно установленным.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'