Протопласт
Характерными особенностями протопласта диатомей, отличающими их от других микроскопических водорослей, являются отсутствие целлюлозной оболочки, крахмала, богатство жирами и сложный набор фотосинтезирующих пигментов. Протопласт диатомей включает цитоплазму, ядро, хлоропласта (хроматофоры),* вакуоли клеточного сока, продукты ассимиляции и мелкие органеллы клетки, оболочкой протопласта служит мембрана (плазмалемма). Цитоплазма расположена по периферии протопласта, у большинства форм, кроме того, она скопляется в центре клетки в виде цитоплазменного мостика, соединенного с периферическим слоем цитоплазменными тяжами. Клеточное ядро расположено в периферическом цитоплазменном слое, а в клетках, имеющих центральный цитоплазменный мостик, - посредине последнего. Хлоропласта лежат преимущественно в постенном слое цитоплазмы. Они бывают или мелкими зернистыми, многочисленными, или покрупнее - дисковидными, менее многочисленными, или пластинчатыми - от одной пластинки до нескольких; последние занимают обычно определенное положение в клетке. В крупных хлоропластах часто присутствуют пиреноиды - от одного до нескольких. В протопласте обычно наблюдаются в заметном количестве мелкие капли масла, являющегося продуктом ассимиляции и запасным питательным веществом, и капли волютина, в строме которых скопляется масло как запасной продукт. Протопласт окружен мембраной (оболочкой), тесно соприкасающейся с панцирем. (Табл. I, 1).
* (В настоящее время установлено тождественное строение хроматофоров водорослей и хлоропластов остальных растений, в связи с чем термип "хроматофор" отпадает.)
Протопласт диатомовых водорослей изучен, насколько позволяла разрешающая способность оптического (светового) микроскопа. Помимо всего сказанного были также известны процессы митоза, мейозиса и набор хромосом у некоторых диатомей (Fritsch, 1935). Детальное и критическое изучение цитологии и цитохимии протопласта диатомей с помощью электронного микроскопа открыло много нового в тонкой структуре его органоидов. Пока наши знания в области тонкой структуры диатомей ограниченны потому, что эти исследования проводятся только последние 5-6 лет, но несомненно в будущем надо ждать новых открытий в этой области. До настоящего времени электронно-микроскопическим методом исследованы протопласты только пеннатных диатомей, имеющих щелевидный и каналовидный шов: Nitzschia palea (Drum, 1963), Mastogloia grevillei (Stoermer a. oth., 1964), Gomphonema parvulum (Drum, Pankratz, 1964a), Amphipleura pellucida (Stoermer, Pankratz, 1964; Stoermer a. oth., 1965a, 1965b), Cylindrotheca fusiformis Reim. et Lewin, Navicula pelliculosa (Reimann a. oth., 1965-1966).
Исследования протопласта производились на серии ультратонких срезов клетки, показавших местоположение и тончайшую структуру органоидов клетки, детали которых до сих пор были неизвестны.
На ультратонком поперечном срезе (порядка 200-300 нм) в клетках навикулоидного типа хорошо видно ядро, имеющее Н-образную форму, оно окружено двойной пористой мембраной с порами диаметром 80-100 нм.* Через поры осуществляется обмен между ядром и цитоплазмой.
* (Единицы измерения для микроскопических и ультрамикроскопических объектов: мкм (микрометр, равен тысячной доле миллиметра), ммкм (миллимикрометр, равен стотысячной доле миллиметра), нм (нанометр, равен десятимиллионной доле миллиметра).)
Если ядро расположено близ хлоропласта, то оно прилегает к его поверхности непосредственно внешним слоем своей мембраны. Подобное расположение ядерной мембраны наблюдалось у многих диатомей (Gibbs, 1962; Stoermer a. oth., 1964; Stoermer a. oth., 1965b). Ядро наполнено ядерным соком, состоящим из нуклеопротеидов, гликопротеидов и ферментов. В нем образуются нуклеиновые кислоты, играющие основную роль в .... веществ, и сосредоточены дезоксирибонуклеиновая кислота (ДНК) .... генетическую информацию вида, и рибонуклеиновая кислота (РНК), принимающая участие в синтезе белка с помощью .... . В ядре находится хроматин, хорошо видимый во ...., расположены ядрышки (от 1 до 8), представляющие .... образования из всех клеточных органоидов. Вследствие .... собственной мембраны его границы не имеют рез .... клетки ядрышко растворяется как ограни .... метафазы митоза) и после окончания деления ..... молодой клетке. Ядрышко принимает участие в синтезе .... функция его не установлена. (Табл. I, 1).
.... диатомовых водорослей, как известно, чрезвычайно .... не форме, размерам и положению в клетке. До настоящего времени исследовано тонкое строение только крупных париетальных .... и нитцшиевых диатомей (Drum, 1962, 1963; .... Pankratz, 1964а, 1964b; Stoermer a. oth., 1964, 1965а). Их хлоропласты имеют пластинчатое (ламелларное) строение, свойственное прочим растениям. Пластинки плоские, тонкие, около 7-8 ммкм, они подобны тонким лентам и бывают одинарными, двойными и тройными. Пластинок в хлоропласте обычно от 4 до 7, между собой они тесно сближены, размеры их зависят от размеров хлоропластов. У диатомей навикулоидного типа пластинки широкие и длинные, так как их хлоропласты крупные, достигают длины клетки. Пластинки состоят из тончайших слоев белкового вещества и липидов, чередующихся между собой. (Табл. I, 1, 2). К пластинкам строго приурочены фотосинтетические пигменты, состоящие из смеси хлорофиллов, фукоксантинов и каротинов. Хлоропласты являются органоидами первичного синтеза органического вещества, вырабатываемого при участии световой энергии, т. е. элементарными фотосинтетическими единицами, при разрушении которых клетка теряет способность к фиксации СО2. Хлоропласт окружен тонкой однослойной мембраной, только у Cymbella affinis обнаружена двойная мембрана (Drum, Pankratz, 1964а, 1964b). У бесцветной гетеротрофной диатомовой Nitzschia alba ламелларные хлоропласты отсутствуют, но наблюдаются органеллы, которые были определены как цэопластиды, пигменты в них совершенно не обнаружены (Lauritis a. oth., 1968).
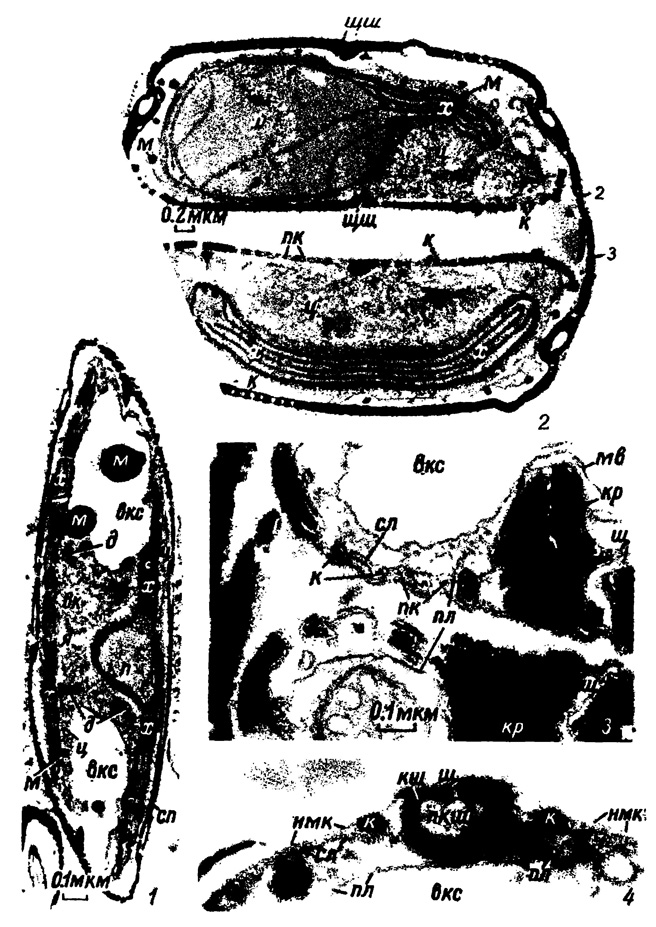
Таблица I. Ультраструктура протопласта клетки и панциря (1-3 - по Stoermer a. oth., 1965а; 4 - по Reimann a. oth., 1965-1966). 1-3 - Amphipleura pellucida Kütz.: 1 - продольный разрез клетки, прошедший чуть косо к ее средней плоскости, 2 - поперечный разрез только что разделившейся клетки в области расположения пиреноида, 3 - то же в области щелевидного шва образующихся створок дочерних клеток; 4 - поперечный разрез клетки Cylindrotheca fusiformis Reimann a. J. Lewin в области каналовидного шва (х 71 000), вк - везикулярный комплекс, вке - вакуоль клеточного сока, г - гипотека, д - диктиосома, к - кремний, кр - кремневое ребро, кш - каналовидный шов, м - масло, мв - мембрана вакуоли клеточного сока, нмк - неокремневшая мембрана клетки, п - пиреноид, пк - пузырьки, в которых отлагается кремний, пкш - полость каналовидного шва, пл - плазмалемма, сл - силикалемма, сп - стенка панциря, х - хлоропласт, ц - цитоплазма, щ - щель шва, щш - хцелевидный шов, э - эпитека, я - ядро
В хлоропласте навикулоидного типа имеется один пиреноид, расположенный близ середины хлоропласта и своей выпуклой поверхностью обращенный к периферии клетки, а плоской - к ее середине, он внедрен в хлоропласт, располагаясь среди его пластинок. У большинства диатомей пиреноид окружен собственной мембраной, хотя у некоторых диатомей не наблюдали. Структура пиреноида гомогенная, аморфная, у некоторых видов замечалась легкая гранулированность, некоторые исследователи наблюдали в пиреноиде скопления тубулярных пятен, состоящих из изогнутых трубочек, а иногда немного масла (Stoermer a. oth., 1965а). пиреноидах некоторых видов присутствовали дисковидные ленты (discand) от одной до трех (Gibbs, 1962; Drum, Pankratz, 1964b). Эти ленты двойные, своими концами прилегают к периферическим пластинкам хлоропласта, в связи с чем высказывалось предположение, что дисковидные элементы функционируют как избирательный механизм первичного или вторичного (Fogg, 1956) продукта фотосинтеза, транспортируя его в хлоропласт (Drum, 1963; Stoermer a. oth., 1964). В промежутках (локулах) между пластинками хлоропласта в процессе фотосинтеза появляются мелкие капельки масла, особенно многочисленны они вокруг пиреноида, но (в самом пиреноиде не откладываются. Было высказано мнение, что пиреноид принимает участие в образовании масла. Пиреноид подвержен изменению, способен делиться, он редуцируется в процессе деления клетки во время митоза или немедленно после его окончания (Drum, Pankratz, 1964а) и снова появляется во вновь возникшей клетке.
Цитоплазма клетки диатомей состоит из гиалоплазмы и эндоплазматической сети, которая пронизана массой вакуолей клеточного сока или несколькими большими, а иногда имеется только одна крупная вакуоль, оттесняющая цитоплазму к стенкам панциря. Цитоплазма у диатомей неподвижна. Некоторое движение цитоплазмы вызывается передвижением ядра при делении клетки и при образовании ауксоспоры, а также поворачиванием хлоропластов под влиянием света и при сокращениях вакуолей. Есть указание, что у планктонных пресноводных Rhizosolenia longiseta и Attheya zachariasii наблюдались сократительные вакуоли в период вегетации (Коршиков, 1930), а у Nitzschia hantzschiana (Pascher, 1932) и у Stauroneis phoenicenteron в период деления клетки. Было высказано предположение, что движение цитоплазмы совершается с помощью протеиновых фибрилл, образующих в цитоплазме динамическую систему. Их поперечная волнистость создает плавное движение цитоплазмы в противоположном направлении, и если фибриллы прикреплены к клеточным стенкам, то волны способны перемещать большие количества цитоплазмы (Haupt, 1962; Drum, 1963; Drum, Pankratz, 1964b).
В цитоплазме расположены многочисленные капельки масла, особенно много их скапливается, когда задерживается размножение клетки; это масло является запасным питательным веществом. Наряду с ними наблюдаются масляные тела, представляющие также скопления запасных питательных веществ, они увеличиваются в период фотосинтеза, идентичность их состава составу капель масла не доказана. Помимо этого у некоторых видов присутствуют еще капли волютина с характерным голубоватым блеском. В отличие от масла волютин нерастворим в эфире, а при окраске живой клетки метиленовой синькой принимает красновато- фиолетовый оттенок. Волютин в виде мелких капель скопляется в цитоплазме или в виде крупных капель (тельца Бючли) занимает обычно определенное положение на концах клетки или по обе стороны центрального цитоплазменного мостика. (Табл. I, 1). Во время деления клетки тельца Бючли исчезают, а после окончания деления появляются в дочерних клетках на прежнем месте. Их рассматривают как запасное вещество, состоящее из нуклеиновой кислоты, соединенной с белковой основой. Масло (жиры), экстрагированное из массы диатомей, имеет характерный запах рыбьего жира. У диатомей как запасное питательное вещество встречается также лейкозин (Коршиков, 1.930), называемый иногда хризозом. Лейкозин, по-видимому, углевод, при автолизе он дает глюкозу. В последнее время отмечена близость лейкозина к ламинарину, свойственному бурой водоросли Laminaria (Lewin, Guillard, 1963). В клеточном соке диатомей обнаружены ионы Cl, Na, К, Са, Mg, SO4, а также аминокислоты глицин, аланин, серин, треопин, валин, лепуин, фенилаланин, цистин, тирозин, пролин и многие другие (Low, 1955; Петрикова, 1959; Kesseler, 1967).
О мелких органоидах клетки диатомей, недоступных для исследования с помощью оптического микроскопа, было мало известно. Электронно-микроскопические исследования их у диатомей только начались, имеются некоторые сведения о митохондриях, аппарате Гольджи, пери- нуклеарных диктиосомах, центросоме и везикулярном комплексе.
Митохондрии (хондриосомы), имеющие вид скоплений палочковидных, нитевидных и зернистых элементов, располагаются всюду в цитоплазме, как близ ядра, так и в постенном слое и в плазменных тяжах. Шадефо (Chadefaud, 1939) указывает, что активные митохондрии расположены близ ядра, а неактивные - вдали от него. Митохондрии - органоиды внутриклеточного обмена, в которых сосредоточены ферменты, участвующие в синтетических процессах, протекающих в клетках, в них же вырабатывается энергия, которая в виде АТФ (аденозинтрифосфорной кислоты) поддерживает различные биологические процессы (Manton, 1961). В частности, цитохромы (белки, в состав которых входит железо) в митохондриях являются катализаторами реакции дыхательного цикла. Таким образом, митохондрии, перерабатывая вещества, поступающие в них, выделяют энергию, концентрирующуюся в соответствующих химических соединениях. Последние под действием биологических катализаторов - ферментов выполняют функции передачи. У Mastogloia grevillei митохондрии, концентрирующиеся в изобилии в камерах септ, по-видимому, вырабатывают материал, выделяемый через каналовидные поры камер (Stoermer a. oth., 1964).
Аппарат Гольджи расположен в цитоплазме вблизи ядра, имеет вид плотных скоплений, состоящих из мелких пузырьков, сеточки, палочек, чешуек, он белковой природы, в нем скапливаются вещества, вырабатываемые клеткой (Whalley, Mollenhauer, 1963). Тельца Гольджи присутствуют в клетках всех животных и принимают участие в их выделительной деятельности. Они найдены у немногих растений, но у диатомовых водорослей довольно обычны, однако их функция у диатомей не установлена. Этот органоид у диатомей не возникает вновь, а переходит в дочерние клетки в результате деления материнской.
Центросома наблюдалась у многих диатомей, по-видимому, она является постоянным клеточным органоидом (Lauterborn, 1896; Drum, 1963; Drum, Pankratz, 1964a). Она всегда расположена в небольшой впадине на одной из сторон ядра. Исследование центросомы у Surirella ovalis (Drum, Pankratz, 1963a) показало, что она сферическая, около 500 ммкм в диам., содержимое ее густозернистое, без типичных центриолей. По периферии центросома имеет нити около 15 ммкм толщиной, которые особенно хорошо представлены в стадии интерфазы, и кажется, что свободными концами они направлены к порам ядерной мембраны, но в ядре не наблюдались. Участие центросомы в кариокинетическом делении клетки диатомей известно давно, впервые это наблюдал Лаутерборн (Lauterborn, 1896).
Перинуклеарный диктиосомный комплекс, представленный скоплениями уплотненного аморфного вещества, всегда расположен вблизи ядра. Диктиосомы давно были известны исследователям (Pfitzer, 1871; Lauterborn, 1896; Hustedt, 1930; Geitler, 1952b, и др.). Высказывалось предположение, что эти органеллы представляют запасное питательное вещество. Однако изменение их во время деления ядра (Hustedt, 1930; Gschopi, 1952; Whalley, Mollenhauer, 1963) вызвало подозрение об их участии в образовании веретена и делении их во время митоза без потери собственной целостности. Однако определенного мнения о функции перинуклеарных диктиосом до сих пор еще нет. (Табл. I, 7).
Некоторые исследователи (Stoermer a. oth., 1964, 1965а) описывают у Amphipleura pellucida и Mastogloia grewillei органеллы трубчатой структуры, названные ими везикулярным комплексом. Эти исследователи указывают на присутствие этого комплекса у диатомей подпорядков Monoraphidineae и Biraphidineae, у которых они располагаются на поверхности цитоплазменной массы. У Mastogloia они примыкают к камерам септ. Предполагают, что везикулярный комплекс функционирует как запасное питательное вещество, потому что во время деления он редуцируется.
В начале 60-х годов в цитоплазме у животных и высших растений открыты очень важные органеллы - рибосомы, состоящие из мельчайших шариков, часть которых как бы нанизана на нити. В них при участии ферментов, сцепляющих аминокислоты между собой, происходит синтез белка. В связи с чрезвычайно важной функцией рибосом следует предполагать, что они имеются и у водорослей, в частности у диатомей.
Протопласт диатомей заключен в тончайшую мембрану, происхождение которой было выяснено электронно-микроскопическими исследованиями. Эту мембрану можно видеть в процессе деления клетки, когда материнская клетка только что разделилась на две дочерние; у последних наружный слой цитоплазмы уплотняется, образуя мембрану - "плазмалемму" (Stoermer a. oth., 1965а; Reimann a. oth., 1965-1966). Плазмалемма окружает протопласт, тесно соприкасаясь с панцирем, и проникает во все его полости, открытые вовнутрь клетки; она является той растительной перепонкой, через которую происходит обмен протопласта клетки с внешней средой. (Табл. I, 3, 4). Химическая природа плазмалеммы точно не установлена. Существовавшее предположение, что она пектиновая (Liebisch, 1929) или протеиновая (Desikachary, Dweltz, 1961), в настоящее время многими отрицается.
Мембранные образования, открытые в течение последнего десятилетия, тщательно изучаются. Выяснено, что они не являются структурно-химическим препятствием для движения веществ (Хокин Л, Хокин М., 1967). Проницаемость мембран у диатомей для неэлектролитов выше, чем у других растительных клеток, но она не у всех диатомей одинакова. Некоторые диатомеи имеют высокую проницаемость для Сахаров, у других она замедленная, а у Caloneis obtusa и Cymbella aspera мембрана почти непроницаема для Сахаров. У галофильных видов диатомей проницаемость для солей очень высокая.
Эти краткие сведения о тонкой структуре протопласта диатомей надо рассматривать как предварительные, они требуют дальнейшего исследования и дополнения, а некоторые данные и проверки.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'