Изменения количества видов и численности почвенных водорослей в течение вегетационного периода
Сезонная динамика водорослей, т. е. колебания их состава и численности, обусловленные сезонными изменениями экологических факторов, является реальным фактом, показанным многочисленными исследованиями. Они обобщены в трех монографиях [Голлербах, Штина, 1969; Рийс, Рахно, 1975; Штина, Голлербах, 1976а]. Однако имеющиеся материалы содержат немало противоречий и не позволяют сделать однозначный вывод о влиянии сезонных факторов, в частности, температуры и влажности почвы, на развитие почвенных водорослей. Противоречия эти следующие. Во-первых, сезонные колебания численности и биомассы водорослей более значительны в пахотных почвах, чем в рядом расположенных целинных почвах [Штина, 1959а; Помелова, 1971; Бусыгина, 1975; и др.] . Во-вторых, сезонные изменения оказались очень различными в разных почвах, и максимум численности падает на разные сезоны года. Однако эти различия не укладываются в какую-нибудь закономерность, например, в представление о зональных особенностях. Для почвенных водорослей нет "зонального" типа периодичности, известного для фитопланктона. Точно так же пределы колебаний численности и биомассы водорослей, как показала сводка Штиной и Голлербаха [1976а], очень сходны в разных зонах. Наконец, развитие водорослей в почве очень динамично, по-видимому, более динамично, чем развитие гетеротрофных микроорганизмов.
Основной фактор, контролирующий численность водорослей - влажность почвы, - подвержен резким колебаниям и имеет не только сезонную динамику, но и более дробную, изменяясь как от месяца к месяцу, так и день ото дня и даже по часам суток. Соответственно обнаружены резкие и часто кратковременные изменения численности и биомассы водорослей в пределах нескольких дней [Домрачева, 1974; Маркова, 1976; Кабиров, Минибаев, 1978].
Из многочисленных исследований, касающихся динамики почвенных водорослей, лишь в нескольких содержатся данные о периодичности развития водорослей в лесных почвах [Штина, 1959а; Байрамова, 1965; Куликова, 1965; Штина и Ройзин, 1966; Новичкова Иванова, 1969; Алексахина, 1974-1982; Чаплыгина, 1977].
Изучение сезонных изменений флоры почвенных водорослей в коренных типах леса проводилось нами в течение двух вегетационных периодов.
Таблица 12
| Тип леса | Отдел водорослей | 1-й год исследований | 2-й год исследований | ||||
| начало лета | лето | осень | весна | лето | осень | ||
| Ельник-черничник | Cyanophyta | - | 2 | 2 | 1 | 2 | 3 |
| Chlorophyta | - | 31 | 30 | 18 | 21 | 24 | |
| Xanthophyta | - | 7 | 9 | 4 | 8 | 8 | |
| Bacillariophyta | - | 0 | 1 | 0 | 1 | 2 | |
| Общее число видов | - | 40 | 42 | 23 | 32 | 37 | |
| Дубо-ельник волосистоосоковый | Cyanophyta | 5 | 8 | 5 | 3 | 5 | 7 |
| Chlorophyta | 26 | 30 | 33 | 20 | 25 | 27 | |
| Xanthophyta | 17 | 11 | 19 | 10 | 12 | 15 | |
| Bacillariophyta | 0 | 0 | 1 | 1 | 2 | 3 | |
| Общее число видов | 48 | 49 | 58 | 34 | 44 | 52 | |
| Сосняк с елью чернично-зеленомошный | Cyanophyta | 2 | 3 | 4 | 2 | 2 | 4 |
| Chlorophyta | 22 | 23 | 23 | 16 | 23 | 19 | |
| Xanthophyta | 5 | 6 | 12 | 17 | 12 | 10 | |
| Bacillariophyta | 0 | 0 | 1 | 1 | 0 | 1 | |
| Общее число видов | 29 | 32 | 40 | 26 | 37 | 34 | |
| Снытево-осоковая ясеневая дубрава | Cyanophyta | - | 9 | 12 | 12 | 6 | 15 |
| Chlorophyta | - | 19 | 21 | 33 | 15 | 19 | |
| Xanthophyta | - | 9 | 12 | 12 | 16 | 10 | |
| Bacillariophyta | - | 1 | 3 | 2 | 2 | 5 | |
| Общее число видов | - | 38 | 48 | 59 | 39 | 49 | |
Наблюдавшиеся сезонные изменения количества видов и численности водорослей носят неодинаковый характер в разных типах, леса и почв, а также в различные годы. Одной из черт сезонной динамики водорослей в лесных почвах является изменение в течение сезона числа видов водорослей разных отделов (табл. 12).
В самом северном из рассмотренных типов леса - ельнике-черничнике - возрастание числа видов водорослей идет постепенно от весны к осени. Заметно увеличение видового разнообразия в подстилке, особенно в ее верхнем 1,5-сантиметровом слое, где отмечены и наиболее значительные колебания численности водорослей с максимальным количеством клеток летом и минимальным - весной (рис. 5). Превышение летней численности водорослей над весенней и осенней составляет в этом слое подстилки 53 и 35% соответственно. В почве наблюдается тенденция к повышению количества водорослей к осени.
Ни в подстилке, ни в почве развитие водорослей не обнаруживает прямой зависимости от влажности в момент отбора проб. Напротив, значительное возрастание влажности подстилки в осенний период, связанное с выпадением атмосферных осадков, совпадает со снижением здесь численности водорослей. С одной стороны, это может быть обусловлено вымыванием клеток водорослей током воды в нижележащие слои почвы. Действительно, в ельнике-черничнике в дождливые периоды наблюдается некоторое увеличение обилия и видового разнообразия водорослей на глубине 5 - 15 см. С другой стороны, несоответствие изменений влажности и количества водорослей может быть обусловлено также усиливающимся влиянием других факторов. Так, при достаточном увлажнении действие влажности на численность водорослей проявляется через усиление поглощения питательных веществ, изменение ростовых процессов у высших растений [Штина, 1959а], через активность беспозвоночных [Домрачева, 1974] и т.д. По мнению Шилдс и Даррелла [Shields, Durrell, 1964], в тех случаях, когда вода не является лимитирующим экологическим фактором, богатство альгофлоры определяется почвенным плодородием.
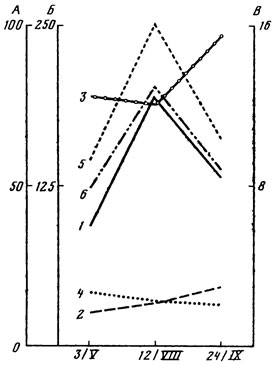
Рис. 5. Сезонные изменения количества водорослей в подстилке и почве ельника-черничника. А - число клеток (тыс. в 1 г абс. сухой почвы или подстилки); Б - влажность почвы или подстилки, %; В - температура, °С. 1 - общее число клеток в подстилке; 2 - общее число клеток в верхнем 5-сантиметровом слое почвы; 3 - влажность подстилки; 4 - влажность почвы; 5 - температура подстилки; 6 - температура почвы
На осеннее падение количества водорослей, вероятно, влияет и снижение температуры. На рис. 5 довольно ясно прослеживается прямая зависимость численности водорослей от температуры подстилки. Факты прямого влияния температуры на количество водорослей указывают также Э. А. Штина [1959а, 1974]; Ш. У. Умарова [1962]; Г. И. Маркова [1974]; Л. Н. Новичкова-Иванова [1974].
По данным Г. Н. Перминовой степень корреляционной связи между численностью водорослей в пятнах тундры и температурой почвы достигала r = 0,51, а температурой воздуха - r = 0,63.
В течение вегетационного периода происходит некоторая перестройка в составе преобладающих видов водорослей. К осени увеличивается разнообразие состава доминирующего комплекса. Так, если в весенне-летний период в верхнем слое подстилки (0 - 1,5 см) наиболее обильной была Coccomyxa solorinae, то осенью к ней присоединяется еще ряд одноклеточных (Chlorella vulgaris, Muriella magna) и нитчатых (Stichococcus minor, Leptosira polychloris) зеленых водорослей, а также некоторые виды Xanthophyta. Встречающиеся в нижнем слое подстилки (1,5 - 6 см) и в верхних слоях почвы (6 - 10 и 10 - 15 см) хламидомонады становятся особенно обильными тоже в осеннее время. Усиление вегетации желто-зеленых водорослей в подстилке отмечается чаще всего в периоды с повышенной влажностью, преимущественно осенью после дождей, что еще раз указывает на их требовательность к влаге.
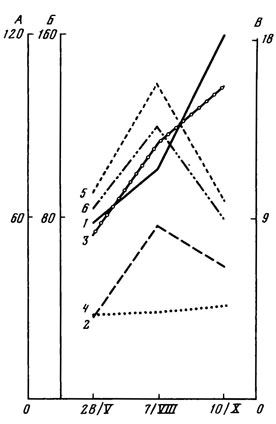
Рис. 6. Сезонные изменения количества водорослей в подстилке и почве дубо-ельника волосистоосокового. Обозначения те же, что на рис. 5
В дубо-ельнике волосистоосоковом в весенний период в подстилке и почве выявлено самое низкое число видов (см. табл. 12) и численность водорослей (рис. 6). Влажность подстилки в этот период была довольно высокой, а почвы - близкой по величине к влажности в два других сезона. Одним из факторов, тормозящих развитие водорослей в весеннее время, являются, по-видимому, значительные понижения температуры ночью вплоть до заморозков. Рост влажности подстилки сопровождается максимумом количества водорослей в осенний период. В почве наибольшая численность водорослей приходится на лето, при этом колебания количества клеток довольно четко повторяют изменения температуры. В более влажном году число видов водорослей было несколько выше, чем в более сухом.
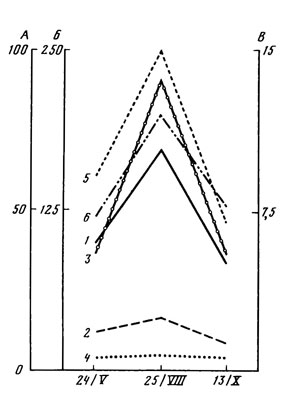
Рис. 7. Сезонные изменения количества водорослей в подстилке и почве сосняка чернично-зеленомощного. Обозначения те же, что на рис. 5
В зависимости от сезона меняется роль отдельных групп водорослей. Летом наиболее массовыми из Chlorophyta являются виды Chlorella, Chlorococcum, Oocystis, Macrochloris и Coccomyxa solorinae. Им сопутствуют обильные нитчатки из Ulotrichales (в подстилке - виды Stichococcus и Chlorhormidium, а в почве - Gongrosira terricola), причем в более влажный год они развиваются интенсивнее, чем в более сухой. Во влажные периоды и обязательно к осени массовыми становятся влаголюбивые хламидомонады. В сезоны с повышенной влажностью в состав доминирующего комплекса входят желтозеленые водоросли. Кроме Botrydiopsis arhiza, которая может быть обильной в любые периоды, весной встречается довольно много видов Characiopsis, а осенью - Monallantus. Летом и особенно осенью усиливается развитие и увеличивается разнообразие синезеленых водорослей, а также отмечается тенденция к некоторой активизации диатомей. Из отдела Cyanophyta весной встречаются как ностоковые (виды Nostoc, Cylindrospermum), так и осциллаториевые (некоторые виды Phormidium), а осенью - преимущественно Nostoc muscorum, N. punctiforme, Cylindrospermum licheniforme. Таким образом, возрастание разнообразия водорослей летом и особенно осенью может происходить за счет развития некоторых теплолюбивых и влаголюбивых видов. Год с более высокими температурами оказывается особенно благоприятным для развития видов Xanthophyta и Cyanophyta.
В песчаной почве под сосняком чернично-зеленомошным наибольшее число видов наблюдается в периоды с самой высокой влажностью: осенью и после обильных дождей летом (см. табл. 12). В слое неразложившейся подстилки (А0) к осени увеличивается обилие зеленых водорослей, одноклеточных (Coccomyxa solorinae) и нитчатых (Chlorhormidium flaccidum var. nitens).
В почве из доминирующих во все сезоны видов Chlorophyta после дождей особенно обильными становятся хламидомонады. Летом значительно представлен Protococcus viridis, осенью он сменяется Radiosphaera sphaerica. Во влажные сезоны доминирующий комплекс обогащается желто-зелеными водорослями - видами Characiopsis и Botrydiopsis arhiza. К осени несколько усиливается развитие сине-зеленой водоросли Phormidium foveolarum.
Колебания численности водорослей в подстилке тоже довольно четко следуют за изменениями влажности (рис. 7). Так, увеличение влажности подстилки летом в 2 - 2,5 раза по сравнению с весенним и осенним периодами сопровождается удвоением количества водорослей. В почве зависимость от влажности выявляется менее четко. В дождливые периоды некоторое накопление водорослей наблюдается на глубине 5 - 10 или 10 - 15 см. В подстилке и почве исследованного сосняка колебания численности водорослей совпадают с кривыми изменений температуры и влажности.
В снытево-осоковой дубраве на темно-серой лесной почве наиболее богатая флора водорослей обнаружена весной, наименее обильная - летом; осеннее увеличение числа видов не достигает весеннего уровня (см. табл. 12). Сезонные изменения количества водорослей в подстилке и почве неодинаковы, но в том и другом случае довольно четко повторяют колебания влажности (рис. 8). В подстилке отмечены наиболее резкие скачки как влажности, так и численности водорослей, при этом наименьшее количество водорослей обнаружено летом, а наибольшее - осенью. В почве колебания влажности происходят более равномерно, соответственно более плавно изменяется и количество водорослей, постепенно увеличиваясь от весны к осени.
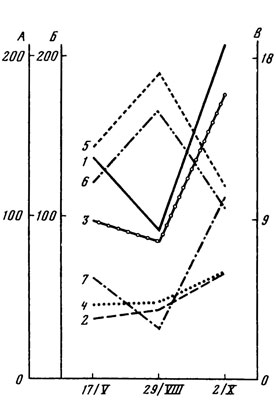
Рис. 8. Сезонные изменения количества водорослей в подстилке и почве снытево-осоковой дубравы. Обозначения 1 - 6 те же, что на рис. 5; 7 - число клеток сине-зеленых водорослей в подстилке
В подстилке преобладающими во все сезоны являются Coccomyxa dispar, Chlorella vulgaris, Ch. terricola. Весной эти виды дополняются зелеными (виды Ulothrix и Chlorhormidium) и желто-зелеными нитчатками, а также обильно развивающимися Nostoc punctiforme. Осенью, кроме одноклеточных зеленых, в состав доминирующего комплекса входят представители отделов Xanthophyta, Cyanophyta, Bacillariophyta. Спектр преобладающих видов Cyanophyta расширяется за счет развития Nostoc paludosum, N. muscorum, N. commune.
В почве доминирующими из зеленых водорослей, кроме видов Chlorella, являются: весной и летом - Stichococcus minor и Chlorosarcina stigmatica, осенью - Chlorococcum humicola. После сильных дождей, прошедших осенью, в почве на глубине 5 - 15 см была обнаружена значительная вегетация хламидомонад, представленных, правда, всего несколькими видами, особенно обильными из которых были Chlamydomonas gloeogama и Ch. elliptica (единственный случай, когда виды Chlamydomonas встретились в большом количестве в темно-серой лесной почве). Из желто-зеленых водорослей во все сезоны доминирует Botrydiopsis arhiza. Весной, кроме нее, обильно растет Gloeobotrys ellipsoideus, а летом и осенью - одноклеточные из Heterococcales, при этом летом в основном преобладают виды Ellipsoidion и Monallantus, а осенью - виды из рода Pleurochloris. Максимальное развитие желто-зеленых водорослей приурочено к периодам с наибольшей влажностью. Представители отдела Cyanophyta входят в состав доминирующего комплекса водорослей только осенью и лишь в самом верхнем слое почвы.
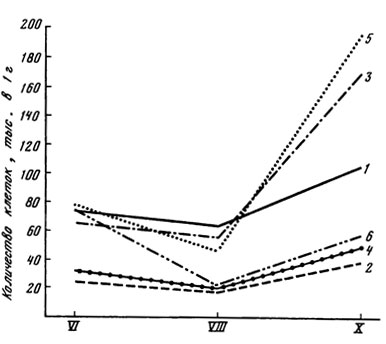
Рис. 9. Динамика численности водорослей в подстилке и почве коренного и производных лесов. 1 - ельник (подстилка); 2 - ельник (почва); 3 - березняк (подстилка); 4 - березняк (почва); 5 - липняк (подстилка): 6 - липняк (почва)
Обнаруженные в исследованных местообитаниях и описанные выше сезонные изменения в развитии водорослей не являются строго характерными вообще для данного типа леса или для данной почвы. Можно говорить лишь о состоянии и степени развития водорослей в исследуемый момент, так как между изученными сроками могут происходить резкие колебания численности водорослей в течение одного месяца и даже в течение одних суток, как это упомянуто выше. Однако же одновременное изучение состава и количества водорослей, проведенное в разных типах леса в близкие сроки, позволило сравнить сезонную динамику в различных почвенно-климатических условиях, обнаружить некоторые тенденции в развитии почвенной альгофлоры, а также проследить влияние на нее ряда экологических факторов.
Наиболее отчетливая корреляция проявляется между развитием почвенных водорослей и сезонными факторами - влажностью и температурой подстилки и почвы. Особенно четкую зависимость от режима влажности показывает численность водорослей [Домрачева, 1974; Новичкова-Иванова, 1969, 1974; Рийс, 1974; Штина, 1974, 1978]. О. Я. Чаплыгина нашла, что коэффициент корреляции численности клеток и влажности почвы лежал в пределах от 0,76 до 0,98, т. е. между этими двумя показателями существовала почти прямолинейная зависимость.
Наиболее существенные колебания в развитии водорослей происходят в подстилке, что обязано резким изменениям ее влажности и температуры. Характер сезонных сукцессий водорослей в подстилке и почве часто различен. Наиболее тесная зависимость динамики численности водорослей от влажности обнаруживается в сосняке чернично-зеленомошном (см. рис. 7) и особенно в снытево-осоковой дубраве (см. рис. 8), т. е. там, где влажность наименьшая и лимитирует развитие водорослей.
Летнее снижение численности клеток, связанное, очевидно, с высыханием почвы, отмечено и в производных типах леса (рис. 9), причем в подстилке оно было более заметным, чем в почве. Видно, что в ельнике некоторое снижение числа клеток происходило только в подстилке и не отмечалось в почве. Вместе с тем во всех типах леса кривая численности резко поднялась в октябре - опять же более заметно в подстилке, особенно в березняке и липняке. К периоду высокой влажности - октябрю - часто приурочено и максимальное число видов водорослей (см. табл. 12, 13).
Изменения количества клеток и числа видов не всегда совпадают, различаясь прежде всего по времени наступления максимумов. Подобное несоответствие было обнаружено и другими авторами [Lund, 1945; Штина, 1959а; Куликова, 1965; Носкова, 1968в]. Было высказано предположение [Штина, 1959а], что видовой состав водорослей является более постоянным признаком альгосинузий. Количество видов водорослей в большинстве случаев четкой корреляции с влажностью не обнаруживает. На видовой состав водорослей влажность оказывает скорее всего косвенное влияние, проявляющееся через жизнедеятельность высших растений и свойства почвы.
Температурный режим почвы воздействует на развитие почвенной альго-флоры, хотя некоторые исследователи не обнаружили корреляции численности и биомассы водорослей с температурой [Раманаускене, 1970; Домрачева, 1972; Рийс, 1974].
Температурный режим почвы в ряде случаев выступает как ведущий фактор в регулировании развития водорослей. Такая зависимость, когда не прослеживается прямой связи с влажностью, а колебания температуры сопровождаются соответствующими изменениями численности водорослей, наблюдается в подстилке ельника-черничника. В других случаях, например, в сосняке чернично-зеленомошном, где изменения температуры и влажности совпадают с колебаниями интенсивности развития водорослей, температура является одним из прямо действующих факторов, усиливающим влияние влажности. В районах с преобладанием достаточно высоких температур непосредственной связи последних с динамикой почвенных водорослей не прослеживается, и в данном случае, очевидно, можно говорить лишь о косвенном влиянии температурного фактора осуществляемом различными путями [Голлербах, Штина, 1969; Рыыс, Рийс, 1972; Рийс, 1974; Штина, 1974,1978].
Ф.Гиндак [Hindak, 1977] нашел в дубово-грабовом лесу максимум биомассы водорослей в летние месяцы, причем пики были особенно резкими в июне; вместе с тем в слое 0 - 1 см отмечался пик, хотя и меньший, чем летом, в декабре. Возможно, что зимний подъем биомассы объясняется методом определения биомассы - по содержанию хлорофилла в почве, и зимний максимум обязан не только водорослям, но и мертвому опаду.
Кроме влажности и температуры почвы, на сезонное развитие водорослей значительное влияние оказывают освещенность под пологом леса, чаще всего связанная с разреженностью растительного покрова, наличие свежего опада, его состав и другие причины.
Эколого-биологические особенности отдельных видов водорослей обусловливают их различное отношение к перечисленным выше факторам, что может привести к определенной периодичности в развитии разных видов водорослей.
Одни виды водорослей, обладая, очевидно, широкой амплитудой приспособляемости, могут достигать максимального развития при самых различных погодных условиях и в разные сезоны. Таковы, например, виды Сосcomyxa, Chlorella, Chlorhormidium и Botrydiopsis. Другие виды водорослей обнаруживают приуроченность к определенным сезонам. Так, только весной были встречены Dictyococcus gernekii, Disporopsis pyrenoidifera, Cylindrocystis crassa, лишь летом - Chloroplana terricola, а осенью - Leptosira polychloris. Летом ни разу не была обнаружена Scotiella levicostata.
Таблица 13
| Тип леса | Отдел водорослей | 1978 г. | 1979 г. | |||
| 12.06 | 20.07 | 12.06 | 16.08 | 16.10 | ||
| Ельник | Сине-зеленые | 2 | 0 | 1 | 0 | 3 |
| Зеленые | 31 | 25 | 20 | 17 | 29 | |
| Желто-зеленые | 9 | 13 | 10 | 5 | 11 | |
| Диатомовые | 1 | 0 | 1 | 0 | 2 | |
| Общее число | 43 | 38 | 32 | 22 | 45 | |
| Березняк | Сине-зеленые | 4 | 2 | 2 | 1 | 7 |
| Зеленые | 33 | 23 | 19 | 11 | 22 | |
| Желто-зеленые | 12 | 16 | 13 | 8 | 17 | |
| Диатомовые | 2 | 0 | 1 | 0 | 4 | |
| Общее число | 51 | 41 | 35 | 20 | 50 | |
| Липняк | Сине-зеленые | 6 | 2 | 3 | 2 | 10 |
| Зеленые | 30 | 28 | 21 | 9 | 23 | |
| Желто-зеленые | 10 | 14 | 12 | 7 | 14 | |
| Диатомовые | 3 | 1 | 1 | 0 | 6 | |
| Общее число | 49 | 45 | 37 | 18 | 53 | |
Желто-зеленые водоросли в основном предпочитают теплое, но влажное время. В частности, только в периоды с самой высокой влажностью отмечено массовое развитие видов Characiops is. Менее требовательные к влаге хлорококковые водоросли [Голлербах, Штина, 1969] в более влажные сезоны уступают ведущую роль влаголюбивым хламидомонадам. Наоборот, в очень сухие периоды из состава альгофлоры выпадают многие хламидомонады и желтозеленые водоросли.
К осени, особенно в лиственных лесах, усиливается развитие и увеличивается разнообразие сине-зеленых и диатомовых водорослей. Это, возможно, связано с кальцефильностью видов Cyanophyta, благодаря чему они должны в первую очередь реагировать на поступление осенью с лиственным опадом значительных количеств биогенных элементов, в том числе кальция. Одной из причин активизации в осеннее время развития сине-зеленых и диатомовых водорослей, по-видимому, является возрастающая в этот период освещенность. Осеннее увеличение развития названных отделов водорослей отмечалось и для других местообитаний [Lund, 1945; Кондратьева, 1958; Rebel, 1959; Штина, 1959а]. Распределение водорослей по сезонам происходит по-разному даже у представителей одного и того же отдела. Так, из сине-зеленых водорослей в осенний период в основном активизируется рост видов из порядка Nostocales. Это связано, вероятно, не только со значительной влажностью в это время и прогреванием почвы за летний период, но и с изменением условий питания. Развитие встречающихся в исследованных почвах видов Phormidium приурочено чаще всего к весне.
Наибольшее развитие диатомовых водорослей приурочено к периодам значительной влажности подстилок (см. рис. 9, табл. 13), даже несмотря на то, что в осенний период отмечались существенные понижения температуры вплоть до заморозков. В теплые же сезоны года, но при очень сильном подсыхании подстилки, особенно ее верхнего слоя, развитие диатомовых водорослей или значительно сокращалось, или прекращалось совсем (см. табл. 12, 13). Обитание диатомей приурочено к самым поверхностным слоям почвы; в наших исследованиях они обнаруживались только в подстилках. Обладая значительной холодостойкостью, светолюбием и солевыносливостью, диатомовые водоросли очень неустойчивы против высыхания. Многие из мелкоклеточных видов диатомовых считаются даже индикаторами почвенной влажности [France, 1913]
Характерно, что в большинстве случаев осенью происходит расширение состава видов доминирующего комплекса водорослей. Т. С. Носкова [1968а] и О.Я.Чаплыгина [1977] подтвердили смену отдельных групп водорослей в связи с их разным отношением к температуре и влажности почвы. Синезеленые из порядка Chroococcales приурочены к влажному и теплому периоду, Nostocales встречаются в течение всего вегетационного периода, но особенно разнообразны летом и осенью, а представители порядка Oscillatoriales приурочены в основном к весеннему и осеннему периодам. Зеленые из порядка Chlorococcales более или менее равномерно распределены в течение вегетационного периода, а видовое разнообразие Chlamydomonas возрастает от весны к лету.
Таким образом, экологические условия, вызывающие изменчивость в развитии почвенных водорослей в течение вегетационного периода, оказывают на них либо непосредственное, либо косвенное влияние, причем к прямо действующим факторам относятся в основном те, которые являются лимитирующими в данном местообитании.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'