Глава III. Основные природные факторы, воздействующие на формирование альгосинузий в лесных почвах
Влияние высших растений
Значительную роль в регулировании жизнедеятельности почвенных организмов играет высшее растение, являясь мощным средообразующим и, в частности, почвообразующим фактором. Оно оказывает существенное влияние на воздушный, гидротермический, световой режимы под пологом леса и соответственно почвы. Высшие растения формируют физико-химические свойства почвы через опад и корневые системы, причем именно в зоне корней осуществляется непосредственный контакт между растениями и водорослями. Таким образом, преобразовывая среду обитания разнообразных почвенных организмов, в том числе водорослей, высшее растение оказывает на них значительное как косвенное, так и прямое воздействие.
В главе III рассматриваются различные аспекты влияния высших растений на водоросли. Сравнение альгосинузий различных лесов, растущих на одинаковой почве, дает возможность выяснить интегральное влияние растений-эдификаторов, которое проявляется через опад и его действие на свойства почвы, через световой и водный режим, через корневую систему.
Для изучения различных сторон влияния высших растений на почвенные водоросли были проведены исследования на уровне биогеоценозов, парцелл, а также ризосферные исследования.
Особенности почвенной альгофлоры различных лесных фитоценозов, развивающихся на однотипных почвах
Для выяснения роли высших растений в формировании водорослевых группировок леса важное значение имеет исходная однородность почвенного покрова.
Примером условий, где влияние высшего растения на развитие почвенных водорослей проявляется достаточно четко, следует считать разные типы древесных насаждений, произрастающие в одной географической зоне, на одном и том же типе почв.
Группировки водорослей в разных типах леса на дерново-подзолистой почве
Одно из исследований с подбором подобного рода условий было выполнено в Серебряноборском и Подушкинском лесничествах Московской области. В первом лесничестве объектами изучения были сосняк сложный дубняково-лещиновый и липняк волосистоосоковый; во втором - ельник-кисличник. Все типы леса расположены на дерново-подзолистой почве.
Подробные ботанические, лесоводственные, почвенные исследования в этих типах леса были проведены С.А.Никитиным [1961], И.Н.Васильевой [1964], С. В. Зонном и Е.А.Кузьминой [1964], И. И. Судницыным [1964], Л. О. Карпачевским и Н.К.Киселевой [1965], А.Е.Максимовой [1969,1974], Л. П. Рысиным и др. [1974]. Кроме того, здесь были выполнены работы по физиологии, микробиологии, зоологии беспозвоночных и ряду других дисциплин. Таким образом, изучение альгофлоры проводилось в рамках общих стационарных исследований.
Во всех рассматриваемых типах леса выявлены характерные особенности альгосинузий, хотя общее число видов водорослей в них различается не очень резко (табл. 14).
В сосняке сложном не только по числу видов, но и по обилию в культурах преобладают виды из отделов Chlorophyta и Xanthophyta. Из зеленых водорослей обильны представители хламидомонадовых, хлорококковых и улотриксовых; из желто-зеленых наиболее обычны одноклеточные из родов Botrydiopsis, Ellipsoid ion, многочисленные виды Characiopsis (они составляют 35,1% от общего числа желто-зеленых), а также Heterococcus chodatii. В подстилке сосняка, состоящей из хвои и значительной примеси лиственного опада, присутствуют сине-зеленые водоросли, хотя и представленные небольшим числом видов (в основном из рода Phormidium). Сосновые леса обычно характеризуются как бедные водорослями [Ваулина, 1956; Rosa, 1962; Новичкова-Иванова, 1969]. В нашем примере большое видовое разнообразие и обилие водорослей в сосняке сложном объясняются, по всей вероятности, значительным участием в древостое лиственных пород, составляющих второй ярус.
В липняке волосистоосоковом разнообразнее всего представлены одноклеточные зеленые водоросли (см. табл. 14). Желто-зеленые водоросли развиты слабее, чем в сосняке сложном. Самой обильной является Botrydiopsis arhiza. В альгофлоре липняка увеличивается роль сине-зеленых и диатомовых водорослей. Синезеленые представлены видами из порядков Oscillatoriales и Nostocales, из них Nostoc paludosum и Phormidium foveolarum входят в число преобладающих. Количество найденных здесь видов отдела ВасiIlariophyta невелико, однако Navicula mutica дает обильные разраста ния. 26% от общего числа видов водорослей, выявленных в липняке волосистоосоковом, найдено только в этом типе леса.
Таблица 14
| Тип леса | Cyanophyta | Chlorophyta | Xanthophyta | Bacillariophyta | Общее число видов | ||||
| число видов | % | число видов | % | число видов | % | число видов | % | ||
| Сосняк дубняково-лещиновый | 4 | 6,6 | 33 | 54,1 | 23 | 37,7 | 1 | 1,6 | 61 |
| Липняк волосисто-осоковый | 5 | 10,3 | 33 | 56,9 | 17 | 29,3 | 2 | 3,5 | 58 |
| Ельник-кисличник | 2 | 3,8 | 31 | 58,5 | 19 | 35,8 | 1 | 1,9 | 53 |
В ельнике-кисличнике обнаружено меньше всего видов водорослей (см. табл. 14). И здесь наиболее многочисленны и разнообразны зеленые водоросли, среди которых превалируют виды-убиквисты из Chlorococcales и представители порядка Chlamydomonadales, разнообразие которых в ельнике особенно велико, в подстилке обильны виды Chlorhormidium и Соссоmyxа. Не входят в доминирующую группу, но играют значительную роль в создании водорослевой синузии ельника-кисличника Scotiella levicostata, Chforoplana terricola, Stichococcus minor. Из желтозеленых водорослей преобладают виды Pleurochloris, Ellipso id ion и Botryochfor is cumulata. В отличие от рассмотренных выше типов леса, в ельнике сине-зеленые водоросли развиты слабо, а диатомовые встречаются единичными экземплярами.
Как видно, разница видового состава и доминирующих групп водорослей определяется прежде всего характером древесной растительности. По мере увеличения доли лиственных пород в составе древостоя возрастает число и обилие представителей синезеленых водорослей (см. табл. 14).
Группировки водорослей в различных лесных насаждениях на выщелоченном черноземе
Интересным объектом для выяснения влияния древесных растений явились искусственные лесные насаждения, созданные на выщелоченном черноземе в Моховском лесничестве Орловской области. Здесь мы имеем единый исходный почвенный фон, на который воздействуют различные виды деревьев.
Альгофлора изучалась в пяти типах лесных насаждений Моховского лесничества: еловом, сосновом, лиственничном, дубовом и березовом (см. табл. 5, участки 23 - 27). Детальные работы по изучению влияния рассматриваемых насаждений на свойства выщелоченного чернозема были проведены С. В. Зонном и Е. А.Кузьминой [1960], В. Н. Миной [1960], И. М.Розановой [1960], Д. Ф.Соколовым [1963]. Микробиологической характеристике почвы исследуемых насаждений посвящены работы Е. В. Руноваи И. Е. Мишустиной [1960], С. В.Егоровой [1962,1968] и других, распределению беспозвоночных животных - Т. С. Перель [1958].
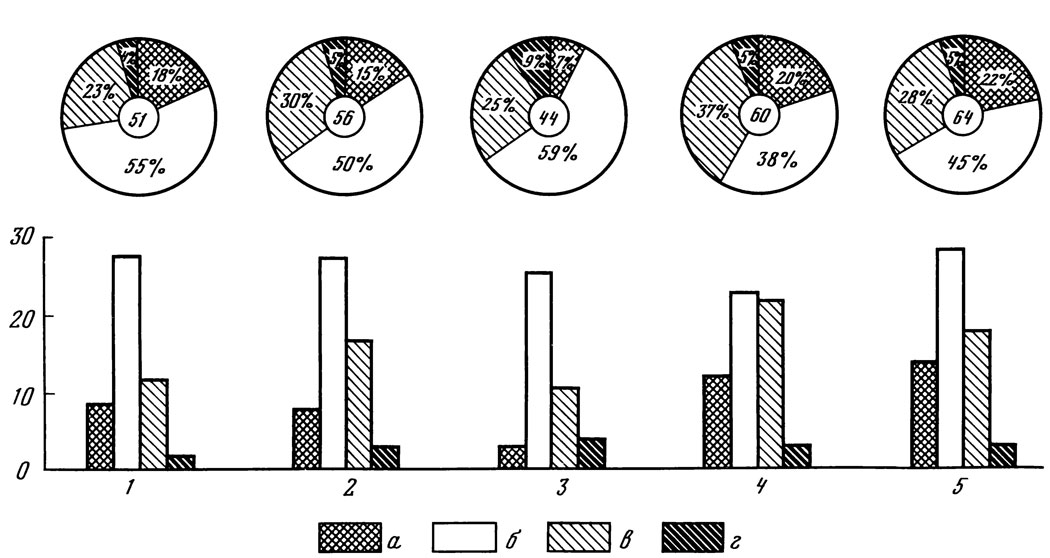
Рис. 10. Соотношение числа видов водорослей разных отделов в лесных насаждениях на выщелоченном черноземе. 1 - еловое; 2 - сосновое; 3 - лиственничное; 4 - дубовое; 5 - березовое насаждения. Во внутреннем круге - общее число видов водорослей, в наружном круге - процентное соотношение разных отделов: а - сине-зеленые; б - зеленые; в - желто-зеленые; г - диатомовые водоросли
Под разными лесными насаждениями в выщелоченном черноземе обнаружены различия в характере альгофлоры, выражающиеся в изменениях числа видов водорослей, соотношении различных систематических групп и степени их развития, а также в составе превалирующих видов водорослей.
Самое большое видовое разнообразие водорослей обнаружено в березовом насаждении, наименьшее - в лиственничном (рис. 10). Интересно, что березовая подстилка и почва под березой оказались наиболее обсемененными бактериями; здесь же обнаружено самое большое количество дождевых червей; в ризосфере березы отмечено максимальное количество микроорганизмов [Перель, 1958; Рунов, Мишустина, 1960]. Указанная повышенная активность многих групп организмов в березовом насаждении может быть связана с наилучшей оструктуренностью почвы, самым высоким pH, наименее выраженной фитонцидностью корней березы и рядом других свойств [Розанова, 1960; Егорова, 1962]. Во всех насаждениях самое большое число видов приходится на зеленые водоросли, второе место по числу видов занимают желто-зеленые водоросли. Синезеленые менее разнообразны, особенно в лиственничном лесу. Диатомовые водоросли во всех изученных лесах представлены единичными видами. Таким образом, распределение водорослей по отделам и в лесах на выщелоченном черноземе остается в основном сходным с наблюдаемым в ранее рассмотренных подзолистых, дерново-подзолистых и темно-серых лесных почвах.
Соотношение систематических групп водорослей в каждом типе леса достаточно специфично, что видно на приводимых спектрах (см. рис. 10); при этом наибольшее разнообразие видов водорослей не обязательно сопровождается обилием особей этого отдела.
В хвойных лесах - еловом, сосновом и лиственничном - на долю видов Chlorophyta приходится больше половины всех обнаруженных видов; зеленые водоросли здесь превалируют и по степени развития. В ельнике разнообразие видов из отделов Xantophyta и Cyanophyta довольно близкое, а в сосняке и лиственничнике число видов первого отдела в 2 - 3,5 раза выше, чем второго.
В еловом лесу в подстилке зеленые водоросли, представленные в основном нитчатыми формами, сопровождаются желто-зелеными и сине-зелеными нитчатками, а в почве из зеленых, кроме видов Ulotrichales, господствуют представители рода Chlamydomonas при субдоминировании желто-зеленых из Heterococcales.
В сосновом лесу выявлено значительное количество видов водорослей. Здесь отмечено более обильное развитие синезеленых и диатомовых, что, возможно, связано с наличием подлеска, состоящего в основном из бузины.
Примесь лиственных пород в хвойных лесах существенно изменяет в них биологический круговорот [Ремезов и др., 1959]. Листья бузины богаты зольными элементами и азотом, поэтому они могут ускорять разложение подстилки в сосняке [Розанова, 1960; Соколов, 1963], что способствует развитию в сосновом насаждении многих видов водорослей, в том числе сине-зеленых и диатомовых.
В лиственничном лесу наряду с общей видовой обедненностью отмечена малочисленность видов синезеленых и диатомовых, которые имеют здесь и самое слабое развитие. В лиственничнике обнаружены сравнительно невысокие запасы питательных элементов, при этом содержание кальция ниже, чем фосфора, кремнезема и калия; довольно низким оказался рН лиственничной подстилки [Розанова, 1960]. В то же время корни лиственницы, обладающие самой высокой фитонцидностью, оказали наибольшее угнетающее действие на все группы микроорганизмов, на рост корешков и величину проростков растений [Егорова, 1962]. Все это вместе взятое, по-видимому, в значительной мере объясняет как обедненность видового состава, так и небольшое разнообразие и развитие указанных выше отделов водорослей в лиственничном насаждении.
В лиственных лесах на долю представителей отдела Chlorophyta приходится не более 38-45% от всех видов водорослей. В дубовом насаждении возрастает процентное содержание желто-зеленых водорослей, почти сравниваясь с содержанием зеленых; в березняке почти одинаково соотношение желто-зеленых и сине-зеленых водорослей (см. рис. 10).
В дубраве превалируют зеленые и сине-зеленые водоросли; последние значительно представлены в подстилке (12 видов) и в верхнем 5-сантиметровом слое почвы (10 видов). Не уступающие зелёным водорослям по разнообразию виды желто-зеленых имеют здесь меньшее обилие в культурах, чем первые. В доминирующий комплекс подтилки входят диатомовые водоросли, хотя видовое разнообразие их незначительно.
В березняке как по сравнению с другими типами насаждений, так и по сравнению с другими отделами водорослей, наиболее обильны сине-зеленые водоросли, представленные видами из Nostocales, и несколько меньше из Oscillatoriales. Богата ими не только подстилка (14 видов), но и весь 10-сантиметровый слой почвы. Из зеленых водорослей больше, чем в дубняке, встречено хламидомонад. Таким образом, видно, что характер доминирующего комплекса водорослей в разных насаждениях варьирует.
Своеобразие каждого из изучаемых насаждений заключается и в видовом составе доминирующих водорослей. Так, преобладающие в подстилках зеленые нитчатки в еловом, сосновом и лиственничном лесах представлены родом Chlorhormidium, к которому в еловом лесу присоединяется еще Stichococcus minor; в дубняке и березняке - родом Ulothrix, дополненным в березовом насаждении обильной Microspora tumidula. Из входящих в доминирующий комплекс колониальных зеленых водорослей под всеми породами присутствует Borodinella polytetras, особенно обильная под березой и сосной, несколько меньше - под лиственницей. Кроме нее, в почве сосняка значительно развиты Palmella, в почве дубняка - Chlorosarcinopsis, а под березняком - Disporopsis. Наиболее же обильны колониальные формы зеленых водорослей в почве березового и соснового лесов. Преобладающие колониальные формы из отдела Xanthophyta представлены: в еловом лесу родом Botryochloris, в березовом - родом Sphaerosorus, в дубовом и сосновом - родом Gloeobotrys. Почвенная альгофлора изученных лесных насаждений содержит определенное количество специфических видов, нередко совпадающих с видами, выявленными для сходных растительных ассоциаций, но произрастающих на другом типе почв.
Как и на дерново-подзолистой почве, здесь обнаружены особенно заметные различия между группировками водорослей хвойных и лиственных лесов. Лиственные леса оказались более насыщенными видами водорослей по сравнению с чистыми хвойными насаждениями. Лиственные леса богаче хвойных синезелеными водорослями. В лиственных лесах обильнее и диатомовые водоросли, однако во всех типах насаждений их видовое разнообразие остается очень небольшим.
Одной из характерных черт альгосинузий хвойных лесов, особенно елового, является разнообразие и активное развитие видов Chlamydomonas, входящих в состав доминирующей группы водорослей. В дубовом и березовом насаждениях хламидомонады не только представлены меньшим числом видов, но и не обнаруживают столь значительного развития. Сходные данные находим у Л. Н. Новичковой-Ивановой [1968], которая указывает на обильное развитие хламидомонад как на основное отличие синузии чистого соснового леса от дубово-лиственнично-соснового. Такие же особенности альгофлоры хвойных лесов описаны нами ранее для более северных районов.
Специфика группировок водорослей в разных лесных насаждениях связана с условиями, складывающимися под пологом различных древесных пород. Благодаря же тому, что изученные насаждения были заложены на одном и том же типе почвы, в идентичных условиях, можно считать, что все эти условия складываются прежде всего в зависимости от вида дерева-эдификатора.
Следует подчеркнуть многообразие влияния высших растений на все экологические особенности лесного биогеоценоза.
Характер растительности влияет на ряд свойств почвы, в частности, на ее активную реакцию, которая в значительной мере определяет степень развития водорослей и тип альгофлоры [Голлербах, 1936; Schelhorn, 1936, Stokes, 1940, John, 1942, Lund, 1947a; 1962; Голлербах, Штина, 1969; По- цене, 1970; Rosa, Lhotsky, 1971]. Низкое значение рН в кислых лесных почвах ограничивает развитие водорослей [Chodat, Chastain, 1957]. Особенно сильно низкая кислотность почв сказывается на развитии наиболее чувствительных к ней видов отделов Cyanophyta и Bacillariophyta, обусловливая или их малочисленность и бедность состава [Flint, 1958, MacEntee, Bold, 1974], или почти полное отсутствие [Schelhorn, 1936, Hofstetter, 1968, Jurgensen, Davey, 1968, MacEntee, 1970]. Брендемюль [Brendemuhl, 1949], сравнивая лиственные и хвойные леса, отметила, что в последних диатомовые водоросли отсутствовали в 80% случаев, в то время как в первых - лишь в 43% случаев, и считала одной из причин бедности флоры ди- атомей в хвойных лесах кислую реакцию почвы.
В разных типах леса складывается различный режим влажности. С влажными местообитаниями и типами леса связано наибольшее развитие водорослей [Rosa, 1962; Носкова, 1968а] и, в частности, отдельных влаголюбивых групп, представители которых рассматриваются как индикаторы почвенной влажности [Бусыгина, 1974, 1976], например, диатомей [France, 1913, Brendemuhl, 1949], сине-зеленых из порядка Chroococcales [Носкова, 1968б], некоторых видов зеленых водорослей [Мейер, 1937; Голлербах, Штина, 1969; Новичкова-Иванова, 1969].
Высшими растениями определяется световой режим в лесу, влияющий на водорослевые группировки. Благоприятным периодом для развития водорослей является время, когда листья на деревьях отсутствуют [Petersen, 1935, Lund, 1945]. Недостаток света, связанный с густым растительным покровом или наличием мощной лесной подстилки, обусловливает снижение развития водорослей в лесных почвах [Brendemuhl, 1949, Hofstetter, 1968, Jurgensen, Davey, 1968; Новичкова-Иванова, 1969]. Косвенное влияние высших растений на водоросли может осуществляться также через изменения температурного и пищевого режима среды обитания водорослей.
Так как образованием, трансформирующим влияние высших растений на живое население почвы, является подстилка, то специфические особенности альгофлоры в разных типах леса могут быть обусловлены различиями в количестве и свойствах опада и подстилки, зависящими от породного состава деревьев. Если учесть к тому же, что основное распространение водорослей в лесах ограничено подстилкой и самыми верхними слоями почвы (глава II), то можно предполагать, что различия в альгофлоре разных типов леса прежде всего будут проявляться в особенностях альгофлоры подстилок. Поэтому целесообразно более подробно рассмотреть роль лесной подстилки в формировании альгосинузий лесов.
Влияние лесной подстилки
Особая роль лесной подстилки в жизни лесных биогеоценозов подчеркивается как геоботаниками [Сукачев, 1961; Работнов, 1978; и др.], так и почвоведами [Зонн, 1964; и др.]. Через подстилку и населяющие ее организмы проходит один из главных путей воздействия растений на почвообразовательный процесс. Рассматривая подстилку как специфическое лесное органогенное образование, обращают внимание на следующие его особенности: подстилка является результатом жизнедеятельности организмов, она регулирует воздействие последних на почву и вынос подвижных соединений; наконец - подстилка является горизонтом наиболее активной биогео-ценотической деятельности. Все микробиологические процессы протекают в подстилке более интенсивно, чем в почвах. Микробное население подстилок своеобразно и характеризуется иными качественными и количественными показателями, чем микрофлора почвы: на 1 г сухого вещества подстилок приходится в десятки и сотни раз больше микроорганизмов, чем на соответствующую навеску почвы [Егорова и др., 1964]. Правда, надо иметь в виду, что объемные массы подстилки и почвы тоже различаются на порядок. Растительному опаду и лесной подстилке принадлежит важная роль и в формировании водорослевых группировок [Штина, Ройзин, 1966; Носкова, 1968а; Алексахина, 1971 - 1982; Чаплыгина, 1977; и др.]. В подстилке создаются некоторые условия, благоприятные для развития водорослей: освещенность, накопление влаги, происходящее даже за счет конденсационной воды. Выше (глава II) было показано, что численность водорослей, как и гетеротрофных микроорганизмов, в подстилке выше, чем в почве (см. рис. 2, 5 - 8; табл. 9, 11). Вместе с тем подстилка обладает рядом свойств, ограничивающих жизнедеятельность некоторые видов водорослей и обусловливающих отбор специфических видов, характерных для подстилок. Такими неблагоприятными для водорослей особенностями подстилки являются: обилие органического вещества, которое интенсивно разлагается массой гетеротрофных организмов - антагонистов водорослей и наличие ряда токсических соединений, содержащихся в опаде или образуемых микроорганизмами. При изучении гетеротрофной микрофлоры установлено [Рунов, Мишустина, 1960; Рунов, Егорова, 1962; Егорова и др., 1964; и др.], что в зависимости от биохимического состава подстилок и почвенно-климатических условий подстилки заселяются специфическими видами и разложение идет по определенному типу. Наибольшие различия в микрофлоре подстилок наблюдались между хвойными и лиственными породами [Егорова и др., 1964; Егорова, Лаврова, 1982; и др.].
Подобные различия упоминались выше и в отношении водорослей (см. рис. 3, глава II). Во всех изученных лесах обнаружены различия в составе и числе видов водорослей между подстилкой и почвенными горизонтами. Отмечена видовая обедненность подстилок по сравнению с почвой, что особенно заметно выражено в хвойных лесах. Этот факт, с одной стороны, объясняется меньшим разнообразием хламидомонад и желто-зеленых водорослей в подстилке, а, с другой стороны, обусловливается небольшим разнообразием видов Cyanophyta и Bacillariophyta, т. е. тех отделов водорослей, основное распространение которых, как это было показано выше, приурочено к подстилке.
При сравнении количества видов и численности водорослей в подстилках восьми различных типов леса (табл. 15) подтвердилась полученная закономерность: с увеличением доли листового опада возрастает роль синезеленых и диатомовых водорослей.
Следует заметить что в таблице приведены данные только по весеннему сроку (это необходимо было для сопоставления результатов). Осенью, когда водоросли описываемых отделов развиваются в лесных почвах наиболее интенсивно, разница между хвойными и лиственными подстилками проступает еще резче.
Таблица 15
| Тип леса | Сине-зеленые | Диатомовые | ||
| % от числа видов | % от количества клеток | % от числа видов | % от количества клеток | |
| Ельник-черничник | 7,1 | <4,0 | 0 | 0 |
| Ельник-кисличник | 8,0 | - | 4,0 | 0 |
| Дубо-ельник волосистоосоковый | 17,7 | 3,6 | 5,9 | 3,1 |
| Липняк волосистоосоковый | 20,7 | - | 6,9 | - |
| Дубо-липняк волосистоосоковый | 22,2 | 9,2 | 7,4 | 3,6 |
| Дубрава снытево-осоковая | 31,6 | 46,3 | 6,3 | 2,8 |
| Дубрава пойменная | 65,3 | 75,0 | 10,2 | 6,8 |
| Дубрава солонцовая | 27,0 | 38,1 | 13,5 | 22,4 |
Примечание. Прочерк означает отсутствие количественного учета.
Значительное развитие видов Cyanophyta в лиственных лесах отмечено и для других местообитаний [Матвиенко, 1950; Штина, 1959а; Gruia, 1965; Носкова, 1968а; Новичкова-Иванова, 1969; MacEntee, 1970; и др.].
В качестве условий, лимитирующих распространение сине-зеленых водорослей (в частности, азотфиксирующих) в хвойных лесах могут выступать факторы, связанные с косвенным влиянием высшего растения [Jurgensen, Davey, 1968], например, кислотность, низкая интенсивность света, менее благоприятные условия влажности и температуры. Действительно, под изученными лиственными лесами обнаружились более глубокие промачиваемость почвы и проникновение корней, большие величины доступной для растений влаги и полевой влагоемкости, более высокая температура почвы, меньшая кислотность подстилки и почвы и более высокое содержание многих зольных элементов, чем под хвойными лесами [Васильева, 1964; Зонн, Кузьмина, 1964; Судницын, 1964].
Как было установлено, лиственные леса вовлекают в биологический круговорот значительно больше зольных элементов питания, в том числе кальция, калия, азота и фосфора, чем хвойные [Ремезов и др., 1959]. В лиственных лесах шире, чем в хвойных, соотношение N : С, т. е. лиственные подстилки относительно обогащены азотистыми веществами [Розанова, 1960].
В изученных лесах подстилка липняка характеризуется минимальной кислотностью (pH горизонта А0 в липняке 6,2, а в ельнике - 5,5), наибольшим содержанием азота и кремния и достаточным количеством кальция [Максимова, 1974], значительной интенсивностью процессов минерализации, высокой активностью разложения клетчатки и интенсивной нитрификационной способностью. [Терехов, Еникеева, 1964; Большакова, 1974].
Благодаря значительной примеси листового опада (за счет присутствия лещины) заметно улучшаются свойства хвойной подстилки в сосняке. Листовой опад, богатый зольными элементами, нейтрализует органические кислоты, увеличивая pH подстилки [Судницын, 1964]. В сосняке, сложном по сравнению с чистыми хвойными насаждениями, интенсивнее проходит минерализация подстилки и опада, в почве содержится больше растворимых органических веществ, отмечается более высокая концентрация подвижных соединений кальция, фосфора и калия [Максимова, 1969, 1974]. С увеличением доли листового опада улучшаются и некоторые физические свойства хвойной подстилки, в частности, возрастает влагоемкость и уменьшается объемный вес подстилки [Теребуха, 1969].
Таким образом, в смешанных и особенно лиственных лесах складываются условия, более благоприятные для развития сине-зеленых и диатомовых водорослей, отличающихся высокой потребностью в кальции, азоте и кремнии и приуроченностью к менее кислым условиям среды и к субстратам, где интенсивнее протекают процессы минерализации [Schelhorn, 1936, Lund, 1945, 1947 а, в, 1962; Brendemuhl, 1949, Hofstetter, 1968, Holm-Hansen, 1968; Кордэ, 1960; Голлербах, Штина, 1969]. Следует присоединиться к мнению К. Розы [Rosa, 1S56, 1962], который большое значение в развитии сине-зеленых водорослей придает листовому опаду.
Вместе с тем в подстилках лиственных лесов оказалась выше ферментативная активность, вследствие чего они содержат легкодоступные, а подстилки хвойных - более закрепленные органические вещества [Рунов, Мишустина, 1960]. В подстилках и почве лиственных насаждений, а также в ризосфере лиственных пород энергичнее протекают процессы нитрификации. В почвах под изученными лиственными лесами обнаружены больший обменный вес, pH и влажность, запас гумуса и количество углеродсодержащих веществ, более интенсивное выделение CO2 [Рунов, Соколов, 1958; Мина, 1960; Соколов, 1963]. Однако подстилки в большинстве случаев содержат меньшее число водорослей, чем почва (см. рис. 3), т. е. некоторые виды выпадают из альгофлоры подстилок.
Определенная селекционирующая роль опада и подстилки связана, очевидно, с наличием в ней летучих и воднорастворимых физиологически активных веществ и фитонцидными свойствами подстилки [Рунов, Еникеева, 1955]. Так, по отношению к гетеротрофным микроорганизмам было обнаружено, что подстилки хвойных пород обладают более сильным угнетающим действием, чем подстилки лиственных [Егорова, 1968; Гришина, 1974]. Сильное токсичное влияние на микроорганизмы оказали экстракты из опада дуба и клена [Рунов, Егорова, 1962]. Авторы объясняют фитонцидность подстилок наличием фенольных группировок в дубильных веществах и накоплением продуктов жизнедеятельности микроорганизмов.
Т. А. Овчинникова [1980] показала токсичное влияние вытяжек из листового опада дуба на развитие хлореллы, выделенной из почвы Красносамарского лесничества. Действие аллелопатических активных веществе зависело от концентрации и срока опыта: со временем происходит адаптация хлореллы, что, возможно, связано с развитием бактерий.
Развитию некоторых видов водорослей в подстилке могут препятствовать грибы, в изобилии ее населяющие. Интенсивное развитие грибов в колбах со средой, инокулированной подстилкой, обнаружено в наших опытах. В обильно развивающихся грибных пленках преобладали представители родов PeniciIlium, Mucor, Trichoderma, Fusarium. В подстилках ельников на среднеподзолистых почвах длина мицелия составила от 581 до 1133 м на 1 г субстрата [Мирчинк, Степанова, 1982]. Т. Г. Мирчинк [1976] отмечает, что грибам принадлежит главная роль в разложении хвойного опада. Должно быть, именно массовое развитие грибов ингибирует водоросли в подстилке хвойных лесов. Существование антагонизма между некоторыми группами водорослей и грибов отмечалось не раз [Портнова-Перминова, 1956; Parker, Bold, 1961, Cameron, 1962; Бажина, Штина, 1967; Третьякова, 1981]. Определенную роль играют и почвенные беспозвоночные, часто наблюдаемые нами при просмотре подстилочного материала и обладающие пищевой избирательностью [Штина и др., 1974 - 1981; Nekrasova et aI., 1976; и др.].
Разнообразие видов в подстилке хвойных лесов уменьшается преимущественно за счет видов хламидомонад и желто-зеленых водорослей [Алексахина, 1971; Чаплыгина, 1976]. Характерным подстилочным родом является, как указано выше, Coccomyxa. С другой стороны, для всех лесов, а особенно для лиственных, характерно проникновение в заметных количествах сине-зеленых и диатомовых водорослей из обычного места их массового размножения - подстилки - в верхний горизонт почвы.
Влияние мозаичного строения растительного покрова
Зависимость почвенной альгофлоры от парцеллярного строения биогеоценоза
Согласно современным представлениям, лесной биогеоценоз есть сложное структурное образование, комплексные части которого по предложению Н. В. Дылиса [Дылис и др., 1964; Дылис, 1969] получили название "биогеоценотических парцелл". Специфика биогеоценотического обмена парцелл прежде всего определяется высшими растениями, поэтому парцеллярные исследования могут стать важным звеном в изучении влияния отдельных эдификаторов леса на характер альгофлоры. Дифференциация парцелл наиболее ярко выражена в смешанных биогеоценозах, поэтому для исследования был выбран широколиственно-еловый лес - дубо-ельник волосистоосоковый в Малинском лесничестве Московской области. Изучены следующие парцеллы: основная - елово-волосистоосоковая, занимающая больше половины общей площади изученного участка, и дополняющие: дубово-снытевая, осиново-снытевая и крупнопапоротниковая в окне.
Разнообразие в растительном покрове вызывает значительные различия между парцеллами в условиях увлажнения и освещенности, в физико-химических свойствах подстилки и почвы, в заселенности микроорганизмами, беспозвоночными и позвоночными животными и в других свойствах [Абатуров, Карпачевский, 1965, Карпачевский и др., 1968; Дылис, 1969; Медведев, 1971; Карпачевский, 1977; Алейникова и др., 1979]. Для изучения влияния парцеллярных различий на характер водорослевых группировок проводили изучение видового состава водорослей в каждой парцелле в пятикратной повторности. Максимальное количество выявленных видов водорослей представлено в табл. 16.
Определено и количество видов, специфичных для той или иной парцеллы: в елово-волосистоосоковой их оказалось 17, в осиново-снытевой - 22, в крупнопапоротниковой - 11 видов [Алексахина, 1971].
Альгофлора разных парцелл различается не только насыщенностью видами, но и премущественным развитием различных групп водорослей. Наиболее богатая водорослевая группировка отмечена в осиново-снытевой парцелле. Она характеризуется преобладанием желто-зеленых одноклеточных водорослей, на долю которых приходится 48,7% от общего числа обнаруженных здесь видов. Наряду с обилием различных Characiopsis в этой парцелле разнообразно представлены роды Pleurochlor is, Monodus.
Для водорослевого сообщества елово-волосистоосоковой парцеллы характерна ведущая роль зеленых водорослей со значительным развитием хлорококковых и хламидомонад. Наименее насыщенной видами оказалась крупнопапоротниковая парцелла, т. е. парцелла, сформированная травянистым растением-эдификатором.
Таблица 16
| Отдел водорослей | Парцелла и число видов | ||||
| елово-волосистоосоковая | осиново-снытевая | дубово-снытевая | крупнопапоротниковая | ||
| Всего | Общие с другими парцеллами | ||||
| Cyanophyta | 5 | 4 | 8 | 9 | 9 |
| Chlorophyta | 40 | 32 | 30 | 36 | 23 |
| Xanthophyta | 22 | 20 | 38 | 24 | 23 |
| Bacillariophyta | 1 | 1 | 2 | 2 | 3 |
| Общее число видов | 68 | 57 | 78 | 71 | 58 |
В осиново-снытевой, дубово-снытевой и крупнопапоротниковой парцеллах повышается значение синезеленых водорослей, среди которых особенно массовыми являются виды Nostoc - N. punctiforme, N. muscorum и N. paludosum. В крупнопапоротниковой парцелле зафиксировано, кроме того, более активное развитие в культурах диатомей, хотя видовой состав их и здесь беден.
Обилие сине-зеленых и диатомовых водорослей, очевидно, связано с лучшими условиями освещения и увлажнения, складывающимися в осиново-снытевой и крупнопапоротниковой парцеллах. Так, освещенность в еловых парцеллах оказалась приблизительно в 3 раза меньше, чем в лиственных и в окнах [Дылис, 1969]. Поступление летних осадков в парцеллах, образуемых дубом или осиной, на 80% выше, чем в еловых, а в связи с этим почва в последних к осени высыхает значительно больше, чем в парцеллах, сформированных лиственными породами [Карпачевский и др., 1968].
Для характеристики степени различия флористического состава водорослей в изученных парцеллах были определены коэффициент дифференциальности и коэффициент специфичности [Голлербах, 1936; Воронов, 1973].
Коэффициент дифференциальности (Kд) рассчитывался по формуле:
| Kд = | a = b -2c | x 100, |
| a + b - c |
где a, b - число видов водорослей в первой и второй парцеллах; с - число видов, общих для этих двух парцелл (табл. 17).
Высокие значения коэффициента дифференциальности (см. табл. 17) показывают, что отмеченные между парцеллами различия в составе водорослей весьма существенны.
Коэффициент специфичности (Kсп) находили по формуле:
| Kсп = | q | x 100, |
| a |
где q - количество специфических видов водорослей в парцелле; а - общее количество видов водорослей в парцелле.
Таблица 17
| Сравниваемые парцеллы | Сине-зеленые | Зеленые | Желто-зеленые | Диатомовые | Общее число видов |
| Елово-волосистоосоковая и осиново-снытевая | 56 | 54 | 50 | 50 | 53 |
| Елово-волосистоосоковая и дубово-снытевая | 73 | 57 | 61 | 50 | 60 |
| Елово-волосистоосоковая и крупнопапоротниковая | 73 | 57 | 50 | 67 | 57 |
| Дубово-снытевая и осиново-снытевая | 58 | 65 | 52 | 67 | 59 |
| Дубово-снытевая и крупно-папоротниковая | 50 | 66 | 58 | 75 | 61 |
| Осиново-снытевая и крупнопапоротниковая | 58 | 64 | 48 | 33 | 55 |
Значения коэффициентов специфичности приведены в табл. 18. Как видим, до 25% от общего числа обнаруженных видов и еще больше - от числа видов некоторых отделов может приходиться на специфические виды. Это еще раз подтверждает значительное своеобразие водорослевых группировок парцелл. В лиственных парцеллах специфических видов оказалось больше, чем в хвойной и травянистой, что свидетельствует о более сильном влиянии лиственных пород на почвенные водоросли.
Таблица 18
| Парцелла | Сине-зеленые | Зеленые | Желто-зеленые | Диатомовые | Общее число видов |
| Елово-волосистоосоковая | 20 | 20 | 9 | 0 | 16 |
| Дубово-снытевая | 22 | 31 | 17 | 50 | 25 |
| Осиново-снытевая | 25 | 23 | 24 | 0 | 23 |
| Крупнопапоротниковая | 22 | 13 | 9 | 33 | 16 |
Экспериментальные данные были подвергнуты статистической обработке с использованием критерия Стъюдента [Урбах, 1964]. Сравнивалось среднее число видов из пяти повторностей, обнаруженное в осиново-снытевой и в крупнопапоротниковой парцеллах, по отношению к среднему числу видов в елово-волосистоосоковой парцелле как основной в этом типе леса (табл. 19).
Как видно, различия в общем числе видов водорослей между парцеллами оказались статистически достоверными при доверительной вероятности (Р) 95 и 99%. Разница в содержании видов выделенных отделов водорослей между парцеллами также, как правило, остается статистически достоверной (при Р = 95, 99 и 99,9%) за исключением двух случаев.
Для того, чтобы оценить не только различия, но и взаимные связи водорослевого населения различных парцелл, обусловленные одной почвенно-климатической средой, был подсчитан коэффициент общности Жаккара [Грейг-Смит, 1967; Воронов, 1973]. Коэффициент (К0) рассчитывался по формуле (обозначения те же, что и в предыдущих формулах) К0 = = с / (а + Ь +с).
Таблица 19
| Отдел водорослей | Парцелла | ||
| елово-волосистоосоковая | осиново-снытевая | крупнопапоротниковая | |
| Cyanophyta | 4,2 ± 0,4 | 6,0 ± 0,5** | 7,0 ± 0,5*** |
| Chlorophyta | 32,8 ± 1,5 | 24,2 ± 1,5*** | 17,8 ± 1,5*** |
| Xanthophyta | 15,4 ± 0,9 | 32,0 ± 1,4**** | 17,2 ± 1,1* |
| Bacillariophyta | 0,6 ± 0,2 | 1,2 ± 0,4* | 2,6 ± 0,2**** |
| Общее число видов | 53,0 ± 2,0 | 63,4 ± 2,0*** | 44,6 ± 1,7** |
Примечание. Приведены средние значения ± оценка стандартного отклонения среднего значения.
* - различия статистически недостоверны при Р = 95%;
** - различия статистически достоверны при Р = 95%;
*** - различия статистически достоверны при Р = 99%;
**** - различия статистически достоверны при Р = 99,9%.
Значения К по общему числу видов водорослей и по числу видов наиболее распространенных отделов представлены в табл. 20. Сравнение коэффициента общности и коэффициента дифференциальности видового состава водорослей разных парцелл показывает, что последний почти во всех случаях имеет большие значения.
Таблица 20
| Парцелла | Парцелла | |||
| елово-волосистоосоковая | осиново-снытевая | дубово-снытевая | крупнопапоротниковая | |
| По общему числу видов водорослей | ||||
| Елово-волосистоосоковая | 1 | 0,47 | 0,4 | 0,43 |
| Осиново-снытевая | 0,47 | 1 | 0,41 | 0,45 |
| Дубово-снытевая | 0,4 | 0,41 | 1 | 0,39 |
| Крупнопапоротниковая | 0,43 | 0,45 | 0,39 | 1 |
| По видовому составу зеленых водорослей | ||||
| Елово-волосистоосоковая | 1 | 0,46 | 0,43 | 0,43 |
| Осиново-снытевая | 0,46 | 1 | 0,35 | 0,36 |
| Дубово-снытевая | 0,43 | 0,35 | 1 | 0,34 |
| Крупнопапоротниковая | 0,43 | 0,36 | 0,34 | 1 |
| По видовому составу желто-зеленых водорослей | ||||
| Елово-волосистоосоковая | 1 | 0,5 | 0,39 | 0,5 |
| Осиново-снытевая | 0,5 | 1 | 0,48 | 0,53 |
| Дубово-снытевая | 0,39 | 0,48 | 1 | 0,42 |
| Крупнопапоротниковая | 0,5 | 0,53 | 0,42 | 1 |
Степень флористической общности для наглядности можно представить в виде схем, построенных по принципу, предложенному Л. Н. Медведевым [1971]. Схемы (рис. 11) показывают связь каждой парцеллы с тремя другими. Число линий, соединяющих пару парцелл, пропорционально величине К0.
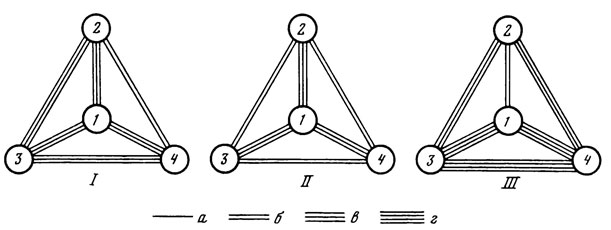
Рис. 11. Схема флористической общности парцелл. I - по общему видовому составу; II - по составу зеленых водорослей; III - по составу желтозеленых водорослей; Парцеллы: 1 - елово-волосистоосоковая; 2 - дубово-снытевая; 3 - осиново-снытевая; 4 - крупнопапоротниковая. Коэффициенты общности (Кsub0/sub) : а = 0,2 - 0,29; б = 0,3 - 0,39; в = 0,4 - 0,49; г = 0,5 - 0,6
Из предложенных схем видно, что елово-волосистоосоковая парцелла имеет самую значительную связь с остальными исследованными парцеллами как по общему количеству обнаруженных видов водорослей, так и по отделам, наиболее разнообразно представленным в данном типе леса. Кроме того, сравнивая состав водорослей, выявленных в разных парцеллах (см. табл. 16), отмечаем, что большинство видов, найденных в основной, типовой, парцелле данного биогеоценоза [Дылис и др., 1964], - елово-волосистоосоковой, являются общими для водорослевых группировок трех других парцелл, образованных эдификаторами-субдоминантами - дубом, осиной и папоротником. Следовательно, в водорослевой группировке ведущей парцеллы отражены закономерности строения альгосинузии исследуемого биогеоценоза. Все это, в свою очередь, позволяет предположить, что для составления общей предварительной характеристики альгофлоры биогеоценоза можно исследовать группировку водорослей в парцелле, занимающей наибольшую площадь - в основной, определяющей и направление всего биогеоценотического обмена в данном типе леса [Дылис, 1969].
Подобное исследование состава водорослей в разных синузиях проведено в ельнике с дубом зеленчуково-кислично-волосистоосоковым, травяной ярус которого обладает значительной пестротой (табл. 21).
Синузии ельника различались по числу видов и составу доминирующих видов. Больше всего видов в основной синузии, где в доминирующий комплекс входят виды из родов Bracteacoccus,Spongiococcum, Chlamydomonas и Botrydiopsis. Отличительной чертой мертвопокровной синузии оказалось многообразие жизненных форм в доминирующем комплексе: кроме одноклеточных и колониальных форм, обильные нитчатки из Ulotrichales. В кисличной синузии преобладали Chlorococcum и одноклеточные желтозеленые.
Полученные данные показали наличие заметной фрагментации водорослевого сообщества в пределах биогеоценоза. Л. П. Приходькова [1977] исследовала сине-зеленые водоросли в трех парцеллах разнотравно-типчаково-ковыльного биогеоценоза Хомутовской степи. Коэффициент флористической общности оказался довольно высоким - 71,4 - 84,6%, 10 видов из 14 были общими для всех парцелл. Однако парцеллы заметно различались по количественному развитию и по встречаемости отдельных видов.
Таблица 21
| Отдел водорослей | Синузия | |||
| зеленчуково-волосистоосоковая | зеленомошная | мертвопокровная | кисличная | |
| Сине-зеленые | 2 | 1 | 1 | 0 |
| Зеленые | 19 | 15 | 17 | 12 |
| Желто-зеленые | 14 | 9 | 12 | 9 |
| Диатомовые | 1 | 1 | 2 | 1 |
| Общее число видов | 36 | 26 | 32 | 21 |
Н. В. Сдобникова 1961], изучавшая синузии водорослей основных растительных ассоциаций степной зоны, обнаружила, что альгологическая карта по сравнению с геоботанической имеет меньшую степень дробности. Можно предположить, что под травяными ассоциациями более важную роль в формировании альгофлоры играют свойства почвы, в лесных же фитоценозах влияние растительности проявляется значительно сильнее. Это предположение подтверждается данными Т. С. Носковой [1968а] и М. Г. Шушуевой [1980], которые нашли, что травянистая растительность в меньшей степени изменяет характер альгосинузий, чем древесная.
Распределение водорослей в пределах проекций крон деревьев-эдификаторов
Одной из причин пространственной гетерогенности лесной почвы и ее населения является специфичность строения древостоя: по мере удаления от ствола дерева изменяются величина опада и запаса подстилки, количество проникающих сквозь крону осадков и света и соответственно свойства почвы [Карпачевский и др., 1968]. Поэтому отбор образцов проводили приблизительно на одинаковом расстоянии от ствола дерева-эдификатора.
О. Я. Чаплыгина [1977] провела специальное сравнение состава и численности водорослей в зависимости от расстояния от ствола дерева. По мере удаления от ствола к периферии кроны отмечено улучшение условий влажности, температуры и pH почвы и подстилки, что подтвердило наблюдения ряда почвоведов [Карпачевский, 1977; Мина, 1960; и др.] об улучшении свойств почвы при удалении от ствола. Наилучшие условия для развития водорослей создаются между кронами деревьев, где в летний период почва лучше прогревается, где больше света, влаги и выше значение pH.
С удалением от стволов увеличивается видовая насыщенность и численность водорослей; большое число видов достигает массового развития, причем отмечается тенденция к увеличению доминантов и субдоминантов из зеленых водорослей и уменьшение роли желто-зеленых (табл. 22).
Таблица 22
| Место отбора проб | Глубина, см | 78-летные сосны | 93-летние ели | ||||||||
| сине-зеленые | зеленые | желто-зеленые | диатомовые | Всего | сине-зеленые | зеленые | желто-зеленые | диатомовые | Всего | ||
| У ствола | Подстилка 0 - 5 | - | 9 | 9 | - | 18 | - | 4 | 6 | - | 10 |
| - | 15 | 22 | - | 37 | - | 10 | 18 | - | 28 | ||
| В середине кроны | Подстилка 0 - 5 | 2 | 12 | 8 | - | 22 | - | 8 | 10 | 1 | 19 |
| 1 | 14 | 24 | - | 39 | - | 10 | 19 | - | 29 | ||
| Между кронами | Подстилка 0 - 5 | 4 | 9 | 15 | - | 28 | 2 | 5 | 9 | - | 16 |
| 4 | 14 | 22 | - | 40 | 2 | 10 | 17 | 1 | 30 | ||
Отмеченные закономерности неодинаково проявляются в разных лесах в связи с особеннями строения кроны разных деревьев. Сосны благодаря более рыхлому строению кроны пропускают под полог леса значительно больше света и влаги, чем кроны елей, что обусловливает более обильное развитие водорослей уже в районе середины крон. Для елей наиболее резкое увеличение числа клеток наблюдается только между кронами (табл. 23).
Таблица 23
| Место отбора проб | Глубина, см | 30-легние | Старые | ||
| сосняк | ельник | 78-летний сосняк | 93-летний ельник | ||
| У ствола | Подстилка 0 - 5 | 96,4 | 87,0 | 187,3 | 112,0 |
| 40,1 | 28,8 | 52,0 | 58,3 | ||
| В середине кроны | Подстилка 0 - 5 | 292,2 | 171,3 | 525,6 | 281,1 |
| 79,9 | 28,8 | 80,8 | 67,7 | ||
| Между кронами | Подстилка 0 - 5 | 497,2 | 393,9 | 771,4 | 550,7 |
| 88,8 | 37,4 | 93,1 | 86,0 | ||
Как видно из табл. 23, на различия распределения водорослей в пределах крон сосны и ели влияет возраст последних: различия наиболее заметны в молодых лесах. Определяя коэффициенты сходства разных лесов по Жаккару и по Малышеву, О. Я. Чаплыгина [1977] обнаружила наибольшее сходство между старыми сосновыми и еловыми лесами (Кж = 63,3%; Км = + 0,3).
Подтверждается отмеченная выше разница в составе и численности водорослей между подстилкой и почвой: в подстилке меньше разнообразие водорослей, но больше их численность (см. табл. 22, 23). Однако надо иметь в виду различный объемный вес подстилки и почвы: при пересчете на объем цифры могли быть более сходными.
Как отмечает О. Я. Чаплыгина [1977], в пределах кроны хвойных деревьев перераспределяется состав видов водорослей: по мере удаления от кроны возрастает роль сине-зеленых, хламидомонад и зеленых нитчатых. В частности, только на уровне середины крон и между кронами встречена Coccomyxa solorinae. Наиболее устойчивыми к неблагоприятным условиям которые создаются у стволов деревьев, оказались убиквисты из зеленых водорослей - Chlorococcum hypnosporum, Ch. minutum, Spongiococcum tet rasporum и убиквисты из желто-зеленых - Pleurochloris anomala и Botrydi opsis eriensis. Очень стойкими к затенению, пониженным значениям pH, недостаточной влажности и температуре почвы оказались также некоторые зеленые водоросли - Bracteacoccus minor, Chlorosarcinopsis minor, некоторые виды Chlamydomonas (Ch. gelatinosa, Ch. gloeogama, Ch. peterfii, Ch. snowiae), а из желто-зеленых водорослей - Pleurochloris imitans, Botrydiopsis arhiza, Ellipsoidion oocystoides, Gloeobotrys chlorinus, Characiopsis, Heterooccus chodatii.
Таким образом, в пределах проекций крон хвойных деревьев наблюдается четкая закономерность увеличения видовой насыщенности и численности клеток водорослей при удалении от ствола дерева к периферии и межкронным пространствам [Чаплыгина, 1977].
Образование группировок почвенных водорослей в ризосфере различных растений
Наряду с очень сильным косвенным влиянием высшие растения действуют на почвенные водоросли и прямым путем. Прямое влияние высшего растения на почвенные организмы проявляется в месте их непосредственного контакта - в ризоплане и ризосферной зоне. Это воздействие осуществляется прежде всего благодаря разнообразным веществам, выделяемым корнями растений в окружающую почву, а также особенностям физико-химического и биологического режимов, складывающихся в ризосферном слое почвы [Красильников, 1958]. Значительное количество работ посвящено накоплению в ризосфере древесных растений различных групп микроскопических организмов - бактерий, грибов, актиномицетов.
Почвенные водоросли как одна из групп организмов, накапливающихся в ризосферной зоне растений, привлекли внимание исследователей сравнительно недавно. Впервые влияние корней на почвенные водоросли было обнаружено Кацнельсоном [Katznelson, 1946] при изучении микроскопического населения ризосферы кормовой свеклы. После того проведено исследование ризосферной альгофлоры у многих растений, причем в основном изучению подвергались травянистые сельскохозяйственные растения. Работ, касающихся распространения почвенных водорослей в ризосфере древесных растений, чрезвычайно мало [Штина, 19546; Chodat, Chastain, 1957; Hadfield, 1960; Байрамова, 1965], при этом сведения об альгофлоре ризосферы лесных древесных пород отсутствуют полностью.
В большинстве случаев наблюдалось определенное влияние растений на распределение водорослей в ризосферном слое почвы [Штина, 1954а, б; 1955, 1956а, 1957, 1959б, 1960в; Gonzalves, Yalavigi, 1960; Hadfield, 1960; Katznelson, 1961; Cullimore, Woodbine, 1963; Штина и др., 1964; Байрамова, 1965, 1967; Мусаев, Умарова, 1967; Османова, 1974]. Это влияние выражается прежде всего в том, что количество клеток в ризосфере может до трех раз превышать содержание их в почве вдали от корней. Видовое разнообразие водорослей в ризосферной зоне оказывается меньше [Штина, 1955; Байрамова, 1965] или больше [Gonzalves, Yalavigi, 1960], чем вне ее. В некоторых случаях ризосферная и контрольная почвы, а также ризосферные зоны разных растений различаются соотношением отдельных групп водорослей [Штина, 1954б, 1956а; Байрамова, 1965, 1967; Steubing, 1967; Пивоварова, Платова, 1973]. При этом каких-либо специфичных видов, характерных для ризосферы растений, не обнаружено.
Влияние растений на водоросли ризосферы определяется, очевидно, прежде всего спецификой корневых выделений, которые используются водорослями в качестве источника пищи. Опытным путем было доказано положительное воздействие корневых выделений, самих корней, а также присутствия высших растений на рост и развитие водорослей [De, Sulaiman, 1950; Штина, 1954б, 1956а; Cullimore, Woodbine, 1963; Перминова, Третьякова, 1964]. Немаловажное значение при накоплении водорослей в ризосфере имеет механическое прикрепление их к корням растений и проникновение с последними в более глубокие слои почвы.
На развитие водорослей в ризосфере оказывает влияние температура и влажность почвы, в основном определяющие сезонную динамику численности ризосферных водорослей, реакция среды и структура почвы [Штина; 1955; Байрамова, 1965; Худякова, 1972], ее плодородие [Katznelson, 1946; Штина, 1960в], возраст и физиологическое состояние растений [Kalznelson, 1946; Штина, 1956а; Байрамова, 1965; Мусаев, Умарова, 1967], поражение болезнями и механические повреждения [Штина, 1956а; Новичкова-Иванова, 1968].
У ряда растений ризосферный эффект не обнаружен. Так, Кацнельсон и соавторы [Katznelson et al., 1956] не наблюдали накопления водорослей в ризосферной зоне яровой пшеницы, Э. А. Штина [19546] - у молодых растений гороха и яровых злаков, а Е. Н. Троицкая [1961] - у некоторых пустынных растений.
Воздействие растения на почвенные водоросли - это лишь одна сторона их взаимоотношений. У водорослей, как и у других почвенных микроорганизмов [Красильников, 1957, 1958], проявляется определенная специфика воздействия на высшее растение, хотя объем собственно ризосферной почвы относительно невелик. Например, в дерново-подзолистой почве при урожае пшеницы 30 ц/га объем ризосферной почвы составил всего 7% [Худякова, 1972]. Лишь небольшую часть от ризосферной почвы составляют населяющие ее организмы. Так, по приблизительным подсчетам, на 1 т ризосферной почвы приходится всего 4 г биомассы водорослей [Новичкова-Иванова, 1968]. Несмотря на это очень небольшое количество, важное значение водорослей, развивающихся в ризосфере, в значительной мере определяется их биохимическим влиянием на высшие растения и на различные группы микроорганизмов, находящихся в ризосферной почве.
Обнаружено, что многолетние и озимые растения, имеющие более развитую корневую систему с большей продолжительностью жизни, накапливают в почве большее количество водорослей [Штина, 1967]. Можно предполагать, что мощная многолетняя корневая система деревьев, создавая в прикорневом слое почвы специфические условия [Смирнов и др., 1964; Келеберда, 1970], должна оказывать определенное влияние на строение водорослевых группировок ризосферы и соответственно на формирование почвенной альгофлоры леса в целом.
Объектами для изучения альгофлоры в ризосфере различных древесных и травянистых растений выбирались [Алексахина, 1972а, б] деревья и травы, являющиеся основными растениями изучаемых лесных биогеоценозов или парцелл. Исследования проводились в следующих типах леса: ельнике- черничнике и сосняке-черничнике на подзолистой почве (ризосфера ели и сосны соответственно); дубо-ельнике волосистоосоковом на дерново-подзолистой почве (ризосфера ели, дуба, осины, а также осоки волосистой, сныти и папоротника); снытево-осоковой ясеневой дубраве на темно-серой лесной почве (ризосфера дуба и сныти). Кроме того, были собраны пробы в искусственно созданных на светло-каштановой почве лесозащитных полосах [Алексахина, 1981].
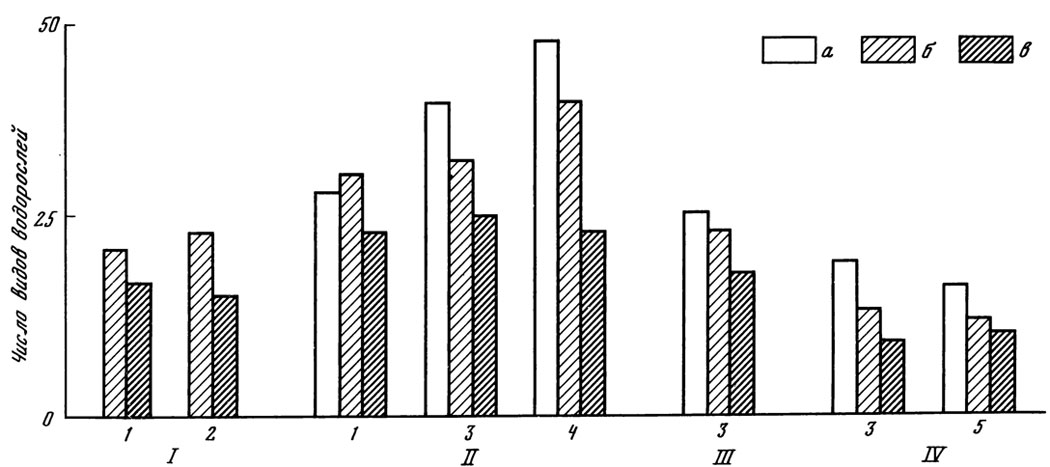
Рис. 12. Число видов водорослей в ризосфере древесных растений. I -подзолистая почва; II - дерново-подзолистая; III - темно-серая лесная; IV- светло-каштановая; 1 - ель; 2 - сосна; 3 - дуб; 4 - осина; 5 - смородина; а - смыв с корней; б - ризосфера; в - контроль
Не менее чем у пяти экземпляров каждого анализируемого вида растения собирались жизнеспособные тонкие корешки диаметром приблизительно в 1 мм. Встряхиванием освобождали корешки от прилипшей почвы, после чего прочно удерживающиеся на корнях почвенные комочки - ризосферную почву - скальпелем счищали в стерильные пакеты. С освобожденных от почвы корней ели, дуба, осины и сныти сразу был приготовлен смыв в колбах со средой Данилова для изучения альгофлоры, непосредственно примыкающей к корням растений (ризопланы). Для контроля каждый раз отбирались образцы почвы вдали от корней. Сезонная динамика численности и видового состава водорослей в ризосфере изучена на примере дуба, вяза, березы, смородины золотистой и типчака, произрастающих в зоне светло-каштановых почв.
Влияние корней и ризосферной почвы на водоросли сказывается уже в процессе выращивания культур. Так, раньше всего налеты водорослей появляются в колбах со смывом с корешков. В колбах, засеянных ризосферной почвой, рост водорослей начинается раньше, чем в колбах с контрольной почвой. Предполагается [Штина, 1954б] стимуляция роста и размножения водорослей самой ризосферной почвой. Возможно имеют значение вещества, вымываемые из корней растений и являющиеся стимулирующим и селективным фактором.
У всех исследованных древесных растений в ризосферной почве обнаружена более разнообразная альгофлора, чем в контрольной почве (рис. 12). В ризосфере возрастает по сравнению с контрольной почвой разнообразие зеленых и особенно желто-зеленых водорослей. В пределах одной почвенной разности число видов водорослей у лиственных пород выражено цифрами более высокого порядка, чем у хвойных. Соотношение количества водорослей в ризосфере и количества водорослей в контрольной почве (BP : BK), превышающее единицу в 1,7 - 4,6 раза (табл. 24), свидетельствует о накоплении водорослей в ризосфере, причем на одной и той же почве у лиственных пород оно обильнее, чем у хвойных. Об этом же говорят и абсолютные цифры: число клеток водорослей в 1 г ризосферной почвы осины почти в 2 раза больше, чем у ели.
Даже при сходном содержании водорослей в ризосфере одной и той же породы на разных почвах (например, в ризосфере ели на дерново-подзолистой и подзолистой почвах) ризосферный эффект увеличивается на менее плодородной почве. Такая же закономерность выявляется в зоне корневой системы дуба на трех разных почвах: дерново-подзолистой, темно-серой лесной и светло-каштановой - в последнем случае обнаруживается самый значительный эффект. Таким образом, древесные растения леса обусловливают накопление почвенных водорослей в своей прикорневой зоне.
В ризосфере разных растений формируются различные группировки водорослей. На подзолистой почве ель накапливает в своей ризосфере в основном виды Chlorophyta (наиболее массовые - Coccomyxa solorinae, Chlorella vulgaris), а также виды Xanthophyta (обилен Pleurochloris anomala). Другая хвойная порода - сосна, взятая в том же местообитании, накапливает в ризосферной зоне преимущественно зеленые водоросли, в частности виды Chlorococcum, Coccomyxa, Pleurastrum, Chlamydomonas.
На дерново-подзолистой почве в ризосферной зоне ели интенсивнее всего развиваются тоже зеленые и желто-зеленые водоросли, т. е. влияние растения на водоросли в общих чертах сохраняет свой характер на разных почвах. Подобные закономерности были обнаружены и в опытах с сельскохозяйственными растениями, в частности, со свеклой [Штина и др., 1964].
Дуб стимулирует в основном развитие зеленых водорослей, доминирующими из которых являются Chlorosarcina stigmatica, Gongrosira terricola, Chlorococcum humicola, виды Chlamydomonas. Желто-зеленые уступают по обилию зеленым водорослям, хотя число видов их тоже значительно. Из сине-зеленых встречаются Nostoc punctiforme.
Таблица 24
| Древесная порода | Место отбора проб | Число клеток (в тыс. на 1 г почвы) | BP : BK1 | |||
| сине-зеленых | зеленых + желто-зеленых | диатомовых | Всего | |||
| Подзолистые почвы | ||||||
| Ель | Ризосфера | 0 | 50,9 | <1,1 | 50,9 | 2,7 |
| Контроль | 0 | 18,9 | 0 | 18,9 | ||
| Сосна | Ризосфера | 0 | 26,5 | <1,2 | 26,5 | 2,2 |
| Контроль | 0 | 12,0 | 0 | 12,0 | ||
| Дерново-подзолистая почва | ||||||
| Ель | Ризосфера | <1,2 | 51,7 | <1,2 | 51,7 | 1,7 |
| Контроль | <1,3 | 30,7 | 0 | 30,7 | ||
| Дуб | Ризосфера | <1,3 | 77,4 | 2,5 | 79,9 | 2,3 |
| Контроль | <1,3 | 34,9 | 0 | 34,9 | ||
| Осина | Ризосфера | 18,9 | 79,2 | 1,2 | 99,3 | 3,1 |
| Контроль | <1,3 | 30,7 | 1,3 | 32,0 | ||
| Темно-серая лесная почва | ||||||
| Дуб | Ризосфера | 20,6 | 44,0 | 0 | 64,6 | 2,1 |
| Контроль | 7,8 | 22,3 | 0 | 30,1 | ||
| Светло-каштановая почва | ||||||
| Дуб | Ризосфера | 9,6 | 18,3 | 1,5 | 29,4 | 4,6 |
| Контроль | 4,8 | 1,6 | <1,7 | 6,4 | ||
| Береза | Ризосфера | 10,3 | 28,7 | 3,5 | 42,5 | 4,5 |
| Контроль | 0 | 7,6 | 1,8 | 9,4 | ||
| Вяз | Ризосфера | 7,5 | 31,6 | 0 | 38,6 | 4,1 |
| Контроль | 0 | 9,3 | 0 | 9,3 | ||
| Смородина | Ризосфера | 3,7 | 11,1 | 4,1 | 18,2 | 4,2 |
| Контроль | <1,8 | 4,3 | <1,6 | 4,3 | ||
1(BP : BK - соотношение количества водорослей в ризосферной и количества водорослей в контрольной почве.)
В ризосферной почве осины обнаруживается преимущественное развитие желто-зеленых водорослей, среди которых превалируют разнообразные Characiopsis. Встречены также виды Botrydiopsis, Pleurochloris, Monallantus, Nephrodiella, Bumilleriopsis. Из присутствующих здесь зеленых водорослей особенно обильны виды Chlamydomonas и Disporopsis pyrenoidifera. В ризосфере осины отмечено больше представителей Cyanophyta, чем у других деревьев на этой почве. Самой многочисленной из диатомей является Pin- nularia intermedia.
На темно-серой лесной почве накопление водорослей в ризосфере дуба происходит за счет видов Chlorophyta с преобладанием Stichococcus mirabilis, Hormidium nitens, Chlorosarcina stigmatica. Увеличивается значение одноклеточных желтозеленых водорослей, встречаются и нитчатки, в частности Heterothrix bristoliana. Возрастает число видов и степень развития сине-зеленых водорослей.
В ризосфере дуба, произрастающего на светло-каштановой почве, тоже преобладают зеленые водоросли Chlorococcum humicola, Chlorosarcinopsis minor и некоторые виды хламидомонад. Субдоминантом является одноклеточная желто-зеленая водоросль Botrydiopsis arhiza. Увеличивается развитие диатомей, особенно Hantzschia amphyoxis, и несколько меньше Pinnularia borealis.
В ризоплане и ризосфере вяза мелколистного на той же почве доминируют зеленые нитчатки - виды Chlorhormidium, Ulothrix, Gongrosira, а также Borodinella и виды Chlorella; обильны желто-зеленые водоросли. Береза бородавчатая, произрастающая на светло-каштановой почве, накапливает в своей ризосфере Pleurococcus naegelii, виды Heterococcus и некоторые виды Phormidium и Nostoc.
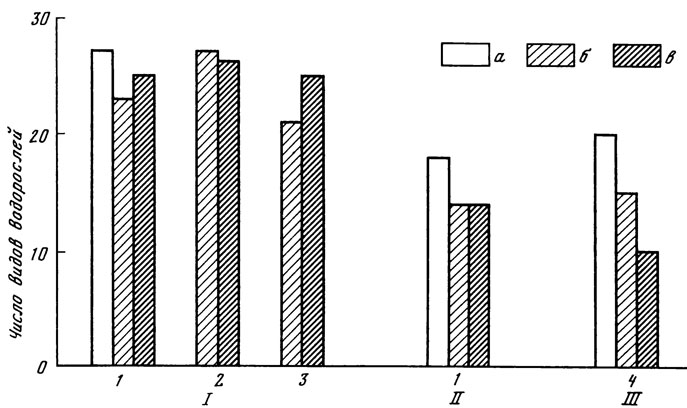
Рис. 13. Число видов водорослей в ризосфере травянистых растений. I - дерново-подзолистая почва; II - темно-серая лесная; III - светло-каштановая; 1 - сныть; 2 - осока; 3 - папоротник; 4 - типчак; а - смыв с корней; б - ризосфера; в - контроль
В ризосфере травянистых растений естественных лесных фитоценозов образуются водорослевые сообщества, менее насыщенные видами по сравнению с ризосферой древесных растений того же фитоценоза (рис. 13).
Число клеток водорослей в ризосфере трав меньше, чем у деревьев (табл. 25).
Ризосферный эффект обнаружен только у осоки на дерново-подзолистой и типчака на светло-каштановой почвах. У всех остальных травянистых растений коэффициент BP : BK меньше единицы, особенно невелик он у сныти. Надо отметить, что и при выращивании культур в колбах со смывом с корней сныти и засеянных ризосферной почвой сныти рост водорослей был самым медленным и слабым. Возможно, ингибирующее действие связано с какими-то токсическими веществами, выделяемыми снытью. Так, в опытах Н. Г. Кротовой [1968], настои листьев и корней сныти снижали всхожесть и энергию прорастания семян, а также рост проростков у осины, лиственницы и ели. Некоторые авторы [Parks, Rice, 1969], обнаружив ингибирующее действие сорных травянистых растений на развитие водорослей, объясняют его влияние фенольных соединений, выделяемых корнями и разлагающимися листьями растений. Определенное значение в распределении водорослей может иметь и разное строение корневых систем у рассматриваемых травянистых растений.
Таблица 25
| Вид растений | Место отбора проб | Число клеток (в тыс. на 1 г почвы) | BP : BK | |||
| сине-зеленых | зеленых и желто-зеленых | диатомовых | Всего | |||
| Дерново-подзолистая почва | ||||||
| Осока | Ризосфера | 10,8 | 27,6 | 0 | 38,4 | 1,3 |
| Контроль | 0 | 29,3 | 0 | 29,3 | ||
| Сныть | Ризосфера | <1,2 | 27,0 | 2,4 | 29,4 | 0,84 |
| Контроль | <1.3 | 34,9 | 0 | 34,9 | ||
| Папоротник | Ризосфера | <1,2 | 32,5 | 1,2 | 33,7 | 0,96 |
| Контроль | 1,3 | 34,9 | 0 | 34,9 | ||
| Темно-серая лесная почва | ||||||
| Сныть | Ризосфера | 4,3 | 19,7 | 0 | 24,0 | 0,71 |
| Контроль | 7,6 | 26,0 | 0 | 33,6 | ||
| Светло-каштановая почва | ||||||
| Типчак | Ризосфера | 12,5 | 1,4 | 5,1 | 19,5 | 5,3 |
| Контроль | 3,7 | 1,7 | 1,5 | 3,7 | ||
Однако даже при небольших различиях в количестве клеток и числе видов водорослей, травянистые растения тоже определенным образом влияют на строение водорослевых сообществ ризосферы. В ризосфере сныти на дерново-подзолистой почве альгофлора отличается преобладанием зеленых водорослей. Осока стимулирует значительное развитие желто-зеленых водорослей; 30% всех обнаруженных здесь видов приходится на долю сине-зеленых, в основном различных видов Phormidium. В ризосферной зоне папоротника превалируют представители Chlamydomonas и других зеленых.
Ризосфера сныти, произрастающей на темно-серой лесной почве, богата одноклеточными зелеными и желто-зелеными водорослями (в основном виды-убиквисты). Вместе с тем отмечено обильное разрастание сине-зеленой водоросли Schizothrix lardacea f. diplosiphon.
В ризосфере типчака на светло-каштановой почве доминируют разнообразные сине-зеленые водоросли, из которых самыми обильными являются виды Oscillatoria и Phormidium. Их дополняют одноклеточные зеленые из Chlorococcales и многочисленные диатомовые водоросли. Таким образом, каждое изученное растение создает довольно своеобразную группировку водорослей.
При сравнении основных альгосинузий в почве парцелл и в ризосфере соответствующих парцеллообразующих растений было обнаружено значительное совпадение. Так, насыщенность видами в ризосфере лиственных пород оказалась выше, чем в ризосфере хвойных. В лиственных парцеллах (особенно в осиново-снытевой) тоже обнаружено большее разнообразие водорослей, чем в парцелле, контролируемой елью. В ризосфере ели, как и в почве елово-волосистоосоковой парцеллы, из обильно развивающихся представителей Chlorophyta самыми массовыми являются тоже виды Chlamydomonas и хлорококковые водоросли. В ризосферной зоне осины и в осиново-снытевой парцелле превалируют желто-зеленые водоросли с доминированием Characiopsis и повышается значение видов Cyanophyta. Сходные группировки водорослей оказываются наиболее развитыми в ризосфере дуба и в почве парцеллы, сформированной дубом. Можно предположить, что, обусловливая развитие своеобразных группировок водорослей в ризосфере, растение определенным образом влияет на формирование альгофлоры всего биогеоценоза. Однако аналогичного соответствия между водорослевыми сообществами ризосферы травянистых растений и альгофлорой парцелл не обнаружено.
Влияние растения на водоросли носит довольно постоянный характер. Например, в ризосфере дуба, исследованного на трех типах почв (дерново-подзолистой, темно-серой и светло-каштановой), доминирующими оказались зеленые водоросли, в ризосфере ели, произрастающей на двух типах почв (подзолистой и дерново-подзолистой) - зеленые и желто-зеленые. Стабильность влияния растений на водоросли отмечена и для сельскохозяйственных растений [Штина и др., 1964]. Однако какие-либо виды водорослей, специфичные только для данного вида высшего растения, обнаружить не удалось.
Однако на ризосферную альгофлору оказывают влияние и свойства почвы, придавая определенный оттенок характеру водорослевого сообщества. Например, ранее было установлено, что для дерново-подзолистой почвы под дубо-ельником волосистоосоковом характерно обильное развитие хламидомонад. В ризосферной альгофлоре всех исследованных деревьев и трав на дерново-подзолистой почве обнаружены виды хламидомонад, чаще всего входящие в доминирующий комплекс. В то же время в темно-серой лесной почве под снытево-осоковой дубравой выявлено лишь незначительное количество этих водорослей. Единичными экземплярами представлены виды Chlamydomonas и в ризосфере изученных на этой почве дуба и сныти. Кроме этого, в ризосфере растений одного и того же вида преобладающие отделы водорослей представлены разными видами в зависимости от условий местообитаний. Так, если в ризосфере дуба на дерново-подзолистой почве наибольшее развитие среди зеленых водорослей получают хлорококковые, тетраспоровые водоросли и в значительной степени хламидомонады, то на темно-серой лесной почве обилие зеленых обеспечивается в основном улотриксовыми и хлорококковыми водорослями. В совокупности с этим признаком появление желто-зеленых нитчаток и возрастание роли сине-зеленых водорослей составляют отличительные особенности ризосферной альгофлоры дуба на темно-серой лесной почве, которые прежде всего могут быть связаны с влиянием свойств почвы. Почва как бы дает общий фон, матрицу, на основе которой создается водорослевый комплекс ризосферы.
Характер влияния растения на водоросли, выражающийся в накоплении последних в ризосфере растений, определяется, по-видимому, спецификой корневых выделений, которые используются водорослями в качестве источников пищи. Немаловажное значение имеет механическое прикрепление водорослей к корням растений и проникновение их в более глубокие слои почвы. Значительную роль играют особенности режима в ризосферном слое почвы: повышенное содержание минеральных солей [Канивец, 1950], более благоприятные показатели влажности и рН, наличие биологически активных веществ [Красильников, 1958], большое количество общего и воднорастворимого гумуса, высокая подвижность органического вещества, большая интенсивность дыхания, повышенная ферментативная активность [Смирнов и др., 1964; Келеберда, 1970; Славина; 1971]. Кроме того, на корнях растений образуется очень сложный комплекс организмов, состоящий из высших и микроскопических грибов, актиномицетов и бактерий, что для древесных растений показано многими исследователями [Рунов, Мишустина, 1960; Рунов, Еникеева, 1961; Рунов, Егорова, 1962; Большакова, 1964; Еникеева, 1966; Егорова, Рагуотис, 1968; Егорова, Степанова, 1972; и др.].
Таблица 26
| Вид растений | Место отбора проб | Количество микроорганизмов (в млн. на 1 г почвы) | ||||
| МПА | KAA | Флуоресцирующие бактерии (МПА) | Спорообразующие бактерии (МПА + сусло) | Актиномицеты (KAA) | ||
| Дуб | Ризосфера | 5,9 | 7,0 | 1,4 | 2,4 | 28 |
| Контроль | 4,6 | 12,7 | 1,2 | 1,9 | 6,7 | |
| Сныть | Ризосфера | 16,6 | 118 | 11,2 | 1,0 | 14 |
| Контроль | 4,6 | 12,7 | 1,2 | 1,9 | 6,7 | |
Проведенные совместно с С. В. Егоровой микробиологические анализы ризосферной зоны дуба и сныти, произрастающих на темно-серой лесной почве, показали, что ризосфера изученных растений богаче заселена микроорганизмами, чем окружающая внеризосферная почва (табл. 26). Отмечены также различия между дубом и снытью в составе ризосферной микрофлоры. Если в ризосферной зоне сныти доминирующими являются неспорообразующие бактерии и бактериальная флора, развивающаяся на крахмало-аммиачном агаре, то в ризосфере дуба развиваются преимущественно спорообразующие бактерии и актиномицеты, т. е. организмы, разлагающие более сложные органические соединения.
Таблица 27
| Фитоценоз | Место отбора проб | Микроорганизмы, млн. клеток в 1 г почвы | Водоросли, клеток в 1 почвы |
| Бор травяной | Контроль | 0,99 | 63,2 |
| Ризосфера сосны | 12,1 | 153,3 | |
| Ризосфера ели | 10,9 | 128,8 | |
| Пихтово-еловая согра | Контроль | 0,9 | 54,6 |
| Ризосфера ели | 6,7 | 73,3 | |
| Сосняк сфагновый | Контроль | 0,3 | 34,6 |
| Ризосфера сосны | 3,9 | 67,2 |
Выявились различия в воздействии корневых систем одного и того же растения на гетеротрофные микроорганизмы и водоросли: в ризосфере дуба создаются благоприятные условия для развития как микробов, так и водорослей, в ризосфере сныти - только для гетеротрофных микроорганизмов. Вероятно, повышенная микробиологическая активность и развитие определенных групп микроорганизмов в ризосферных зонах разных растений определенным образом влияет на характер водорослевых группировок ризосферы. К. И. Еропкин [1977], изучая консортивные связи высших и низших растений, показал соотношения между микоризой сосны и ели и эпифитной ризосферной микрофлорой (табл. 27). Данные относятся к июлю 1975 г.
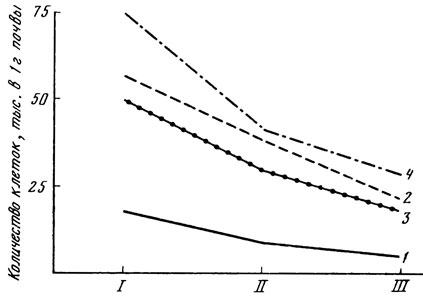
Рис. 14. Динамика численности водорослей в ризосфере разных растений на светло-каштановой почве. I - весна; II - лето; III - осень; 1 - контрольная почва; 2 - ризосфера вяза; 3 - ризосфера дуба; 4 - ризосфера березы
На примере лесных насаждений на светло-каштановой почве показано, что характер водорослевых группировок меняется в течение вегетационного периода. Происходит перегруппировка в составе доминирующего комплекса водорослей ризосферы: весной, в самый влажный из всех исследуемых периодов, довольно обильно вегетируют влаголюбивые виды зеленых (в частности, хламидомонады и нитчатки из Ulotrichales) и желтозеленых водорослей; в более сухие летний и осенний периоды из зеленых преобладают виды-убиквисты из Chlorococcales; к осени усиливается развитие синезеленых водорослей.
В течение сезона в ризосферной зоне растений отмечаются колебания числа видов и количества клеток водорослей. Наибольшее разнообразие (табл. 28) и максимальная численность (рис. 14) водорослей в ризосфере найдены весной, причем это соответствует самой высокой влажности почвы. Летом и осенью до периода дождей почва подсыхает так сильно, что в контроле содержание водорослей падает до 1 - 6 тыс. на 1 г почвы, тогда как в ризосфере оно доходит до 20 - 30 тыс. на 1 г.
Интересно отметить, что Р. А. Османова [1974] не нашла водорослей в контрольной пустынной почве Туркмении, но обнаружила их в ризосфере пяти солянковых растений в количестве от 1,4 до 101,6 тыс. клеток в 1 г почвы. Здесь, возможно, сказывается благоприятное влияние повышенной влажности почвы вблизи корней. С другой стороны, Л. А. Байрамова [1965] во влажных субтропиках Азербайджана нашла в ризосфере чайного куста лишь небольшое превышение численности водорослей по сравнению с контролем (отношение BP : ВК колебалось от 1,1 до 1,5).
Можно высказать предположение, что накопление водорослей в ризосфере обязано не только и, может быть, не столько корневым выделениям, но и в особенности - повышенной влажности прикорневого слоя почвы.
Как видно из табл. 28, во все сезоны ризосферная почва богаче контрольной по числу видов; в смыве с корней (ризоплане) число видов до 2,3 раза больше, чем в контроле. Наиболее разнообразен спектр видов водорослей в ризоплане типчака: весной 11 видов сине-зеленых, 8 - зеленых, 6 - желто-зеленых и 7-диатомовых. Подтверждается отмеченная ранее закономерность - накопление диатомей в ризосфере злаков [Штина, 1954б].
Таблица 28
| Вид растения | Зона исследования | Весна | Лето | Осень |
| Дуб черешчатый | Смыв с корней | 24 | 18 | 16 |
| Ризосфера | 19 | 13 | 13 | |
| Контроль | 14 | 9 | 7 | |
| Смородина золотистая | Смыв с корней | 22 | 16 | 15 |
| Ризосфера | 21 | 12 | 12 | |
| Контроль | 18 | 10 | 7 | |
| Типчак | Смыв с корней | 32 | 20 | 20 |
| Ризосфера | 24 | 17 | 12 | |
| Контроль | 21 | 12 | 9 | |
| Вяз мелколистный | Смыв с корней | 23 | 16 | 12 |
| Ризосфера | 20 | 13 | 12 | |
| Контроль | 17 | 8 | 11 | |
| Береза бородавчатая | Смыв с корней | 23 | 19 | 16 |
| Ризософера | 18 | 18 | 13 | |
| Контроль | 21 | 11 | 8 |
Очевидно, обнаруженные взаимоотношения высших растений с альго-флорой ризосферы можно рассматривать как один из вариантов консортивных отношений, где растения (дерево, куст, трава) служат детерминантом консорции, а сопутствующие ему определенные по численности и составу ризосферные водоросли, относящиеся к особой функциональной группе - эккрисотрофам [Работнов, 1974в], являются консортами. Своеобразные ризосферные группировки водорослей, созданные исследованными растениями, могут, вероятно, влиять на конкурентные отношения самих растений, участвовать в аллелопатических взаимоотношениях в биогеоценозе [Работнов, 1974а, в].
Следует обратить внимание на новые методы изучения ризопланы. В частности, интересно определить плотность водорослей на поверхности корней, как это сделала И. С. Гузева [1978] для микроорганизмов, показав, что, хотя последние заселяли лишь 1,4 - 10% поверхности корней, плотность их на единицу поверхности корня была на 2 - 3 порядка выше, чем плотность на поверхности почвенных частиц.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'