Распределение водорослей в профиле лесных почв
Водоросли - фототрофные организмы, но распространение их в почвах не ограничивается верхним слоем толщиной в несколько миллиметров, куда проникают лучи света видимой части спектра. Убедительно доказано присутствие водорослей в более глубоких горизонтах почвы, при этом разными методами показано существование там водорослей в вегетативном состоянии [Bristol-Roach, 1927; Petersen, 1935; Голлербах, 1936; Штина, 1959а; Аристовская, 1965; Звягинцев, 1965; и др.]. Представления о существовании жизнеспособных водорослей в толще почвы подтверждаются также фактами факультативной гетерографии у зеленых [Parker, 1961], желто-зеленых [Belcher, Miller, 1960; Casselton, 1966] и сине-зеленых [Кузьменко, 1977, 1981] водорослей.
Использование водорослями простых Сахаров и органических кислот на свету и в темноте является систематическим признаком [Bold, Wynne, 1978]. Меттинг [Metting, 1981] указывает, что, по данным Болда и его учеников, 40 - 50% зеленых и желто-зеленых водорослей являются хемо-гетеротрофами. У облигатно-фототрофных сине-зеленых водорослей темновой образ жизни приводит к различным типам деградации клеток [Гусев, Никитина, 1979]. В то же время описано около 30 видов, способных к гетеротрофному росту. Авторы предполагают наличие среди сине-зеленых водорослей (цианобактерий) всех переходных форм от облигатной фототрофии до гетеротрофии.
Глубинная флора почвенных водорослей формируется из представителей, обитающих на поверхности и в верхних горизонтах почвы; в некоторых случаях принимает участие и водная альгофлора (например, при затоплении почв речной водой или при орошении). Пассивное проникновение водорослей в глубь почвы может происходить разными путями: заносом клеток током воды, микро- и мезофауной, корневыми системами растений, при обработке почвы [Голлербах, 1936; Tchan, Whitehouse, 1953; Штина, 1959а; Reisigl, 1964; Голлербах, Штина, 1969].
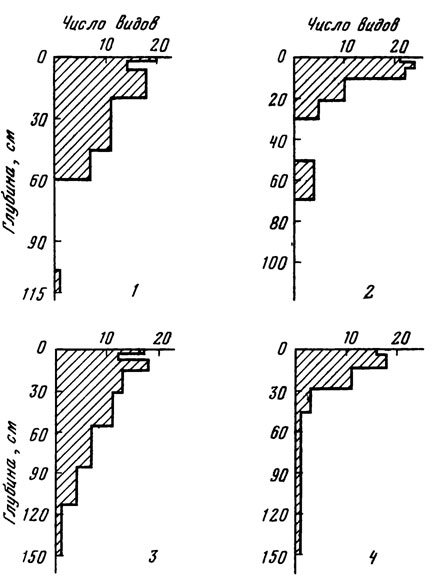
Рис. 3. Число видов водорослей в профиле почв под коренными типами леса. 1 - почва под ельником-черничником; 2 - почва под дубо-ельником волосисто-осоковым; 3 - почва под сосняком чернично-зеленомошным; 4 - почва под снытево-осоковой в ясеневой дубравой
Во всех изученных нами лесных почвах с глубиной происходят уменьшение видового разнообразия (рис. 3) и снижение численности водорослей (рис. 4), причем изменения по профилю числа видов и количества клеток водорослей различаются по характеру. Падение численности водорослей начинается сразу с поверхности почвы, при этом в верхнем 10-сантиметровом слое оно происходит особенно резко (рис. 4).
Наибольшее количество водорослей во всех типах леса обнаружено в подстилке, причем в тех случаях, когда подстилка была разделена морфологически на два слоя (в ельнике-черничнике и сосняке чернично-зеленомошном), населенность верхнего 1,5 - 2-сантиметрового слоя до 3 раз превышала количество клеток водорослей нижнего слоя (табл. 11). Содержание водорослей в горизонте А1 в 3,5 - 7 раз меньше, чем в горизонте А0, в горизонте А2 оно уменьшается примерно еще во столько же раз по сравнению с вышележащим горизонтом. Некоторые авторы [Гаврилов, 1950] вообще не обнаружили водоросли в горизонте А2.
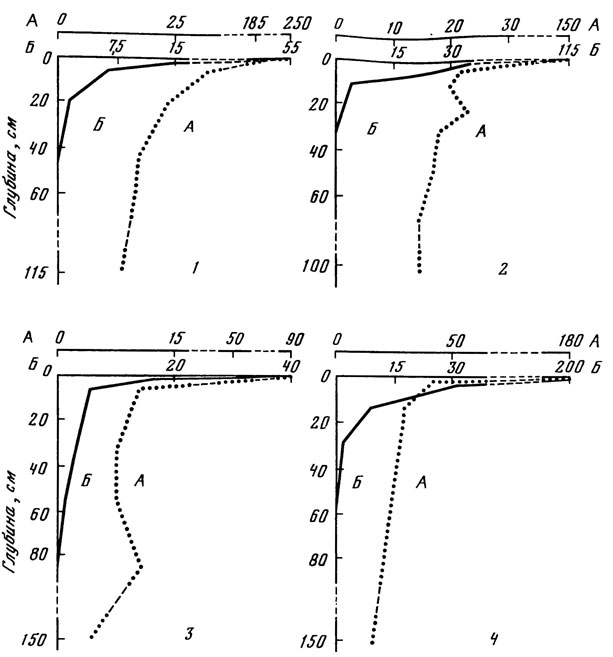
Рис. 4. Количество водорослей в профиле почвы под коренными типами леса. 1 - ельник-черничник; почва сильноподзолистая; суглинистая (24/IX); 2 - дубо-ельник волосистоосоковый; почва дерново-подзолистая, суглинистая (10/X); 3 - сосняк чернично-зеленомошный; почва дерново-подзолистая, пылевато-песча- ная (13/Х); 4 - дубрава ясеневая снытево-осоковая; почва темно-серая лесная, суглинистая (30/IX). А - влажность, %; Б - количество клеток, тыс. в 1 г абс. сухой почвы
Закономерное уменьшение по профилю числа видов начинается лишь с некоторой глубины от поверхности, причем эта глубина зависит от типов почв и леса. В подстилке и самом верхнем горизонте почвы отмечаются значительные колебания в количестве видов водорослей. Важно отметить, что подстилка часто оказывается обедненной видами по сравнению с нижележащими горизонтами. В некоторых случаях максимальное число видов отмечается на глубине 1 - 5 см. В хвойных лесах наиболее богатая альго-флора бывает приурочена к еще большей глубине - к слоям 5 - 10 см и даже 10 - 15 см. В этих же слоях обнаруживается иногда и возрастание численности водорослей. Эти явления могут быть связаны, с одной стороны, с вымыванием водорослей осадками, с другой стороны, в этих слоях возрастает разнообразие хламидомонад и желто-зеленых водорослей, приспособленных к жизни в более глубоких слоях почвы. Немаловажное значение имеют физико-химические свойства самой почвы. Так, под ельником-черничником слой 1,5 - 6 см в большинстве случаев оказывался наименее насыщенным видами водорослей по сравнению с выше- и нижележащими слоями. Вероятно, этот слой, представляющий нижнюю полуразложившуюся часть подстилки, в силу своей уплотненности, более высокой кислотности и плохой смачиваемости [Орлов, 1966а], а также вследствие колебаний влажности и температуры оказывается мало пригодным для развития водорослей. Случаи возрастания численности и количества видов водорослей на глубине 10 - 20 см по сравнению с поверхностью наблюдались и для других типов почв и фитоценозов [Bristol-Roach, 1927, 1928; Lund, 1945; Зауер, 1956; Куликова, 1965; Голлербах, Штина, 1969; Штина, 1974] . Л. М. Зауер считает, что это явление свидетельствует о микростратификации почв и обусловливается действием различных факторов. Распределение водорослей глубже по профилю происходит таким образом, что горизонты, содержащие водоросли, могут чередоваться с горизонтами, в которых водоросли отсутствуют. Это наблюдается, например, под ельником-черничником и дубо-ельником волосистоосоковым (см. рис. 2).
Максимальная глубина проникновения водорослей в почву под разными типами леса меняется в значительных пределах. Так, методом прямого счета было выявлено, что глубина, до которой еще обнаруживаются клетки водорослей, колеблется от 30 см в дерново-подзолистой суглинистой почве под дубо-ельником (см. рис. 4) до 84 см в дерново-подзолистой песчаной под сосняком чернично-зеленомошным (см. рис. 4). Методом культур присутствие водорослей выявляется по всему профилю почвы вплоть до глубины 150 см, при этом вместо доминантов верхних горизонтов - видов Chlorophyta - в нижних слоях обнаруживаются преимущественно одноклеточные желто-зеленые: Characiopsis saccata, Ch. minuta, Ch. gracilis, Botrydiopsis arhiza. В разные сроки наблюдается неодинаковое распределение водорослей по слоям почвы: если в сухие периоды с глубиной происходит уменьшение числа видов и количества клеток водорослей, то в периоды обильных дождей отмечается некоторое накопление водорослей в слоях 5 - 10 и 10 - 15 см.
Глубина распространения водорослей зависит от свойств и строения почвы. В частности, чем легче почва по механическому составу, тем глубже проникают в нее водоросли и тем более равномерно происходит снижение числа видов и количества клеток по профилю. Глубина проникновения водорослей в почву может зависеть также от особенностей роста и развития корней высших растений [Flint, 1958], наличия вечной мерзлоты [Бусыгина, 1972] и ряда других факторов. Плотные горизонты могут затруднять занос водорослей в более глубокие слои и горизонты почвы. Полученная в наших исследованиях глубина проникновения водорослей в почву, равная 1,5 м, совпадает с показателями максимальной глубины, указанной для целинных почв и другими авторами [Голлербах, 1953; Поцене, 1961; Штина, 1965; Гаухман и др., 1970].
Несмотря на распространение водорослей в профиле почвы на значительную глубину, наибольшее развитие и активная деятельность их обнаруживаются в подстилке и самых верхних слоях почвы.
В большинстве почвенно-альгологических работ приводятся обобщенные данные по всем изученным слоям или описываются водоросли только собственно почвенных горизонтов. Сведения об альгофлоре лесных подстилок встречаются относительно редко [Зауер, 1956; Штина, Ройзин, 1966; Голлербах, Штина, 1969; Чаплыгина, 1977] . Вместе с тем подстилки представляют специфическое природное образование, играющее в жизни леса важную роль [Морозов, 1949; Зонн, Алешина, 1953; Ремезов и др., 1959, 1964; и др.]. Опад и образующаяся из него подстилка - особый биогеоценогический горизонт [Работнов, 1974б] - являются продуктом жизнедеятельности леса и важным звеном в процессе взаимодействия между лесной растительностью и почвой. Подстилки аккумулируют и после разложения возвращают почве значительные количества органических и минеральных веществ, играют важную роль в регулировании. гидротермического и воздушного режимов почв, в большой мере определяя характер ее живого населения.
Обилие органического вещества, аккумуляция зольных элементов питания растений, специфические особенности светового и гидротермического режимов, высокая заселенность соответствующей гетеротрофной микрофлорой, представители которой часто являются антагонистами водорослей, - все эти факторы оказывают селективное влияние и на альгофлору лесных подстилок и соответственно почвы. Значительный слой опада может мешать развитию типичных наземных водорослей, затенять почву, препятствуя проникновению в нее света [Штина, 1965], и тем самым снижать развитие водорослей и обеднять их видовой состав [Новичкова-Иванова, 1969; Чаплыгина, 1977]. На важную роль опада в Нормировании водорослевых группировок лесных почв указывали Э. А. Штина и М. Б. Ройзин [1966], М.М. Голлербах и Э.А. Штина [1969].
Подробные сведения о значении подстилок приводятся в главе III в связи с вопросом о влиянии высших растений на почвенные водоросли. Здесь мы указываем особенности распределения водорослей в подстилках и в верхних слоях почвы. Показана стратификация разных групп водорослей в почвах под коренными типами леса в период максимальной численности клеток (см. табл. 11).
Таблица 11
| Горизонты, глубина, см | Сезон | Число клеток (в тыс. на 1 г почвы) | |||
| сине-зеленых | зеленых + желто-зеленых | диатомовых | Всего | ||
| Ельник-черничник | |||||
| А'0 0 - 1,5 | Лето | <1,6 | 77,3 | 1,6 | 78,9 |
| А"0 1,5 - 6 | <1,5 | 21,2 | 0 | 21,2 | |
| А2 6 - 10 | 0 | 13,5 | 0 | 13,5 | |
| А2 10 - 15 | 0 | 3,8 | 0 | 3,8 | |
| Дубо-ельник волосистоосоковый | |||||
| А0 0 - 1 | Осень | 21,2 | 93,4 | 5,2 | 119,8 |
| А1 1 - 5 | <1,6 | 41,3 | 0 | 41,3 | |
| А1А2 5 - 10 | 0 | 22,7 | 0 | 22,7 | |
| А2 10 - 15 | 0 | 9,1 | 0 | 9,1 | |
| Сосняк чернично-зеленомошный | |||||
| А'0 0 - 2 | Лето | <1,7 | 69,0 | 0 | 69,0 |
| А"0 2 - 5 | 0 | 25,8 | 0 | 25,8 | |
| А1А2 5 - 10 | 0 | 16,7 | 0 | 16,7 | |
| А1А2 10 - 15 | 0 | 20,8 | 0 | 20,8 | |
| Снытево-осоковая ясеневая дубрава | |||||
| А0 0 - 2 | Осень | 113,7 | 80,2 | 14,4 | 208,3 |
| А1 2 - 5 | 18,2 | 48,2 | 1,5 | 67,9 | |
| А1 5 - 10 | <1,6 | 28,2 | 0 | 28,2 | |
| А1 10 - 15 | 0 | 19,0 | 0 | 19,0 | |
По нашим данным, подстилки являются основным местом развития сине-зеленых водорослей (см. табл. 11). В почве виды Cyanophyta и участие их в доминирующем комплексе наблюдается лишь в верхних 5 см горизонта A1 в дубраве и смешанном лесу. Случаи проникновения сине- зеленых водорослей в более глубокие горизонты почвы не наблюдались (табл. 11). Большинство авторов рассматривают сине-зеленые водоросли как приуроченные к поверхностным или самым верхним слоям почвы [Petersen, 1935; Ваулина, 1956; Штина, 1959а; 1965; Schwabe, 1963; Голлербах, Штина, 1969; Чаплыгина, 1975, 1977].
Другой группой водорослей, развитие которых приурочено к подстилке, являются диатомеи, встречающиеся в почве лишь спорадически и в самых верхних слоях. Виды Bacillariophyta как наиболее резко реагирующие на изменение глубины охарактеризовывают и другие авторы [Lund, 1945; Ваулина, 1956; Пивоварова, 1968; и др.]. В то же время Франсе [France, 1913] считает, что диатомеи, в частности Navicula, избегают света и более распространены в нижних слоях почвы. В наших исследованиях в почвенных горизонтах попадались только пустые панцири диатомей.
Из других отделов водорослей в комплекс видов, доминирующих в подстилках, входят зеленые нитчатки из Ulotrichales: виды Chlorhormidium, Stichococcus, Ulothrix. Постоянным компонентом водорослевого сообщества подстилки во всех изученных биогеоценозах является тетраспоровая водоросль Соссоmуха: в хвойных лесах чаще всего это Соссоmyxa solorinae, а в дубовом - С. dispar.
В то же время видовой состав водорослей в подстилке оказывается обедненным хламидомонадами и желтозелеными водорослями. Последние Ланд [Lund, 1962] отнес к "теневым" формам. Неустойчивость к свету видов Xanthophyta отмечалась и Флинт [Flint, 1958]. Данные, подтверждающие "теневыносливость" желто-зеленых водорослей, получены О. Я. Чаплыгиной [1975, 1977]. Вероятно, в первую очередь этим свойством видов Xanthophyta можно объяснить тот факт, что в подстилке происходит развитие лишь ограниченного числа видов. Такими видами являются, например, Botrydiopsis arhiza и нитчатая желтозеленая водоросль, не идентифицированная до вида, - в подстилке дубравы, Gleobotrys ellipsoideus - в горизонте А0 ельника-черничника и два вида Characiopsis : Ch. saccata и Ch. minutissima, участвующие в доминирующем комплексе водорослей подстилки сосняка чернично-зеленомошного. Виды Chlamydomonas развиваются слабо только в самом верхнем слое подстилки. В нижнем торфянистом слое подстилки (1,5 - 5 см) хламидомонады встречаются уже в заметных количествах и даже входят в доминирующий комплекс водорослей, хотя и здесь их видовое разнообразие меньше, чем в почвенных слоях.
С выпадением многих видов Xanthophyta и Chlamydomonas из состава альгофлоры подстилки может быть частично связана ее видовая обедненность по сравнению с нижележащими почвенными слоями.
В почве по обилию и видовому разнообразию превалируют зеленые и желто-зеленые водоросли, при этом в верхних горизонтах обильно развиваются одноклеточные, нитчатые и колониальные формы этих отделов. Из одноклеточных зеленых водорослей здесь многочисленны виды Chlorococcum, Chlorella, Radiosphaera, Chlorochytrium; их дополняют разнообразные хламидомонады. По всей населенной хламидомонадами почвенной толще распространяются виды: Chlamydomonas gloeogama, Ch. globosa, Ch. snowiae, Ch. gelatinosa, Ch. oblongella, при этом три первых являются массовыми и на глубине 45 см. Наибольшее число видов Chlamydomonas обнаружено в слое 5 - 15 см, затем их разнообразие постепенно начинает уменьшаться, а глубже 55 см, куда проникает только Chlamydomonas platyrrhyncha, хламидомонады обнаружены не были.
Одноклёточные желто-зеленые водоросли представлены в верхних горизонтах почвы видами Botrydiopsis, Ellipsoidion, Pleurochlor is, Characiopsis, Monallantus, которые в большинстве случаев входят в число доминирующих форм. Желто-зеленые нитчатки в почве имеют небольшую глубину распространения, ограничиваясь лишь верхним слоем 2 - 13 см, при этом встречаются не во всех лесах.
Колониальные формы водорослей, в число которых из зеленых входят Chlorosarcina stigmatica, Chlorosarcinopsis minor, а из желто-зеленых - Gloeobotrys ellipsoideus, Botryochloris cumulata, B. minima, глубже 10 - 15 см почти не проникают. Нитчатые зеленые водоросли в почвенных слоях представлены видами Ulothrix, Chlorhormidium, Gongrosira, Stichococcus, Leptosira, Pleurastrum, из них в доминирующий комплекс водорослей постоянно входит Chlorhormidium flaccidum var. nitens, превалирующий до глубины 15 см, реже - Gongrosira terricola, Ulothrix variabilis, Stichococcus minor. Распространение зеленых нитчаток в основном ограничено верхним 5 - 10 (иногда 15)-сантиметровым слоем почвы, лишь Ch. flaccidum var. nitens в небольших количествах встречается до 28 см. Только один раз в почве под ельником-черничником в горизонте В2С на глубине 100 - 114 см был обнаружен представитель порядка Ulotrichales - Pleurastrum terrestre, причем это был единственный присутствующий в этом горизонте вид водорослей. Случаи, когда глубинными формами оказываются зеленые или желто-зеленые нитчатки, и в литературе описываются довольно редко [Ваулина, 1956; Штина, 1959а; Поцене, 1970]. Кроме указанного выше случая, во всех остальных изученных нами типах почвы до самых нижних горизонтов проникают только одноклеточные водоросли: Chlorella vulgaris, Botrydiopsis arhiza, Characiopsis saccata, Ch. minuta, Ch. gracilis, Ch. lunaris, Ellipsoidion regulare, из них глубины 1,5 м достигает только Botrydiopsis arhiza.
Размеры водорослей, обнаруженных нами в лесных почвах, часто соответствуют минимальным значениям, указанным в диагностических признаках для данного вида, или в некоторых случаях бывают даже мельче, чем приведено в описаниях при совпадении всех остальных признаков. Мелкоклеточность почвенных водорослей является одним из приспособлений их к условиям жизни в такой специфической среде обитания, как почва. На эту отличительную черту почвенных водорослей указывают многие исследователи [Bristol, 1920; Gistl, 1932; Голлербах, 1936; Lund, 1945; Болышев, Манучарова, 1947; Brendemuhl, 1949; и др.]. Мелкоклеточность можно считать наследственно стойким признаком, характерным для почвенных водорослей [Голлербах, 1949]. Мелкие размеры почвенных водорослей, очевидно, облегчают их передвижение между частицами почвы и способствуют лучшему выживанию при засухе, недостатке питательных веществ и воздействии других неблагоприятных факторов внешней среды [Штина, Голлербах, 1976а, б], а также, согласно Д. Г. Звягинцеву [1973], адсорбции на поверхности почвенных частиц.
Таким образом, при вертикальном распределении почвенных водорослей одним из основных факторов является градиент глубины, определяющий интенсивность освещения слоев почвы. Влияние этого фактора выражается в уменьшении с глубиной как количества клеток, так и числа видов водорослей. Это положение подтверждается, в частности, тем фактом, что разные слои одного и того же почвенного горизонта (например, слои 6 - 10 и 10 - 15 см горизонта А2 мощного подзола, слои 6 - 10 и 10 - 15 см горизонта А1/А2 дерново-подзолистой пылевато-песчаной почвы; слои 2,5 - 5, 5 - 10 и 10-15 см горизонта A1 темно-серой лесной почвы) значительно отличаются друг от друга численностью (см. табл. 11) и видовой насыщенностью водорослей: нижележащие слои содержат до 10 раз меньше клеток водорослей и до 2 раз меньше видов, чем вышележащие (см. рис. 3, 4). Вместе с тем физико-химические свойства почвы, погодные условия, состояние высших растений (особенно корневой системы), определенные свойства различных видов водорослей и ряд других факторов смещают прямое воздействие градиента глубины на распределение водорослей по профилю почвы в ту или иную сторону.
Сходные данные о быстром снижении количества водорослей по профилю лесных почв приведены в ряде отечественных [сводка: Штина, Голлербах, 1976а; Чаплыгина, 1977] и зарубежных публикаций. Гиндак [Hindak, 1974], определявший биомассу водорослей в почве по содержанию хлорофилла а, показал резкое падение этого показателя в пределах первого сантиметра пойменной почвы под лесом. Численность клеток водорослей в тенистом лесу Индии снизилась от 24 тыс. в 1 г почвы в слое 0 - 0,5 см до 9,6 тыс. в слое 2 - 5 см и до 6,2 тыс. на глубине 10 - 15 см; в освещенном лесу соответственно показатели снижались от 57,3 тыс. до 36,3 и 17,8 тыс. в 1 г почвы.
Таким образом, изменение количества водорослей и числа их видов по профилю почвы носит довольно сложный характер. Особенно неравномерно распределяются водоросли в самом верхнем горизонте почвы, где часто приходится сталкиваться с микростратификацией водорослей. В связи с этим альгологическому анализу желательно подвергать не средний образец из всего рекомендуемого для изучения 15-сантиметрового слоя почвы, а целесообразно выполнять послойное (через каждые 5 см) определение состава и количества водорослей. Значительно заниженные данные, полученные Т. С. Носковой [1972] при определении количества водорослей во всем слое почвы толщиной 0 - 10 см по сравнению с результатами анализа отдельных составляющих его слоев 0 - 3 и 3 - 10 см, еще раз свидетельствуют в пользу послойного определения состава водорослей в лесных почвах в пределах верхнего горизонта.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'