Реликтовые элементы
Современный флористический состав растительного покрова того или иного региона представляет собой результат длительного исторического развития флоры. Реликтовые виды, сохранившиеся с древнейших времен геологической истории, - свидетели тех многообразных преобразований физико-географической обстановки, которые претерпела та или иная территория до настоящего времени. Как отмечал Е. В. Вульф (1941), реликты служат теми вехами, руководствуясь которыми мы воссоздаем историю развития растительного покрова Земли. Вслед за Е. В. Вульфом (1941) под реликтовым видом мы понимаем остаток более или менее древней флоры, имеющий реликтовый ареал, занимаемый им с момента вхождения в состав флоры. Реликтовый вид является как бы отражением процесса исторического развития флоры. Возраст реликта определяется временем вхождения вида в состав флоры. Под реликтовым понимается остаток некогда более обширного ареала, формирование которого происходило при других, ранее существующих условиях. В таком понимании термин "реликтовый вид" является понятием историко-географическим, связанным с историей расселения вида.
Л. В. Бардунов (19736), характеризуя флорогенетические особенности моховых флор, подчеркивал, что малые размеры тела мхов позволяют им поселяться в подходящих по условиям экологических микронишах и переживать в них значительные макроклиматические изменения. Медленное течение эволюционного процесса обусловили обширные ареалы у мхов, древний возраст большинства видов и слабую дифференциацию в случае разобщенных ареалов. При этом древний возраст мхов и их способность переживать крупные физико-географические перестройки приводят к тому, что бриофлора любого региона всегда оказывается более древней по сравнению с флорой цветковых растений, причем содержит достаточно большую реликтовую группу, и реликтовые виды распространены более широко по сравнению с таковыми высших растений. Подобные особенности характерны и для лишайниковых флор.
Широкоизвестная адаптивная способность лишайников, малые размеры их талломов позволяют им значительно лучше по сравнению с цветковыми растениями выживать в неблагоприятных для них условиях. Вследствие большей ригидности генотипической структуры, как отмечал А. Н. Окснер (1948), лишайники в значительно меньшем количестве и более медленно образуют новые биотипы по сравнению с цветковыми растениями. Это является причиной, с одной стороны, затяжного процесса эволюции, с другой - древнего возраста большинства видов лишайников. В то же время их адаптивные возможности позволяют им иррадиировать далеко за пределы их первоначального ареала, что способствует формированию обширных ареалов, характерных для большинства видов лишайников, а также выживать в неблагоприятных условиях далеко от основной части ареала. Поэтому лишайниковая флора того или иного района имеет в своем составе значительное число реликтовых элементов.
Е. П. Коровин (1961) отмечал, что следы древнего третичного расселения флоры правильнее всего искать в ареалах современных родов или по крайней мере их подразделений, ибо чрезвычайно редки случаи среди цветковых растений, когда вид без изменений переживает длительный этап в течение всего третичного времени. В отличие от цветковых растений многие виды лишайников характеризуются обширными дизъюнктивными ареалами. Широкие дизъюнкции их ареалов нередко не находят объяснений в современных природных условиях и, видимо, обусловлены историческими причинами.
Так, Н. А. Миняев (1940, с. 415) писал: "Примитивный образ жизни и связанный с этим консерватизм в процессе эволюционного развития, характерный, видимо, для всех низкоорганизованных растительных организмов, в том числе для мхов и лишайников, способствовал не только тому, что отдельные их виды сумели до настоящего времени сохранить в почти неизменном виде сбои прежние, подчас очень древние, раннетретичные ареалы, но также и тому, что в пределах любой современной конкретной, т. е. иначе - территориальной, флоры мы находим значительно больше реликтовых элементов среди представителей этих групп, чем среди высших растений". Далее он отмечал, что по ним, по этим "живым ископаемым", мы можем в значительной мере воспроизвести те связи во флорах, которые уже потеряны высшими растениями и которые, к сожалению, мы никогда не восстановим документально с помощью палеонтологического метода.
Не ставя целью данной работы анализ всех реликтовых видов лишайников во флоре Монголии, остановимся лишь на отдельных представителях выделенных нами реликтовых групп. Эти группы объединяют так называемые климатические реликты (Окснер, 1974), представляющие собой следы флоры, существовавшей при других климатических отношениях. Их климатические потребности находятся в некотором противоречии с современными климатическими условиями.
В лихенофлоре Монголии можно различить реликтовые элементы флор разного возраста: 1) третичного периода, 2) плейстоцена, 3) голоцена.
Реликтовые элементы флор третичного периода. А. А. Юнатов (1954), характеризуя растительность территории Монголии третичного периода, отмечал, что, судя по косвенным данным, поскольку прямых палеоботанических свидетельств у нас нет, можно полагать появление в этой стране степных и пустынно-степных ландшафтов по меньшей мере с миоцена. В то же время, как указывал А. А. Юнатов, на юге страны могли существовать и участки пустынь, а в горных районах севера - леса тургайского типа и их дериваты. Похолодание климата в конце плиоцена-плейстоцене, резко выраженные процессы орогенеза на рубеже третичного и четвертичного периодов сильно отразились на распределении растительного покрова и привели к разрыву ареалов ряда видов растений, в том числе и лишайников, а также к распаду и исчезновению термофильных элементов тургайской флоры. Однако флоры, господствовавшие в третичный период, сохранили некоторые реликтовые элементы в составе современной лихенофлоры Монголии.
В числе реликтовых элементов третичных флор среди лишайников можно выделить группу пустынно-степных реликтов. К наиболее древним реликтовым элементам в лихенофлоре Монголии можно отнести так называемые гондванские элементы (в смысле М. Г. Попова (1931)), проявляющие генетические связи с южными ксерофитно-субтропическими флорами. К таким реликтовым типам могут быть отнесены некоторые виды желтоокрашенных акароспор, например Acarospora sulphurata Arnold (рис. 33).
В разделе, посвященном географическим элементам лихенофлоры Монголии, было показано, что Acarospora sulphurata Arnold характеризуется обширным ареалом, с крупными дизъюнкциями на Африканском и Евразиатском континентах. Как можно было видеть, изолированные участки ареала этого вида приурочены к пустынным районам Юго-Западной Африки, к пустынным и прибрежным районам Северной Африки, к средиземноморским районам Европы и территории Восточного Средиземья. После обширного разрыва на территории Туранской и Туркестанской провинций этот лишайник появляется на территории Монголии, где изолированное местообитание его известно в районе Котловины Больших Озер (Schubert, Klement, 1971). Прослеживающиеся географические и генетические связи этого вида с южными ксерофитно-субтропическими флорами и его изолированное местообитание на территории МНР позволяют причислить его к древним реликтовым типам, к "гондванскому" элементу (в смысле М. Г. Попова), или, точнее, к гондвано-средиземноморскому (Невский, 1937). О наличии пустынных реликтов в составе флоры сосудистых растений этой страны писал А. А. Юнатов (1950, 1954), отмечая, что наиболее древними (палеогеновыми) являются те элементы, которые пришли в Монголию с запада, тотчас после того как здесь сложились пустынные условия, причем их родиной является область древних африканских пустынь (флора вельвичии М. Г. Попова) .
Группа желтоокрашенных акароспор подрода Xanthothallia Н. Magn. насчитывает 77 видов, основное распространение которых, как отмечал Р. Сантессон (Santesson, 1944), связано с областями, характеризующимися жарким и засушливым климатом. Лишь немногие из них проникают далеко на север. Так, в Арктике известен лишь один арктоальпийский вид из этой группы лишайников - A. chlorophana (Wahlenb.) Massal. В Африке отмечены 25 видов, причем 18 из них являются африканскими эндемиками (Голубкова, 1980). Наиболее богаты эндемичными видами желтоокрашенных акароспор пустынные районы Южной и Юго-Западной Африки - пустыни Калахари, Намиб, Карро. Вторым очагом видового разнообразия этой группы лишайников являются горнопустынные районы юго-запада Северной Америки и Мексики, где известно также 25 видов желтоокрашенных акароспор, из них 14 - эндемики флоры этих районов. Богато представлена эта группа лишайников также в районе Средиземноморья, где встречается 16 видов, из них 6 - эндемики этого региона. В Азии по направлению с запада на восток происходит постепенное убывание видового состава: в Монголии встречается 9 видов, в Средней Азии - 5, в Южной Сибири - 4, на Дальнем Востоке - 2. Интересно, что очаг видового разнообразия подрода Xanthothallia Н. Magn. рода Acarospora, связанный с районами Южной и Юго-Западной Африки, совпадает с центром развития древнейшей ксерофильной флоры, о котором писал М. Г. Попов (1927). М. Г. Попов указывал, что именно в пустынях Карро и Юго-Западной Африке вероятнее всего в эпоху мела или, может быть, юры происходило формирование первичной пустынной флоры, которую он именует флорой вельвичии. Эта флора, по мнению М. Г. Попова, имела кардинальное значение для истории развития пустынной флоры Голарктики. О древнем африканском жарком аридном центре формирования некоторых ксерофильных групп лишайников упоминали А. Н. Окснер и Н. А. Миняев (1940). А. Н. Окснер указывал также на два других центра формирования древней ксерофильной пустынной флоры - это район Средиземья и районы Центральной и Юго-Западной Америки. Как полагал М. Г. Попов (1927), некоторые типы древней ксерофильной флоры вельвичии сохранились и до наших дней. Характерными особенностями их ареалов являются следующие: 1) пустынные типы, явно имеющие корни в Южной Африке, во флоре вельвичии, располагают в северном полушарии центрами своего развития в Северной и Северо-Восточной Африке, а также в Юго-Западной Азии; 2) типичной чертой всех этих групп является то, что они имеют изолированных представителей в юго-западных частях Северной Америки; 3) в Старом Свете они подходят вплотную к Атлантическому океану, и, наоборот, к востоку они более или менее быстро выклиниваются. Как можно видеть, эти особенности построения ареалов типов древней ксерофильной флоры вельвичии совпадают в общих чертах с ареалом подрода Xanthothallia Н. Magn. Распространение этой группы лишайников иллюстрирует ареал Acarospora schleicheri (Ach.) Massal. (рис. 34). Как отмечал М. Г. Попов (1927), такое построение ареала обусловлено в первую очередь современными причинами, но вряд ли только одними ими. Имеется много оснований предполагать, что и исторические причины сыграли в этом значительную роль. Обильное развитие и формирование типов флоры вельвичии в районах Голарктики, по мнению М. Г. Попова, несомненно нужно связывать с позднейшей геологической эпохой - с концом третичного периода и современным, но возникновение их относится безусловно к очень древней эпохе, к началу тертиэра.
К числу древних, донеогеновых, реликтовых типов, видимо, можно отнести также формы, проявляющие генетические связи с сонорскими пустынными флорами Северной Америки (Acarospora peltastica Zahlbr., A. nevadensis Н. Magn.).
Эпилитные ксерофитные пустынные виды Acarospora peltastica Zahlbr. и A. nevadensis Н. Magn. характеризуются дизъюнктивными ареалами, одна часть которых находится на территории Центральной Азии, другая - в юго-западных районах Северной Америки.
Эти виды относятся к группе коричневоокрашенных акароспор, к подроду Acarospora. Древнесредиземноморское подцарство Голарктики является одним из центров, концентрирующих виды этой систематической группы лишайников. В этом районе земного шара встречаются 154 вида из этой группы лишайников, из них 44 вида - эндемики района. Эндемиками Средиземноморской области являются 23 вида, эндемиками Центральной Азии - 15 и 6 видов эндемичны для Ирано-Турана. Во флоре Мадреанского (Сонорского) подцарства Голарктики встречается 50 видов коричнево- окрашенных акароспор, из них 29 видов - ее эндемики. Таким образом, район Древнего Средиземья (в понимании М. П. Попова (1927, 1931, 1940)) является одним из центров видового разнообразия рода. По-видимому, район Древнего Средиземья был одним из центров видообразования этой группы лишайников, о чем свидетельствует не только концентрация видов, в том числе эндемичных, в этих районах земного шара, но также значительное разнообразие морфотипов.
Такие центры развития, как европейско-бореальный, видимо, следует считать вторичными для этой группы лишайников. Анализируя систематический состав эндемичных видов европейского бореального центра, можно сделать вывод, что среди них преобладают представители серии Aspicilioides Н. Magn. секции Acarospora. Эта группа, самая многочисленная (135 видов) среди коричневоокрашенных акароспор, объединяет наиболее типичные для рода Acarospora морфогенетические признаки - однообразное по строению слоевище в виде чешуйчато-ареолированных корочек, многочисленные апотеции, погруженные в слоевище, мелкие, многочисленные в сумках (до 200 - 400) эллипсоидные споры. Виды этой серии близки между собой систематически и слабо отграничены друг от друга анатомо-морфологическими признаками (толщиной парафиз, высотой гимениального слоя, формой ареол или чешуек, числом апотециев в каждой из них и некоторыми другими признаками), Так, из 29 эндемиков горных районов Средней Европы 20 принадлежат к этой серии видов. В европейской флоре Циркумбореальной области в настоящее время резче всего выражен тип Acarospora. Как подчеркивал В. Л. Комаров (1908, с. 148), "обилие близких видов указывает лишь на распыление в данной области родового и секционного типа применительно к отдельным географическим или топографическим ареалам, разобщенным между собой, как это обычно имеет место в горных долинах." И далее он отмечал, что такое распыление морфологического типа соответствует текущему процессу видообразования, но не древнему центру.
Acarospora peltastica Zahlbr. входит в число видов подсерии Inactivae Н. Magn., в группу Subglobosae Н. Magn., характеризующуюся в отличие от других многочисленных представителей этой подсерии округлой формой спор. В составе группы 10 видов, 8 из них распространены в пределах территории Древнего Средиземья (в понимании М. П. Попова (1927)). Один вид - A. plinthina Н. Magn. - является эндемиком горных районов Средней Европы (Венгрии); A. ferdinandii Hue - эндемик австралийской флоры, что свидетельствует о древнем возрасте этой группы в целом и о ее гондванских генетических связях. Остальные виды распределяются на земном шаре следующим образом: A. trachytica Jatta - эндемик Средиземноморской области; ареал 2 видов - A. elbursensis Н. Magn. и A noduiosa (Duf.) Hue - связан с Ирано-Туранской областью; 2 вида - A. coloradiana Н. Magn. и A epilutescens Zahlbr. - являются эндемиками Сонорского подцарства Голарктики; один вид A. superans Н. Magn. - центральноазиатский эндемик; один вид - A. reagens Zahlbr. - сонорско-древнесредиземноморский и один вид - A. peltastica Zahlbr. - сонорско-центральноазиатский вид. Ареал последнего связан с пустынными районами юго-запада Северной Америки (Калифорнией и Аризоной), где он обитает на гранитных скалах и почве (Magnusson, 1929). Вторая часть ареала этого вида простирается в южных гобийских районах Монголии, где он растет на скалах в горных полынно-злаковых степях хр. Адж-Богдо. A. nevadensis Н. Magn. относится к другой подсерии - Reagentes Н. Magn. серии Aspicilioides Н. Magn., представителям которой свойственно покраснение слоевища при воздействии хлорной известью, к группе видов Globosae Н. Magn., также характеризующихся округлой формой спор. В составе группы 5 видов, распространение которых также связано в основном с территорией Древнего Средиземья: A. sphae- rospora Н. Magn. - европейский вид, встречающийся в Южной Франции и Австрии; A. bornmuelleri Steiner - синдско-анатолийско-гобийский вид; A. fuscohepatica (Nyl.) Hue - среднеазиатско-монгольский; A. jenisejensis Н. Magn. - южносибирско-монгольский; A. nevadensis Н. Magn. - сонорско-древнесреди-земноморский вид. A. nevadensis Н. Magn. имеет обширный дизъюнктивный ареал, на территории Азии охватывающий горы Памиро-Алая (Голубкова, 1972, 1973а) и в пределах Монголии южные районы Гобийского Алтая (Голубкова, 1981). Третий, изолированный, участок ареала этого вида находится в Юго-Западной Америке, в горных районах штата Невада (Magnusson, 1929). Дизъюнктивные ареалы A. peltastica Zahlbr. и A. nevadensis Н. Magn., характеризующиеся тихоокеанским разъединением, генетические связи этих видов с флорами территории Древнего Средиземья позволяют отнести их к формам, о которых Е. П. Коровин (1961) писал, что подобные формы являются древними остатками видообразования в зоне Тетиса до разъединения Старого и Нового Света. Как отмечал Е. П. Коровин, они могли зародиться в верхнемеловую эпоху в пустынях Древнего Средиземья.
Рассматривая древние связи пустынных флор Евразии и Северной Америки, М. М. Ильин (1945, с. 31) писал: "некогда, именно в период связи Американского континента и континентов Старого Света, в области "древнего разлома земли", или истинного Средиземноморья, которое, очевидно, следует датировать верхним мелом, была развита в этой части Земли своеобразная флористическая область, в которой были достаточно широко распространены ксерофитно-пустынные территории. Несмотря на широкое распространение этой области от Калифорнии до Центральной Азии, можно установить общность развития этой флоры в меловой период и обмен представителями между крайними районами этой флоры".
Древние географические связи флоры Монголии с пустынными флорами Северной Америки фиксируют также представители сем. Heppiaceae. Как отмечал монограф этой группы лишайников американский ученый Ветмор (Wetmore, 1970), виды сем. Heppiaceae часто встречаются в аридных и семиаридных областях земного шара, где обитают на почве и скалах. В частности, лишайники этого семейства, по свидетельству Ветмора, достигают массового развития в пустынных районах юго-запада Северной Америки, где некоторые из них являются доминантами лишайниковых группировок в составе пустынных растительных сообществ. Представители сем. Heppiaceae относятся к числу характерных лишайников лихенофлоры гобийских пустынно-степных районов Монголии, они обычны также в горных степях и других районах этой страны. В то же время следует отметить, что в пустынно-степных районах Казахстана и Средней Азии лишайники этого семейства неизвестны, за исключением Happia lutosa (Ach.) Wetm. и центрально-азиатского вида Peltula impressula (Н. Magn.) Golubk., приводимых для Центрального Тянь-Шаня (Инашвили, 1975; Бредкина, 1979, 1981а). Но вероятнее всего в этом районе проходит юго-западная граница их центральноазиатского ареала. Рассмотрим ареалы некоторых видов сем. Heppiaceae.
Основной ареал Peltula obscurans (Nyl.) Gyeln. охватывает пустынные районы юго-запада США, где в некоторых районах этот вид развивается обильно и входит в число доминантов лишайниковых группировок в пустынных растительных сообществах (Wetmore, 1970). Южнее он проникает в Мексику и Южную Америку (Перу, Бразилия). Этот лишайник встречается также в Африке - в Эфиопии, на островах Зеленого Мыса и на Канарских островах (Swinscow, Krog, 1979). На территории Евразии он известен только во Франции и Италии (Wetmore, 1970) и далее, после огромной дизъюнкции, появляется в Азии, в степных районах Монголии. В Монголии этот вид является редким, он был найден в горных степях южной части Хэнтэя (Schubert е. а., 1969) и в Восточной Гоби, в горах Баян-Мунх, где обитал в луково-ковыльковых степях на камнях и скалах в хорошо освещенных местах (Голубкова, 1981). Эти изолированные местонахождения P. obscurans (Nyl.) Gyeln. на территории МНР, по-видимому, реликтовые.
В составе группы пустынно-степных реликтов - некоторые виды рода Aspicilia - представители так называемой группы кочующих аспицилий - Aspicilia esculenta (Pall.) Flag., A. lacunosa Mereschk., A. vagans Oxn. Распространение этих лишайников связано в основном со степями, пустынями, полупустынями Турана, где они обитают на глинистой и солонцеватой, нередко каменистой почве. На территории МНР, в ее западной части, главным образом в Котловине Больших Озер, имеются самые восточные, изолированные от основной части ареала, местообитания этих видов на Евразиатском континенте.
Так, ареал Aspicilia esculenta (Pall.) Flag, простирается от районов Юго-Восточной Европы - Греции, Румынии (Moruzi е. а., 1967), Крыма, Саратовской, Волгоградской, Астраханской областей, захватывая Закавказье (Армения) (Абрамян, 1980) и далее, к востоку - Арало-Каспийскую котловину, плато Устюрт, Туркмению (Липский, 1905; Бредкина, Макарова, 1971; Джураева, 1973, 1978), достигая на юге территории Ирана, Карагандинской и Акмолинской областей. Восточная граница основной части ареала этого вида проходит в Восточно-Казахстанской области, в котловине оз. Зайсан, и южнее - в Тянь-Шане, в Иссык-Кульской котловине (Андреева, 1959). Этот лишайник известен также в Северной Африке - Алжир (Flagey, 1896) и недавно впервые был указан для района Великих равнин Северной Америки. На территории Монголии имеется лишь одно местонахождение этого вида в Котловине Больших Озер, в окрестностях оз. Убсу-Нур (Klement, 1965). Здесь он обитает на солончаке на северо-восточном берегу оз. Убсу-Нур, на высоте 750 м над ур.м. Можно полагать, что это изолированное северо-восточное местообитание A. esculenta (Pall.) Flag., возможно, реликтовое и служит свидетельством более обширного ареала вида в прошлом, по-видимому В миоцене-плиоцене.
Ареал другого вида из группы кочующих аспицилий - Aspicilia lacunosa Mereschk. связан главным образом с Туранской, или Арало-Каспийской, провинцией (Тахтаджян, 1978); он охватывает пустыню Бетпак-Дала (Андреева, 1963, 1965), Прибалхашье (Голубкова, 1980), достигая на севере Карагандинской области (Окснер, 19396), на юге - Туркмении (Джураева, 1978), Ферганской долины, Алайского хребта (Шафеев, 19536) и степей Киргизии (Мережковский, 1911). Восточная граница этой основной части ареала вида также проходит в районе оз. Зайсан. На территории Монголии, в южной части Котловины Больших Озер, в предгорьях Монгольского Алтая, имеется изолированное местообитание этого лишайника (Klement, 1965), где он обитает на почве в полупустыне.
Ареал Aspicilia vagans Oxn. также связан в основном с каменистыми степями, полупустынями и пустынями Турана, где он встречается от низовьев Волги, Восточного Закавказья, на юге достигая Туркмении и Ирана, на севере Южного Урала, на востоке Киргизии (Окснер, 1971). Изолированное местонахождение этого лишайника имеется в Чуйской степи, на Алтае. На территории Монголии A. vagans Oxn. встречается в пустыне Джунгарская Гоби, где, по-видимому, проходит восточная граница основной части ареала этого вида. После некоторой дизъюнкции, падающей на Монгольский Алтай, этот лишайник появляется в Долине Озер, где изолированное местообитание его было обнаружено восточнее сомона Юсун-Булак, в полупустыне на высоте 1600 - 1700 м над ур. м. (Klement, 1965). Возможно, что до поднятия Монгольского Алтая этот пустынно-степной вид имел в юго-западной части Монголии сплошной ареал.
Ареал группы кочующих аспицилий охватывает пустынно-степные районы Северной Африки, Юго-Восточной Европы, Казахстана, Средней Азии, достигая на востоке Центральной Азии, а на юге территории Ирана, а также район прерий Северной Америки. Этот обширный ареал свидетельствует о древнем происхождении этой группы в целом и ее генезисе в пределах территории Древнего Средиземья. Современным центром видового многообразия этой группы лишайников являются пустыни и полупустыни Туранской провинции Западноазиатской подобласти Древнесредиземноморского подцарства (Тахтаджян, 1974, 1978). Из 8 видов этой группы в Туранской провинции встречается 7 - Aspicilia esculenta (Pall.) Flag., A. emiliae Tomin, A. vagans Oxn., A. fruticulosa (Eversm.) Flag, A. aspera (Mereschk.) Tomin, A. hispida Mereschk., A. lacunosa Mereschk. Один вид - A. tominii Oxn. - эндемик Чуйской степи. В Северной Африке встречаются 2 вида - A. esculenta (Pall.) Flag, и A. fruticulosa (Eversm.) Flag.; 2 вида - A. esculenta (Pall.) Flag, и A. hispida Mereschk. - известны в Северной Америке. А. А. Еленкин (1901а) в работе, посвященной кочующим лишайникам пустынь и степей, показал генетические связи этой группы лишайников с некоторыми пустынно-степными прикрепленными формами аспицилий, в частности с ирано-туранским видом Aspicilia desertorum (Kremplh.) Mereschk., и тем самым выявил связи этой группы со среднеазиатским флористическим центром.
Как мы полагаем, господствовавшая на рубеже миоцена - плиоцена обстановка на материке Евразии могла способствовать расширению географических ареалов этих среднеазиатских видов до западных районов Центральной Азии. По данным недавних исследований (Hsu, 1972; Монин, Шишков, 1979), примерно на границе между миоценом и плиоценом на месте Средиземного моря находилась жаркая пустыня с солеными озерами, лежащими ниже уровня океана. Этот пустынный центр не мог не оказать влияния на климат Евразии. На территории Европы в это время господствовал сухой и теплый климат и широкое распространение получили степные ландшафты (Монин, Шишков, 1979). Е. М. Лавренко (19546) на основании палеозоологических и палинологических данных датировал возникновение степей на юге европейской части СССР миоценом, при этом отмечая, что на протяжении верхнего сармата, меотиса и плиоцена полоса открытых пространств продолжала расширяться. С миоценом связано формирование степных ландшафтов Центральной Азии (Девяткин, Шилова, 1970). А. А. Юнатов (1954) также писал, что появление в Монголии степных и пустынно-степных ландшафтов можно полагать по меньшей мере с миоцена. До альпийской фазы орогенеза, когда произошла значительная перестройка почти всей поверхности Монголии и сформировался современный высокогорный и средне- горный рельеф Монгольского Алтая, Хангая и Хэнтэя, западная часть Монголии была территорией, благоприятной для миграционного потока растений с юго-запада, который и оставил в этом районе своих мигрантов (Соболевская, 1958). Как можно было видеть, основное распространение представителей кочующих аспицилий на территории Монголии связано с районом Котловины Больших Озер. Как известно, Котловина Больших Озер является своеобразной ареной проникновения на территорию МНР целого комплекса туранских, казахстано-северотуранских, среднеазиатских и других видов цветковых растений (Юнатов, 1950, 1974; Грубов, Юнатов, 1952; Грубов, 1959, 1963; Карамышева, Банзрагч, 1976, и др.). Можно полагать, что особенно интенсивные взаимосвязи между среднеазиатским и центральноазиатским флористическими центрами существовали до формирования современного рельефа Северо-Западной Монголии до третично-четвертичного горообразования. Процессы орогенеза привели к разрыву ареалов многих из этих видов, в том числе и выше упомянутых аспицилий. В то же время возникновение молодых цепей Алтая, Сайлюгема и Танну-Ола, отделивших высоким барьером замкнутые котловины этой части Монголии от северо-западных воздушных течений, способствовало ксерофитизации этих районов Монголии (Мурзаев, 1952). Современный пустынный климатический режим, господствующий в Котловине Больших Озер, по-видимому, способствует консервации среднеазиатских пустынно-степных видов, имеющих в этом районе свои реликтовые местообитания.
В группу пустынно-степных реликтов, видимо, можно включить и некоторые другие виды аридного элемента, имеющие на территории Монголии изолированные местообитания, причем дефект их ареала на Евразиатском континенте позволяет предполагать, что этот разрыв мог произойти в результате третично-четвертичного орогенеза. Среди них некоторые пустынно-степные виды родов Acarospora.
Аридный эпилитный лишайник Acarospora bornmuelleri Anzi характеризуется дизъюнктивным синдско-анатолийско-гобийским ареалом. Одна часть ареала этого вида связана с Синайским полуостровом, территорией Ирака и Анатолией (Турция) (Magnusson, 1929; Galun, 1970). После значительной дизъюнкции, падающей на Иранское и Среднеазиатское нагорья, этот вид появляется в гобийских районах Монголии (Голубкова, 1981). Здесь он рассеянно встречается в низкогорных хребтах в гобийских пустынях, в Гобийском Алтае, в хр. Адж-Богдо, достигая на востоке южных степей Восточной Монголии (Дариганга). Таким образом, огромное разъединение в ареале этого вида может быть объяснено орогенезом в пределах Средиземноморско-Гималайской геосинклинали. Как известно, наибольшей интенсивности горообразование достигло в основном в конце неогена - плейстоцене. Ранее, возможно, этот вид имел сплошной ареал.
Как было отмечено выше, в третичный период на территории современной Монголии, в северной части страны, наряду с ксерофитной флорой располагалась лесная мезофильная флора тургайского типа (Юнатов, 1954). Е. М. Лавренко (1930, с. 351) писал, что "факт существования в третичное время (плиоцен) в пределах современной умеренной зоны северного полушария циркумполярной зоны широколиственных лесов является одним из твердо установленных в генетической географии растений". Однако, как известно, в дальнейшем, в связи с похолоданием, наступившем в плиоцене, и последующим горообразованием и плейстоценовыми оледенениями эта зона распалась на ряд изолированных участков. Как указывал А. Н. Окснер (1946а), мезофильная листопадная флора нашла приют как в приатлантической полосе Западной Европы, так и под защитой горных систем в рефугиальной зоне, от Пиренеев до Карпат, и далее - в Южном Крыму, на Кавказе, Южном Урале, Алтае, на Дальнем Востоке и местами в восточной части Северной Америки. Он писал также, что отдельные представители этой флоры сохранились в окрестности Красноярска, Минусинска, Саянах, а кроме того, как показывают лихенологические данные, в Прибайкалье и южном Забайкалье, где им же были собраны такие лишайники, как Lobaria retigera (Bory) Trevis., Cetraria laureri Krempelh., Phaeophyscia endococcina (Koerb.) Moberg, Heterodermia speciosa (Wulf.) Trevis., которые он считал реликтами термофильной неморальной флоры в современной флоре этих районов. Районы Прибайкалья и южного Забайкалья отнесены А. Н. Окснером (1946) к числу рефугиумов термофильной неморальной флоры. В этих районах известны также среди сосудистых растений многочисленные реликты широколиственных лесов. Они наиболее широко распространены на юго-восточном побережье Байкала и прилегающих к нему северных склонах хр. Хамар-Дабан, в меньшем количестве встречаются на всех побережьях Байкала, на склонах Приморского хребта, в Восточном Саяне, в районе Иркутска и в других местах (Малышев, 1957, 1960, 1961 и др.). В этих же районах были найдены неморальные виды мхов (Бардунов, 1963). Многие из них Л. В. Бардунов относил к реликтам третичного периода. Л. В. Бардунов на основании своих данных и данных других исследователей (Лавренко, 1938; Ильин, 1941; Бардунов, 1963 и др.) приходит к выводу, что Алтае-Саянская область является важным рефугиумом третичных реликтов. Следует указать также на работу Н. В. Седельниковой (1977а), в которой автор приводит для флоры Алтая (Горная Шория) 18 видов лишайников - реликтов неморальной флоры, возраст которых она датирует плиоценом.
Как известно, относительно возраста реликтов неморальной флоры в составе таежной флоры Южной Сибири, а также во флоре Южного Урала и лесостепных и степных районов европейской части СССР до сих пор нет единого мнения ботаников, и этот вопрос остается дискуссионным. Большинство исследователей относит к третичному возрасту термофильную реликтовую флору широколиственных лесов Русской равнины, Южного Урала, Западной, Средней и Восточной Сибири (Крылов, 1891; Коржинский, 1894; Ильин, 1940, 1941; Клеопов, 1941; Попов, 1949, 1953; Гричук, 1949, и др.). Ю. Д. Клеопов (1938, 1941) датировал термофильные лесные реликты лесостепи и степи Русской равнины межледниковым рисс-вюрмским временем. Эту точку зрения впоследствии разделяли многие ботаники. В. В. Ревердатто (1940) высказал мнение о послеледниковой иммиграции реликтов широколиственных лесов на территорию Средней Сибири. В дальнейшем эта гипотеза получила развитие в одной из работ Г. Э. Гроссета (1962).
В отношении лишайников этот вопрос, видимо, не может быть решен однозначно. Одни реликтовые типы современных листопадных лесов во флоре Монголии, возможно, являются третичными реликтами, другие - голоценовыми. В формировании лесной мезофильной лихенофлоры Монголии, вероятно, значительна роль тургайского элемента. Многие представители автохтонных тургайских родов лишайников (Evernia, Bryoria, Cetraria и др.) в связи с наступившим в конце плиоцена - плейстоцене похолоданием подверглись сильному влиянию процесса бореализации и были ассимилированы горно-таежной флорой. Однако в составе тургайской флоры могли присутствовать и некоторые тропогенные формы - элементы субтропических мезофильных флор. Наиболее термофильные из них, по-видимому, погибли под влиянием холодного и сухого климата плейстоцена. Но более микротермные формы, особенно выходцы из горных районов тропических областей, могли сохраниться в современной флоре Монголии в качестве реликтов, например Heterodermia speciosa (Wulf.) Trevis., H. leucomela (L.) Poelt, Leptogium hildenbrandii (Garov.) Nyl., L. burnetiae Dodge, Pannaria conoplea (Ach.) Bory. В разделе, посвященном географическим элементам лихенофлоры, подробно обсуждались ареалы этих видов лишайников. При этом было показано, что эти виды характеризуются обширными ареалами, прерываемыми огромными дизъюнкциями, приуроченностью изолированных участков ареалов к рефугиумам третичной неморальной флоры как Евразии, так и Северной Америки, изолированным систематическим положением в составе флоры Монголии и связью с определенными местообитаниями, которым присущи особые микроклиматические условия с повышенной влажностью. Важную роль в сохранении этих видов начиная с третичного времени в современной флоре Монголии играли их высокие адаптивные способности (Окснер, 1946а; Седельникова, 1977а).
Реликтовые элементы флор плейстоцена (перигляциально-степная и гляциальная группы реликтов). Картина формирования флоры и растительного покрова в плейстоцене как для Урала, так и в целом для Сибири была освещена на основе глубокого флористического анализа в интересных работах И. М. Крашенинникова (1954а, 1954б). Им же была разработана концепция "плейстоценового флористического комплекса". Как полагал И. М. Крашенинников, во время оледенений плейстоцена в средних широтах Евразии существовала перигляциальная лесостепь, где степи чередовались с сосновыми и лиственничными лесами. Периоды усиления континентальности климата в плейстоцене, вызвавшие деградацию или полное исчезновение широколиственных пород, способствовали развитию светлохвойных и мелколиственных насаждений. Вместе с ними, особенно при возрастании сухости климата, получила широкое распространение своеобразная бореально-ксерофитная растительность, состоящая из древних (прастепных) элементов, которые принадлежали в своих исходных типах, вероятно, к альпийской, субальпийской и горно-ксерофитной ветви, отчасти доледниковой растительности (Крашенинников, 1954б). При этом в особо сухие и холодные климатические периоды группировки древних бореально-степных элементов, первоначально возникнув в горных областях Северной и отчасти Центральной Азии, позже, в эпохи, синхроничные последним оледенениям Европы, проникли туда, сочетаясь вместе с сосновыми, лиственничными и березовыми лесами, частью вдоль перигляциальной зоны, частью в некотором отдалении от нее, в экстрагляциальном поясе (Крашенинников, 1954а, 1954б). Таким образом, "плейстоценовый флористический комплекс" сочетает экологически разнородные флористические элементы, у которых центром ареала, более обширного в прошлом и более ограниченного в настоящем, с рядом дизъюнкций, являются преимущественно горные области Южной Сибири и Северной Монголии, особенно районы Байкало-Саяно-Хангае-Алтайского сооружения.
Е. М. Лавренко (1981) в статье о развитии перигляциальной лесостепи отмечал, что точку зрения И. М. Крашенинникова в основном следует признать верной, но с уточнением, что не во все фазы оледенений это была лесостепь, часто это были почти безлесные степи. Как показывают палеозоологические и палеопалиноло- гические данные, во время ледниковых фаз плейстоцена, преимущественно последнего вюрмского (валдайского) оледенения, в условиях резко континентального, холодного и сухого климата обширные пространства в средних широтах Евразии занимали перигляциальные тундры, а южнее - перигляциальные степи (Юрцев, 1968, 1974, 1981; Величко, 1973; Лавренко, 1981). При этом на обширных перигляциальных пространствах соприкасались и взаимопроникали биомы не только тундр и холодных степей, но и высокогорная растительность, спустившаяся с обледеневших гор.
Северная часть Монголии, как и Восточная Сибирь, относится к районам, где до настоящего времени сохранилась многолетняя мерзлота, и именно на этой территории в лесной зоне и даже в Арктике (к востоку от Лены) осталось множество реликтовых степных, луговых, травяно- и кустарничково-тундровых позднеплейстоценовых (перигляциальных) фитоценозов (Лавренко, 1981). Как степная, так и лесная растительность МНР в наибольшей мере, чем где-либо в Северной Евразии, сохранила черты комплекса холодной плейстоценовой лесостепи. Как показало изучение лесов МНР (Леса Монгольской Народной Республики, 1978), только для Монголии, например, характерен высотно-поясной комплекс псевдотаежных (сухомшистых) типов лиственничных лесов, широко представленный здесь в центральном Хангае и Монгольском Алтае. О былом распространении таких лесов в других районах Монголии и в горах Южной Сибири свидетельствуют реликтовые признаки плейстоценового комплекса, обнаруживаемые как в подтаежных лиственничниках юго-восточного и северо-восточного Хангая, кедровниках ритидиевых южного Хэнтэя, так и за пределами Монголии, в ритидиевых лиственничниках Прибайкалья и Алтая. Эти реликтовые черты в лесах Монголии, Забайкалья и других районов Южной Сибири проявляются в широком распространении "сниженных альпийцев". В таких районах, как Северная Монголия, в составе "плейстоценового флористического комплекса" в значительной степени представлены элементы, эволюция которых проходила в условиях криоксерической обстановки высоких нагорий Центральной Азии. В формировании этого комплекса большая роль принадлежала не только ксерофилизованным потомкам бореальной флоры, но и криоксерофилизованным дериватам степной и пустынно-степной центральноазиатской флоры (Грубов, Юнатов, 1952).
К видам, характеризующим древний "плейстоценовый флористический комплекс", сохранившимся в современной флоре Монголии в качестве реликтов плейстоценового времени, можно отнести ряд видов лишайников. Как и среди плейстоценовых реликтов цветковых растений в современной флоре Южной Сибири, Средней Азии (Ревердатто, 1934, 1940; Овчинников, 1940; Соболевская, 1958, и др.), среди реликтовых видов лишайников можно выделить 2 группы: 1) группу перигляциально-степных реликтов; 2) группу гляциальных реликтов. Эти две группы реликтов были дифференцированы В. В. Ревердатто (1940) в работе, посвященной развитию послетретичной флоры Средней Сибири. В. В. Ревердатто полагал, что в Сибири в плейстоцене, во время максимального горного оледенения, высокогорная растительность под влиянием постепенно опускавшейся снеговой линии, а затем образовавшихся ледников мигрировала в долины и межгорные равнины, где происходило смешение высокогорной альпийской и степной ксерофитной флоры. В дальнейшем; в условиях холодного и сухого климата, часть "альпийцев", по-видимому, погибла, другие же сохранились до настоящего времени в составе современной степной и лесной флоры без изменений. Кроме того, в суровых условиях вблизи ледников как "альпийцы", так и степные ксерофиты образовали новые циклы форм. Исходя из этого, В. В. Ревердатто различает 2 группы плейстоценовых реликтов в современной флоре Сибири. Первая группа - гляциальные реликты (собственно) - это растения, существовавшие в альпийской зоне горных хребтов в периоды, предшествовавшие оледенению, и отчасти во время оледенения, и в настоящее время произрастающие преимущественно в высокогорном поясе, т. е. это типичные альпийцы или арктоальпийцы, и лишь в таких районах, как Монголия, Забайкалье, Средняя Сибирь, они встречаются в степях и лесах в нижних поясах гор и на прилегающих возвышенных равнинах. Вторая группа реликтов - перигляциально-степные реликты - это растения горных степей, развившиеся в условиях перигляциальной зоны; впоследствии они стали компонентами флор обширных пространств горных и предгорных степей Сибири.
К числу собственно гляциальных реликтов, или иногда их называют "сниженными альпийцами", в современной флоре Монголии можно отнести следующие виды лишайников.
Psoroma hypnorum (Vahl) S. Gray, Pannaria praetermissa Nyl., Peltigera scabrosa Th. Fr., Collema polycarpon Hoffm., Lecidea leptoboloides Nyl., L. athroocarpa (Ach.) Ach., L. limosa Ach., Psora rubiformis (Ach.) Hook., Toninia alutacea (Anzi) Jatta, Catillaria subnitida Hellb., Lecanora crustacea (Savicz) Zahlbr., Aspicilia cinereoruiescens (Ach.) Massal., A.-fimbriata (H. Magn.) Oxn., A. gyrodes (Nyl.) Hue., A. perradiata (Nyl.) Oxn., A. dudinensis (H. Magn.) Oxn., A. aquatica Koerb.
Pachyospora verrucosa (Ach.) Massal., Hypogymnia subobscura (Vain.) Poelt, Cladonia lepidota Nyl., Umbilicaria caroliniana Tuck., Lasallia pensylvanica (Hoffm.) Llano, L. pertusa (Ras- sad.) Llano, Acarospora badiofusca (Nyl.) Th. Fr., Blastenia leucoraea (Ach.) Th. Fr., Caloplaca epithallina Lynge, C. jungermanniaea (Vahl) Th. Fr., C. cinnamomea (Th. Fr.) Oliv., Solorina octospora Arnold, S. spongiosa (Sm.) Anzi, S. crocea (L.) Ach., Phaeophyscia constipata (Norrl. et Nyl.) Moberg.
Одни из этих видов обитают в горно-лесных ценозах Монголии, другие - в горно-степных или встречаются в горах по каменистым склонам. Большинство из них редкие на территории МНР лишайники, найденные в 1 - 2 местонахождениях, некоторые распространены более широко. Распространение этих лишайников связано главным образом с северными, в основном северо-западными районами страны. Особенно богата гляциальными реликтами флора Хангая.
В лесах Северной Монголии встречены такие арктовысокогорные лишайники, как Psoroma hypnorum (Vahl) S. Gray, Pannaria praetermissa Nyl., Catillaria subnitida Hellb., Hypogymnia subodscura (Vain.) Poelt, Cladonia lepidota Nyl., Phaeophyscia constipata (Norrl. et Nyl.) Moberg и некоторые другие виды. Помимо обитания этих арктовысокогорных видов в необычных для них экологических условиях, под пологом леса, о реликтовой природе их свидетельствуют изолированные участки их ареалов в пределах МНР, обычно далеко отстающие от основной части ареалов.
Примером может служить Psoroma hypnorum (Vahl) S. Gray. Этот арктовысокогорный мультирегиональный лишайник найден на территории Монголии в двух местонахождениях. Первое из них - в Хангае, в лиственничном лесу западнее г. Цэцэрлэг, на горе Булган-Ула, на высоте 1800 м над ур.м., второе - в южном Хэнтэе, в долине р. Тэрэлджийн-Гол, в лиственнично-березовом лесу, на высоте 1840 м над ур. м. (Schubert, Klement, 1971; Голубкова, 1981; Цогт, 1981). Основной ареал этого вида (Окснер, 1956; Инашвили, 1975; J∅rgensen, 1976) связан с арктическими районами Европы, Азии и Северной Америки, Гренландией, о-ва Ян-Майен. После обширной дизъюнкции Psoroma hypnorum (Vahl) S. Gray часто встречается в горных районах Европы, известен на Кавказе и Южном Урале, а также в Гималаях, за пределами Голарктики - в Южной Америке, Андах, достигает на юге Огненной Земли, о-ва Южная Георгия, а также растет в Австралии и Новой Зеландии. В Сибири широко распространен на п-ове Таймыр (Пийн, Мартин, 1978; Пийн, 1979а, 19796), в восточной Якутии (Окснер, 1939а; Городков, 1956), на Чукотском полуострове (Almquist, 1879, 1883; Nylander, 1888; Vafnio, 1909; Городков, 1938, 1958; Макарова, 1973, 1979; Макарова, Катенин, 1979), на п-ове Ямал (Андреев, 1982). В горных районах Азии до наших находок в Монголии известен был только в Гималаях (J∅rgensen, 1978).
В Хангае в лиственничном лесу горы Булган-Ула был собран и другой арктовысокогорный лишайник - Pannaria praetermissa Nyl. (Schubert, Klement, 1971), который также был найден в районе Хангая, в злаково-разнотравных степях, в окрестностях оз. Цаган-Нур. Помимо Хангая, этот вид был зарегистрирован в южном Хэнтэе, в кедрово-лиственничном лесу в хр. Барун-Борхын-Нуру и в лиственничном лесу в юго-западной части Монгольского Алтая (Голубкова, 1981; Цогт, 1981). Как отмечал Йоргенсен (J∅rgensen, 1978), монограф сем. Pannariaceae, P. praetermissa Nyl. имеет арктоальпийское распространение и является обычнейшим представителем рода Pannaria в Арктике. Этот вид часто встречается на севере Фенноскандии, но становится редким на равнинах в южных районах, довольно обычен в Исландии, а также в Шотландии; имеется несколько местонахождений его в Карелии, в северо-западной части Ладожского озера, откуда он и был описан. В остальной части Европы P. praetermissa Nyl. встречается вдоль горных хребтов от Пиренеев до Балкан. За пределами Европы этот лишайник встречается в Северной Америке, главным образом в арктических и субарктических районах, но вдоль хребта Скалистых гор проникает далеко на юг, вплоть до Калифорнии. Этот вид известен также на Кавказе. В Азии Р. рrаеtermissa Nyl. встречается вдоль берегов Северного Ледовитого океана; южнее, в горах Южной Сибири, имеется изолированное местонахождение этого лишайника на Алтае и в Кузнецком нагорье (Седельникова, 1978). Помимо находок P. praetermissa Nyl. в горно-лесном поясе в горах Монголии, этот вид был найден в Гималаях и в горных районах Японии. Изолированные местонахождения этого лишайника в различных регионах Азиатского континента позволяют предполагать, что, вероятно, в плейстоцене он мог иметь на территории Азии более обширный ареал.
Изолированное местообитание в Северной Монголии имеет Hypogymnia subobscura (Vain.) Poelt. Ареал этого лишайника охватывает арктическую Европу (о-ва Новая Земля, Шпицберген, Скандинавия) и высокогорья Средней Европы; в Альпах он встречается на высоте до 3000 м над ур.м., достигая нивального пояса (Lynge, 1928, 1934а; Poelt, 1962). В Северной Америке Н. subobscura (Vain). Poelt обычен на северном побережье Аляски; южнее имеется несколько изолированных местонахождений этого вида; в Канаде - на севере Квебека и в провинции Альберта, в США - в горах Колорадо (Ohlsson, 1973; Thomson, 1979). В Азии этот вид имеет преимущественно арктическое распространение. Он указывается для о-ва Преображения (Malme, 1932), п-ова Таймыр (Пийн, Трасс, 1971; Пийн, 1979а, 19796), восточной Якутии (Городков, 1956), Магаданской области (Локинская, 1970), Чукотского полуострова (Vainio, 1909; Malme, 1932; Городков, 1958; Локинская, 1970; Макарова, 1973, 1979а; Макарова, Катенин, 1979; Андреев, 1980), а также для Камчатки (Du Rietz, 1929; Трасс, 1963). Находки этого лишайника на территории МНР - первые в глубине Азиатского континента. В Монголии Н. subobscura (Vain.) Poelt найден в северо-западной части страны, в лиственничных лесах в хр. Цаган-Шибэту, а также в Хангае, в хр. Хан-Хухэй и на горе Булган-Ула, в окрестностях г. Цэцэрлэг; кроме того, он был обнаружен в южном Хэнтэе, в долине р. Тэрэлджийн-Гол, и на горе Богдо-Ула в окрестностях г. Улан-Батора (Schubert е. а., 1969; Schubert, Klement, 1971; Голубкова, 1981). Обитание этого арктовысокогорного лишайника в горнолесном поясе горных районов Монголии, вдали от основной части его ареала, позволяет отнести этот вид к числу "сниженных альпийцев" и рассматривать его как гляциальный реликт в современной флоре Монголии.
Помимо лиственничных лесов, "сниженные альпийцы" встречаются также в березовых и сосновых лесах Монголии. Так, например, в северо-западной части Средней Халхи, в горах Яргайт, в ущелье Сайрын-Худук в березовом лесу с примесью тополя и других деревьев лиственных пород был найден арктовысокогорный лишайник Phaeophyscia constipata (Norrl. et Nyl.) Moberg (Голубкова, 1981). Этот же вид приводится для Хангая (Бязров и др., 1983): он обитает на скалах среди мхов в светлых лесах среднегорной части госхоза Тэвшрулэх. В долине р. Онон, на северном склоне хребта Эрэн-Даба-Нуру, на высоте 1300 м над ур. м., в сосновом лесу довольно часто встречается арктовысокогорный вид Cladonia lepidota Nyl., который обитает здесь на скалах среди мхов и в их расщелинах (Голубкова, 1981; Цогт, 1981).
Помимо лесных сообществ, арктовысокогорные и высокогорные виды лишайников принимают участие в сложении степных ценозов Монголии. К числу таких видов можно отнести Psora rubiformis (Ach.) Hook., Acarospora chlorophana (Wahlenb.) Massal., A. badiofusca (Nyl.) Th. Fr., Lecidea leptoboloides Nyl., L. athroocarpa (Ach.) Ach., Lecidella inamoena (Mull. Ar.) Hertel, Lecanora crustacea Zahlbr., Aspicilia cinereorufescens (Ach.) Massal. Из них особенно широко распространенным видом является Acarospora chlorophana (Wahlenb.) Massal. Этот эпилитный арктовысокогорный голарктический лишайник часто встречается в горных степях Хангая, Хэнтэя, Хобдоского горно-степного района, Монгольского Алтая, Монгольско-Даурского района, Средней Халхи и Котловины Больших Озер (Klement, 1965; Schubert е. а., 1969; Schubert, Klement, 1971; Голубкова, 1981; Бязров и др., 1983).
На почве, в расщелинах и у основания скал в горных и равнинных степях Монголии нередок арктовысокогорный голарктический лишайник Psora rubiformis (Ach.) Hook. Он найден в Хангае, в Хобдоском горно-степном районе, в злаково-карагановых степях в Средней Халхе и Восточной Монголии (Schubert, Klement, 1971; Голубкова, 1981, Цогт, 1981).
На скалах и щебнисто-галечном панцире по склонам гор, в горных и горно-луговых степях, часто встречается Lecidella inamoena (Mull. Arg.) Hertel (Голубкова, 1981). Этот лишайник на территории Монголии был найден в районе Прихубсугулья, Хангая, Монгольского Алтая, Прихинганья и Хобдоского горно-степного района. Ареал L. inamoena (Mull. Arg.) Hertel охватывает все районы Арктики, высокогорья Европы, Азии и Северной Америки; в Южной Америке этот вид встречается в северных Андах (Hertel, 1971а, 1977). В Азии L. inamoena (Mull. Arg.) Hertel, помимо Монголии, известен из Малой и Средней Азии (Таласский Алатау, Ферганский хребет, Восточный Памир), а также в горах Непала, где обитает на высоте 3350 - 5000 м над ур.м. (Hertel, 1977).
В горных степях Хангая и Средней Халхи (Унджул сомон) на силикатных скалах на высоте 1350 - 1800 м над ур.м. был найден высокогорный евразиатский лишайник Lecidea leptoboloides Nyl. (Hertel, 1977). Основной ареал этого вида связан с горами Европы - Высокими Татрами, Южными Карпатами, Балканами, где он встречается главным образом в альпийском поясе. Кроме Монголии, в Азии он был собран в горах Турции и в Гималаях, где обитал на высоте 4340 - 5000 м над ур. м. (Hertel, 1971b, 1977).
Высокогорный лишайник Lasallia pertusa (Rassad.) Llano характеризуется дизъюнктивным ареалом, одна часть которого охватывает Южную Сибирь (Хакасская авт. обл., Прибайкалье, Забайкалье) и Северную Монголию, вторая - Гималаи (Llano, 1950; Frey, Poelt, 1977), третья - горы Эфиопии (Krog, 1973). Причем как в Гималаях, так и в горах Эфиопии L. pertusa (Rassad.) Llano обитает на больших абсолютных высотах - 3600 - 5050 м. На территории МНР этот вид ведет себя как типичный "сниженный альпиец". Он часто встречается на скалах в лиственничных лесах, по склонам гор, реже в горных степях на высоте 1300 - 1900 м над ур. м., изредка поднимаясь до высоты 2300 м над ур. м. (Голубкова, 1981). Широкие дизъюнкции в ареале этого вида, особенно между азиатской и восточноафриканской частями его ареала, свидетельствуют, видимо, о третичном возрасте этого лишайника. Распространение L. pertusa (Rassad.) Llano, с одной стороны, в Гималаях, а с другой - в Монголии и Южной Сибири говорит о существовании, видимо в плейстоцене, флористических связей между этими двумя районами Азии.
Нередко несколько видов из числа "сниженных альпийцев" встречаются в одном местонахождении, в одних и тех же условиях местообитания. Например, в долине р. Чулутын-Гол в Хангае на базальтовых скалах, сильно задернированных, поросших мхами, совместно обитает несколько арктовысокогорных видов рода Solorina - S. octospora Arnold, S. spongiosa (Sm.) Anzi, S. saccata (L.) Ach. Вместе с ними произрастал также гипоарктомонтанный, редкий в Монголии лишайник - Peltigera venosa (L.) Baumg. Виды рода Solorina - очень редкие на территории Монголии лишайники, причем S. octospora Arnold и S. spongiosa (Sm.) Anzi встречаются только в Хангае (Голубкова, 1981; Цогт, 1981). Это местонахождение, в долине р. Чулутын-Гол, изолировано от остальной части их ареалов и свидетельствует о его реликтовом характере. Так, ареал S. octospora Arnold связан в основном с арктическими и горными районами Европы, в Азии имеются лишь изолированные местонахождения этого лишайника - на Таймыре, в Якутии, на Чукотском полуострове, Алтае и в Монголии (Савич, 1975; Макарова, 1979а; Пийн, 1979а, 1979б).
Дизъюнктивный ареал Solorina spongiosa (Sm.) Nyl. охватывает Арктику (Новая Земля, Исландия, острова Медвежий, Преображения, Кольский полуостров) и горы Средней Европы, Кавказ; этот лишайник известен также в Прибалтике (о-в Моон). В Азии имеются разрозненные местонахождения этого вида - на п-ове Таймыр (Пийн, Мартин, 1978), в западной части Чукотского полуострова, на Камчатке (Савич, 1922; Трасс, 1963), в Монголии (Хангай) (Голубкова, 1981; Цогт, 1981) и в Средней Азии (Терскей-Ала-Тоо).
В Северной Америке этот вид встречается от Аляски и о-ва Баффинова Земля до о-ва Ньюфаундленд, оз. Онтарио, штатов Колорадо и Вашингтон (Thomson, 1979).
Ряд видов лишайников в составе флоры Монголии отнесен нами к перигляциально-степным реликтам. Среди них Aspicilia transbaicalica Oxn. - эпилитный лишайник, обитающий на валунах и выходах горных пород в горных степях Хангая, Хэнтэя, Монгольско-Даурского горно-степного района, в северных районах Средней Халхи (рис. 42). О его реликтовой природе свидетельствует прежде всего дизъюнктивный ареал. Помимо Монголии, ареал этого вида охватывает горные степи Забайкалья (Окснер, 1934), Саян и северного Алтая (Окснер, 19716; Седельникова, 1977б). Далее к западу A. transbaicalica Охn. встречается в Центрально- Казахстанском мелкосопочнике (Андреева, 1961; Бредкина, Макарова, 1971), а после западносибирской дизъюнкции снова появляется на Урале (Башкирская АССР), в Предуралье (Куйбышевская область) и на восточном склоне Приволжской возвышенности (Окснер, 1971б). Таким образом, в распространении A. transbaicaiica Охn. можно отметить особенности, характерные для ареалов реликтовых видов перигляциальных степей (Крашенинников, 1954а, 19546; Лавренко, 1981). Кроме того, хорошо прослеживаются тесные генетические связи A. transbaicaiica Охп. с арктоальпийскими формами. Вместе с эндемиком горно-степных районов Северной Монголии - A. changaica (Klem.) Golubk. - этот вид образует один систематический ряд с европейскими видами - арктоальпийским видом A. manstrucata (Wahlenb.) Ach. и высокогорным Lecanora (Aspicilia) nunatakkorum Poelt (Klement, 1965). Такая тесная связь с субальпийскими и арктоальпийскими формами, как полагал И. М. Крашенинников (1954а), указывает на вероятность происхождения этих элементов флоры от древних бореально-ксерофитных биотипов "плейстоценового флористического комплекса". И. М. Крашенинников подчеркивал, что часто подобные систематические серии и еще более обширные циклы близких видов в своем распространении охватывают не только Урал и горную Сибирь, но также горы Средней Азии, Кавказа, Крыма, Западной Европы, обычно с дизъюнкциями, падающими на разделяющие эти центры равнины.
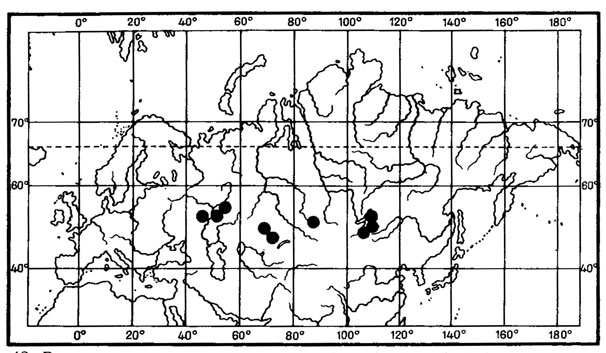
Рис. 42. Распространение на земном шаре Aspicilia transbaicalica Oxn. (Ориг.)
К числу перигляциально-степных реликтовых видов во флоре МНР, видимо, можно отнести Phaeorrhiza sareptana (Tomin) I Mayrh. et Poelt, представленный в этом районе разновидностью - var. sphaerocarpa (Th. Fr.) Mayrh. et Poelt (рис. 44). Это горный ксерофнтный лишайник, обитающий на почве между камнями, на дерновинках мхов и растительных остатках в горных степях различного типа, особенно в условиях их петрофильных вариантов, на кобрезиевых пустошах, иногда среди березово-лиственничной уремы, в пределах высот 1500 - 3400 м над ур.м. Как можно видеть из карты распространения этого вида на территории МНР (Голубкова, 1981), его ареал в Монголии связан в основном с северо-западными районами страны - Прихубсугульем, Хангаем, Хобдоским горно-степным районом, Монгольским Алтаем. Изредка этот лишайник встречается в Средней Халхе, а также в северо-западных хребтах Гобийского Алтая (хр. Баян-Цаган-Ула). Как в случае Aspicilia transbaicalica Охn., о реликтовой природе этого лишайника свидетельствует значительная дизъюнкция его ареала. В Азии, за пределами Монголии, Phaeorrhiza sareptana (Tomin) Mayrh. et Poelt собирался А. А. Еленкиным в Саянах (образцы в Гербарии БИН АН СССР); кроме того, этот вид встречается в реликтовых степных комплексах Северо-Восточной Азии - в районе р. Индигирки (Афонина и др., 1979, 1980; Юрцев, 1981), где был отмечен в келериевых, овсецовых, твердовато-осочковых и петрофитных степях, а также в лугостепях. За пределами Восточной Сибири и Монголии известен из Казахстана - из Центрально-Казахстанского мелкосопочника (Андреева, 1961). После обширной дизъюнкции этот лишайник встречается в Приволжье - на восточной окраине Приволжской возвышенности, в Саратовской области, где был собран по склонам в пустынных и полынно-типчаковых степях и южнее в окрестностях Волгограда. Причем только в Приволжье отмечена вторая разновидность этого вида - var. sareptana. Западнее, после обширной дизъюнкции, Ph. sareptana (Tomin) Mayrh. et Poelt появляется в Альпах. Имеются еще два изолированных местонахождения этого лишайника - в Северо-Западной Норвегии и в Северной Америке (США - горы штата Колорадо) (Mayrhofer, Poelt, 1978). Указание этого вида для Антарктики (Lamb, 1968) Майргофер и Пельт ставят под сомнение.
Phaeorrhiza sareptana (Tomin) Mayrh. et Poelt относится к олиготипному роду, в составе которого имеется еще один вид - Ph. nimbosa Mayrh. et Poelt. Последний является арктоальпийским, широко распространенным на земном шаре лишайником. Однако Ph. nimbosa Mayrh. et Poelt относится к числу криоксерофитных лишайников и как в высокогорьях, так и в Арктике обычно встречается в сухих, хорошо освещенных местообитаниях. Морфологически эти два вида очень близки между собой и долгое время считались одним видом. Реликтовый ареал Ph. sareptana (Tomin) Mayrh. et Poelt, а также его широкое распространение в реликтовых степных комплексах Восточной Сибири, двойственность его экологии (обитание как в среднегорных, так и в высокогорных поясах), генетическая близость с арктоальпийской формой - все это дает основание причислить этот вид к группе видов "плейстоценового флористического комплекса".
К группе перигляциальных реликтов в флоре Монголии, видимо, можно отнести Caloplaca tominii Savicz (рис. 43). Основная часть ареала этого ксерофитного напочвенного лишайника охватывает обширные пространства от низовьев Волги (Савич, 1930; Келлер, 1936), Казахстана до горных районов Средней Азии, Монголии и Восточной Сибири, где он встречается спорадически главным образом в составе криофитных степных и пустынных сообществ горных районов. С. tominii Savicz рассеянно растет в Средней Азии, где он был собран на каменистых горных склонах Алайского хребта и адырах Ферганской долины (Шафеев, 1953а, 1953б). На Восточном Памире этот вид являемся одним из компонентов синузий эпигейных лишайников в составе высокогорных криофитных пустынь, в пределах высот 3800 - 4100 м над ур. м. (Бредкина, Голубкова, 1977), приводится также для афганской территории Памира (Poelt, Wirth, 1968). Имеются сведения о находках этого лишайника в Центральном Тянь-Шане, в криофитных степях отрогов хр. Кызарт и Сусамырской долины, на высоте 2100 - 2600 м над ур.м. (Бредкина, 1981а), а также в центральном Копетдаге и северо-западной Туркмении (Джураева, 1974, 1978). В Казахстане С. tominii Savicz распространен в Центрально-Казахстанском мелкосопочнике (Окснер, 19396; Андреева, 1961). Далее к востоку этот лишайник встречается в северо-западных районах МНР, где является редким видом. Он был собран на карбонатной почве и растительных остатках на горном каменистом склоне в окрестностях оз. Дод-Hyp в Дархатской котловине в Прихубсугулье и в Монгольском Алтае, в хр. Дарвийн-Нуру, в горной степи на высоте 2260 м над ур. м. (Цогт, 1976б; Голубкова, 1981; Цогт, 1981). В Восточной Сибири распространение этого лишайника связано с реликтовыми степными сообществами лесостепного ландшафта среднего течения р. Индигирки, где он обитает на почве в твердоосочково-житняковых, житняковых и келерево-житняко- вых горных степях на эродированных участках (Афонина и др., 1979, 1980; Юрцев, 1981). Кроме того, С. tominii Savicz недавно был найден в реликтовых степных сообществах в бассейне р. Юкон, в северо-западной части Канады (Nimis, 1981). Здесь, в районе оз. Клуэйн, на высоте 450 - 700 м над ур. м., этот лишайник обитает на лёссовых отложениях, столь характерных для перигляциальных зон, в ассоциации Artemisia - Agropyron yukonensis. В этом растительном сообществе Нимисом была описана эпигейная лишайниковая синузия Fulgensio - Caloplacacetum tominii. Нимис рассматривает С. tominii Savicz перигляциальным реликтом во флоре этого района. По сообщению Томсона (Thomson, 1982), ревизия американских образцов Caloplaca cirrochra (Ach.) Th. Er. показала, что многие из них являются С. tominii Savicz, в том числе, например, образцы, приводимые в работе Лумана (Looman, 1964), посвященной эпигейным группировкам лишайников североамериканских прерий. Таким образом, приводятся новые местонахождения этого вида на территории Северной, а также Южной Америки. В Канаде, помимо бассейна р. Юкон, С. tominii Savicz встречается на о-ве Элсмир и в провинции Саскачеван, в США - в штатах Вашингтон, Монтана, Вайоминг и Южная Дакота, в Южной Америке - в Перу. Имеются еще два изолированных участка ареала этого вида. Один из них в восточной Гренландии, другой - в Южной Норвегии (Lynge, 1940; Nordin, 1972), где он известен из 13 местонахождений в районе, характеризующемся крайне континентальным климатом. Как предполагает Алнер (Ahlner, 1949), миграция С. tominii Savicz в Норвегию, возможно, произошла в предбореальный период. Обширный дизъюнктивный ареал этого вида, приуроченность изолированных участков ареала к реликтовым комплексам Якутии и Северо-Западной Канады, его изолированные местонахождения на территории Монголии, его экология, связь с криофильными степными и пустынными сообществами - все это дает основание отнести этот вид к числу перигляциальных реликтов во флоре Монголии. С. tominii Savicz принадлежит к секции Gasparrinia рода Caloplaca. Представители этой секции преимущественно эпилитные лишайники, широко распространенные в горных и пустынно-степных районах земного шара. Особенно богата видами этой группы флора Средиземноморья, а также горные районы Средней Европы, где эти лишайники встречаются в сухих, хорошо прогреваемых и освещаемых местообитаниях (Poelt, 1954). Генетически С. tominii Savicz наиболее близок к С. citrina (Hoffm.) Th. Fr. - виду с широкой экологией, встречающемуся на различных субстратах (известняковые скалы, мрамор, гниющее дерево, почва и др.) и широко распространенному по всей Голарктике (Европа, Азия, Северная и Центральная Америка), найденному также и на территории МНР (Schubert, Klement, 1971; Бязров, 1976). Эти два вида, характеризующиеся соредиозными слоевищами, морфологически очень близки друг к другу. Слабая морфологическая дифференциация С. tominii Savicz и его распространение в пределах ареала С. citrina (Hoffm.) Th. Fr. свидетельствуют о его молодости. Можно предположить, что его обособление произошло в плейстоцене, а ареал оформился в перигляциальной зоне во время оледенения.
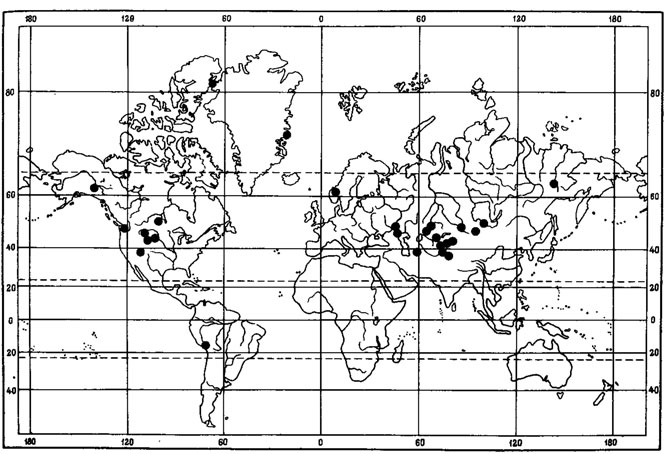
Рис. 43. Распространение на земном шаре Caloplaca tominii Savicz. (Ориг.)
Таким образом, на территории Монголии "плейстоценовый флористический комплекс" сочетает разнородные по экологии и происхождению элементы лихенофлоры. Он представлен как мезофильными, так и криоксерофитными арктовысокогорными и высокогорными формами, а также криоксерофитными горно-степными видами лишайников. Эволюция некоторых из этих форм, видимо, протекала в ксерических условиях горно-степных районов Южной Сибири и Северной Монголии.
Помимо отдельных видов, сохранившихся в современной флоре Монголии в качестве реликтов плейстоценового времени, в некоторых районах страны можно наблюдать своеобразные лишайниковые группировки, которые, вероятно, можно отнести к числу реликтовых, характеризующих древний "плейстоценовый флористический комплекс". Например, в Монгольском Алтае на некоторых участках по нижней границе лесного пояса в лиственничных лесах, особенно в лиственничниках ритидиевых или ритидиево-разнотравных, можно наблюдать эпигейные группировки лишайников, образованные типичными тундровыми видами - Cetraria nivalis (L.) Ach., С. cucullata (L.) Ach., Thamnolia vermicularis (L.) Ach., Alectoria ochroleuca (Hoffm.) Massal. В их составе и некоторые свободноживущие, кочующие формы - Evernia perfragilis Llano, Cornicularia aculeata (Schreb.) Ach. Такое сообщество арктовысо- когорных лишайников в составе лесных ценозов, по-видимому, можно рассматривать как реликтовое, сохранившееся в современном растительном покрове с плейстоценового времени.
В некоторых районах Северо-Западной Монголии, особенно по каменистым склонам, можно встретить совместное обитание тундровых и степных видов лишайников. Например, в долине р. Тургэн-Гол по каменистому склону в лиственничном редколесье среди травянистого покрова между разбросанными валунами и отдельными лиственницами совместно с горно-степными и степными видами лишайников, такими как Phaeorrhiza sareptana (Tomin) Mayrh. et Poelt (рис. 44), Lecidea asiaecentralis H. Magn., Acarospora schleicheri (Ach.) Massal., Parmelia taractica Krempelh., Diploschistes bryophilus (Ehrh.) Zahlbr., обитали арктовысоко-горные виды - Stereocaulon alpinum Laur., Cetraria nivalis (L.) Ach., Lecidea limosa Ach., Peltigera lygnei Gyeln., гипоарктомонтанный вид Physconia mustigena (Ach.) Poelt, бореальный Cornicularia aculeata (Schreb.) Ach. и другие лишайники. Возможно, что такое характерное сочетание типично тундровых и степных видов лишайников могло существовать в перигляциальных тундрово-cтепных фитоценозах. Следует отметить, что видовой состав вышеупомянутого сообщества весьма сходен с составом лишайников реликтовых степных комплексов среднего течения р. Индигирки (Афонина и др., 1979, 1980; Юрцев, 1981).
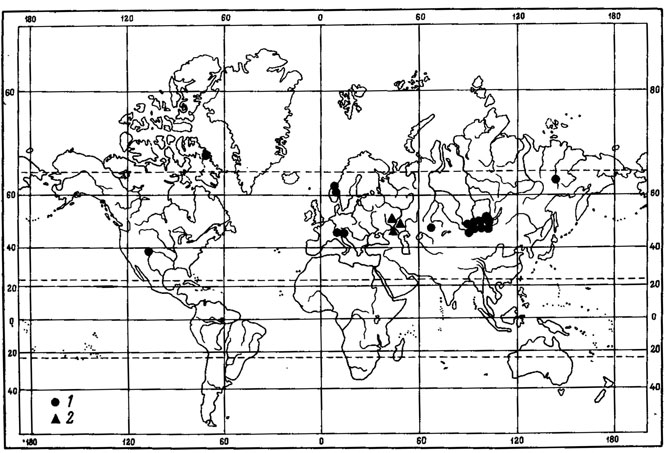
Рис. 44. Распространение на земном шаре Phaeorrhiza sareptana (Tomin) Mayrh. et Poelt var. sareptana (1) и var. sphaerocarpa (Th. Fr.) Mayrh. et Poelt (2). (По: Mayrhofer, Poelt, 1978, с дополнениями автора)
Реликтовые элементы флор голоцена. Голоценовая группа реликтов включает виды, проникшие на территорию Монголии во время климатического оптимума, когда происходило потепление климата. О наличии климатического оптимума во время голоцена имеется достаточное количество данных (Берг, 1947; Марков и др., 1965; Марков, Величко, 1967). Для этого времени характерно изменение границ растительных зон по направлению к северу. Например, в Европе зона умеренных лесов продвинулась к северу на 5°, в Норвегии леса вышли на северное побережье, средиземноморская растительность перешагнула через Альпы и продвинулась к северу на 2 - 3° (Монин, Шишков, 1979). Значительно смещались также северные границы ареалов растений в высоких широтах, что сопровождалось потеплением климата в верхнем голоцене (Сукачев, 1922; Тихомиров, 1941, и др.). Заметно переместилась вверх и верхняя граница леса в горах. Для тропических и экваториальных областей также имеются данные о существовании климатического оптимума. Теплый и влажный климат был также и в ныне засушливых районах Азии (Монин, Шишков, 1979). На Дальнем Востоке средний голоцен характеризуется широким распространением широколиственных и хвойно-широколиственных лесов за пределы их ареалов (Минкина, Федорова, 1936; Нейштадт, 1957; Гроссет, 1962). В Восточной и Средней Сибири распространились лиственничные леса со значительным участием пихты, кедра, ели, расширились площади темнохвойных лесов, в составе которых имелось значительное число видов неморальной флоры (Ханминчун, 1980).
На территории МНР суббореальный период голоцена также характеризуется климатическим оптимумом (Виппер и др., 1978; Савина и др., 1981). Здесь господствовал влажный и довольно теплый климат, происходило обводнение озер и резко увеличилась облесенность горных склонов всех экспозиций. В Хангае, например, в этот период преобладают кедрово-лиственничные и кедровые леса с елью, лиственнично-сосновые и другие леса в сочетании с горно-лугово-лесными группировками, по южным склонам были развиты лугово-разнотравные степи. Максимальная облесенность в суббореальный период была характерна и для Хэнтэя, а также для районов северо-востока Монголии, долин рек Онон и Улдза. В Хэнтэе, например, в это время господствовали лиственничные леса с кедром, елью, было много сосны. Здесь же расширялись площади темнохвойных лесов; в долину р. Улдза проникали широколиственные породы (Голубева, 1978).
В составе голоценовых реликтов среди лишайников можно выделить 2 группы видов: группу "пацифических" и группу "атлантических" реликтов. "Пацифическими" реликтами М. М. Ильиным (1941) была названа группа третичных реликтов в таежной флоре Сибири, центром происхождения которых явились Восточная Азия и отчасти Северная Америка. После дизъюнкции эти виды вновь появляются в Алтайских и Саянских горах, отсутствуя в Европе. Группа видов, растущих, помимо горных районов Сибири, в лесах Европы и Кавказа, но на Дальнем Востоке или вообще в Восточной Азии не встречающихся, М. М. Ильиным была названа "атлантическими" реликтами, но, как отмечал автор, не в узком значении этого слова, а в смысле проникновения их в Сибирь с западной оконечности Евразиатского материка.
Рассмотрим вначале группу "пацифических" реликтов. В ее составе можно выделить 3 подгруппы: 1) виды восточноазиатского распространения; 2) виды "пацифического" распространения; 3) виды тропического распространения. Мы их объединяем в группу "пацифических" реликтов в широком смысле этого слова, так как все эти виды имеют ареалы (или участки ареалов), приуроченные к районам Тихоокеанского побережья (азиатского, североамериканского, австралийского) или тихоокеанских островов, а в пределах Алтае-Саяно-Хангайской горной страны имеют изолированные местонахождения. Можно предположить, что иммиграция этих видов на территорию Монголии происходила в голоцене с востока.
В составе реликтовых видов восточноазиатского распространения Hypogymnia fragillima (Hillm.) Rassad., H. duplicatoides (Oxn.) Rassad., Phaeophyscia hirtuosa (Krempelh.) Golubk., Parmelia cochleata Zahlbr., Cetraria rugosa (Asah.) Sato. Все они - лесные мезофильные эпифитные лишайники, относящиеся к бореальному и неморальному элементу.
Основной ареал Hypogymnia fragillima (Hillm.) Rassad. связан с о-вом Сахалин, Приморским краем (Сихотэ-Алинь), Японией, п-овом Корея (Рассадина, 1953, 1956, 1971). Его западная граница, видимо, проходит в районе Северо-Западного Китая, в провинции Хэйлунцзян, в южных отрогах Малого Хингана (Гербарий БИН АН СССР). В этих районах Н. fragillima (Hillm.) Rassad. обитает в хвойных и смешанных лесах, где растет на стволах деревьев, валеже, изредка на замшелых скалах. На территории МНР имеется изолированное местообитание этого вида в Центральном Хангае, в долине р. Хух-Сум. Здесь он обитает в кедрово-лиственничном лесу на коре лиственницы, на высоте 2000 м над ур. м. (Бязров и др., 1983). Как уже отмечалось, современный центр наибольшего видового и типового разнообразия рода Hypogymnia находится в районе Восточной Азии. Н. fragillima (Hillm.) Rassad. относится к секции Delavayi Rassad., объединяющей наиболее примитивные и, видимо, наиболее древние формы этого рода; большинство из них является восточноазиатскими видами. Изолированный участок ареала этого вида на территории Монголии, по-видимому, реликтовый и свидетельствует о более обширном ареале этого вида, возможно, в период климатического оптимума голоцена.
Второй вид гипогимний - Hypogymnia duplicatoides (Oxn.) Rassad. - относится к секции Metaphysodes Rassad. Основной ареал этого лишайника связан с Японией и Дальним Востоком (Приморским и Хабаровским краями), где он обитает на стволах деревьев хвойных пород, валеже в кедрово-березовых редколесьях, заболоченных лиственничных марях, в хвойных лесах (Окснер, 1938; Рассадина, 1956, 1971). В Монголии, в Хангае, имеется изолированный участок ареала этого вида. В этом районе он является редким видом, растущим в лиственничных и кедрово-лиственничных лесах среднегорного пояса на коре деревьев хвойных пород (Бязров и др., 1983). Как и Н. fragillima (Hillm.) Rassad., этот дальневосточный лишайник, видимо, во время климатического оптимума в голоцене расширил свой ареал одновременно с увеличением площадей темнохвойных и лиственничных лесов, достигнув на западе центральных районов Монголии. Распад ареалов этих видов мог произойти уже в субатлантический период в связи с возрастанием континентальности климата, сокращения облесенности гор.
Остальные виды этой реликтовой группы - неморальные лишайники; они связаны в своем распространении с широколиственными породами. Основная часть ареала Phaeophyscia hirtuosa (Mereschk.) Moberg (табл. IV, 1) охватывает леса Японии, Восточного Китая (Zahlbruckner, 1930), Дальнего Востока (Приморский край). После значительной дизъюнкции этот вид появляется в Прихинганье, где был собран в пойме долины р. Нумэргин-Гол на коре вяза (Голубкова, 1981). На территории Монголии известны и другие изолированные местообитания этого вида, Он был найден в лесу на коре деревьев лиственных пород, в 60 км северо-восточнее г. Улан-Батора (Vezda, 1965), а также в Хангае, в долине р. Эгин-Гол, в лиственничнике с зарослями кустарников.
Основной ареал Cetraria rugosa (Asah.) Sato связан с лесами Японии; изолированное местонахождение этого вида имеется в Центральном Хангае, западнее г. Цэцэрлэг, на горе Булган-Ула (Schubert, Klement, 1971). С. rugosa (Asah.) Sato относится к секции Nephromopsis (Рассадина, 1950), представители которой являются главным образом восточноазиатскими и тропическими лишайниками.
В Хангае был собран и другой неморальный вид - Parmelia cochleata Zahlbr. (Schubert, Klement, 1971), основной ареал которого также приурочен к лесам Японии (Zahlbruckner, 1927). Проникновение этих неморальных элементов в восточные районы Монголии могло произойти в среднем голоцене в результате расширения ареала дальневосточных широколиственных и хвойно-широколиственных лесов, откуда они могли проникнуть и в Хангай. Возрастающая аридность климата после климатического оптимума в голоцене, видимо, явилась причиной разъединения ареалов этих мезофильных лесных видов.
К реликтовой группе видов "пацифического" распространения относится Hypogymnia enteromorpha (Ach.) Nyl. Дизъюнктивный ареал этого лишайника приурочен к побережьям Тихого океана. На Дальнем Востоке он встречается на побережье Охотского моря, в Магаданской области - в окрестностях г. Магадана, в Хабаровском крае - в окрестностях Аяна и нижнем течении р. Уды, на побережьях о-ва Сахалин (Рассадина, 1956), где обитает на стволах и ветвях деревьев хвойных пород, на мхах в хвойных лесах, сфагновых болотах, в кедровых стланиках. Помимо этих районов, в Восточной Азии он отмечен в восточной части Китая - в провинции Шаньси и на о-ве Тайвань. На территории Северной Америки ареал Н. enteromorpha (Ach.) Nyl. приурочен к Тихоокеанскому побережью, от Аляски до Калифорнии. Кроме того, этот лишайник растет также в Австралии, Новой Зеландии и на о-ве Тасмания. Это вид гипогимнии восточноазиатских филогенетических связей. Он относится к секции Delavayi Rassad. подрода Tubulosa (Bitt.) Krog. Как уже отмечалось, представители этой секции входят в число наиболее примитивных форм в роде Hypogymnia; центром их видового многообразия является район Восточной Азии. Генетически Н. enteromorpha (Ach.) Nyl. наиболее близок к восточноазиатско-американскому виду Н. delavayi (Hue) Rassad. Эти виды вместе с восточно-азиатским видом Н. fragillima (Hillm.) Rassad. и восточноазиатско-американским видом Н. duplicata (Sm.) Охn. образуют один систематический ряд. Центральное место в этом году занимает эпифитная фертильная форма Н. Delavayi (Hue) Rassad. А. Н. Окснер (1974) относил этот вид, по-видимому древний, к реликтам третичного времени во флоре Приморского края. К реликтам третичного времени во флоре Дальнего Востока, вероятно, можно причислить и Н. enteromorpha (Ach.) Nyl., о чем свидетельствует его обширный дизъюнктивный ареал, а также его более примитивный для лишайников способ размножения с помощью спор, в то время как большинство представителей этого рода размножается вегетативно, фрагментами слоевища, соредиями, изредка изидиями. Крог (Krog, 1968) относила этот вид к амфипацифическому элементу; По ее определению, амфипацифический элемент объединяет виды, приуроченные к побережьям Тихого океана, но отсутствующие в районе Берингового моря и Берингового пролива. Как она полагала, эти виды являются представителями древней, уже дифференцированной амфипаци- фической третичной флоры, которая разорвала свой замкнутый ареал, приуроченный к районам Тихого океана, в северной его части, когда мигрировала в более южные районы в связи с похолоданием климата в третичное время, может быть, в середине олигоцена или позднего миоцена. На территории Монголии, в Хангае, имеется изолированный участок ареала этого океанического вида (Бязров и др, 1983). По свидетельству Л. Г. Бязрова, в восточной части Хангая, на северном макросклоне магистрального хребта, этот лишайник растет на коре лиственницы и кедра, на валеже в лесах среднегорного и высокогорного поясов, но при этом встречается редко. Можно предположить, что этот третичный океанический вид в одну из влажных и теплых фаз голоцена (во время климатического оптимума) мог расширить свой восточно-азиатский ареал к западу, вплоть до горных районов Монголии, а впоследствии, в позднем голоцене, в связи с усилением континентальноcnb климата вновь сократить его, оставив на территории Хангая небольшой изолированный реликтовый участок ареала.
Группа реликтовых видов тропического распространения объединяет лишайники, основное распространение которых связано с тропическими районами земного шара, а также районами Восточной Азии. На территории Сибири и Монголии имеются изолированные местонахождения этих лишайников. К этой группе реликтов относятся виды рода Lobaria - L. isidiosa (Mull. Arg.) Vain., L. retigera (Bory) Trevis. (рис. 45), Parmelia rudecta Ach., Pseudevernia cirrhata (Fr.) Schub. et Klem.
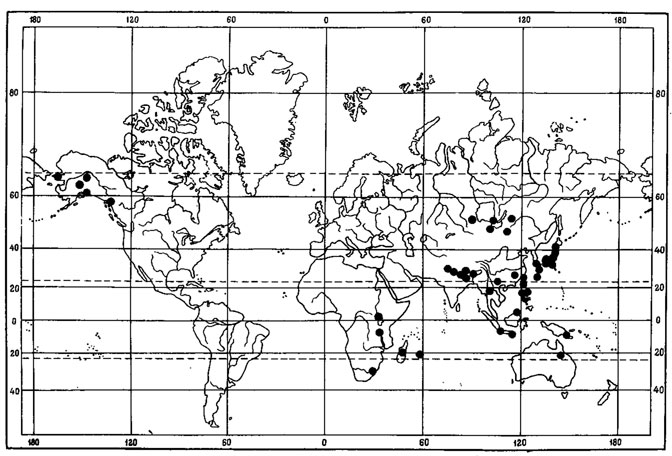
Рис. 45. Распространение на земном шаре Lobaria retigera (Bory) Trevis. (По: Yoshimura, 1971, с дополнениями автора)
Как уже указывалось, род Lobaria насчитывает 70 видов, основное распространение которых связано с тропическими и субтропическими районами земного шара. Центром наибольшего видового разнообразия рода являются тропические и субтропические области бассейна Тихого океана, где встречается большое число эндемиков из этой систематической группы лишайников. В умеренных районах Голарктики особенно богаты лобариями флоры Дальнего Востока, приатлантических районов Европы и Северной Америки. Основной ареал Lobaria isidiosa (Mull. Arg.) Vain. (Yoshimura, 1971) связан с Индией, о-вом Шри Ланка, Бутаном, СРВ, Таиландом, Филиппинскими островами, а также с островами Калимантан, Ява, Суматра, Сумбава, Новая Гвинея (рис. 46). В этих районах он обитает на замшелых скалах и часто на стволах деревьев. В более северных районах этот лишайник встречается в Юго-Восточном Китае (провинция Юньнань, о-в Тайвань), Японии (о-в Хонсю), на Дальнем Востоке. Изолированное местообитание этого лишайника в глубине Азиатского материка, в районе Восточного Саяна, на границе Иркутской области и Монголии, в долине р. Эхе-Огун, видимо, реликтовое.
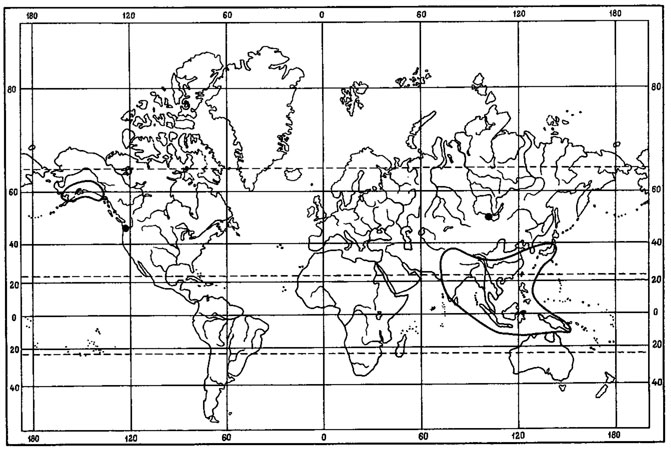
Рис. 46. Распространение на земном шаре Lobaria isidiosa (Mull. Arg.) Vain. (По: Yoshimura, 1971)
Lobaria retigera (Bory) Trevis. (рис. 45) характеризуется еще более обширным ареалом, охватывающим Восточную и Юго-Восточную Азию, Японию, Китай, Таиланд, Непал, Филиппинские острова, острова Ява, Новая Гвинея, западные тихоокеанские острова, а также Австралию, Центральную и Юго-Восточную Африку, о-в Мадагаскар, Маскаренские острова, где он обитает на замшелых скалах и на основаниях стволов деревьев (Yoshimura, 1971). Севернее известны изолированные местообитания этого лишайника - на Дальнем Востоке (Блюм, 1975), в Забайкалье (Окснер, 1946а), на Алтае (Рассадина, 1938), в Горной Шории (Седельникова, 1977а, 19776), на территории Монголии (Прихубсугулье, Хэнтэй) и на тихоокеанском побережье Аляски (Krog, 1968). На территории Монголии этот вид распространен довольно широко в районе Прихубсугулья, где обитает в горах на замшелых скалах в лиственничных и березовых лесах; он был найден в бассейнах рек Эгийн-Гол и Аригийн-Гол (Голубкова, 1981). Еще одно местообитание этого лишайника было обнаружено в южном Хэн- тэе, в долине р. Тэрэлджийн-Гол, в лиственнично-березовом лесу с подлеском из рододендрона; здесь он довольно часто встречается на замшелых валунах и скалах.
Изолированное, наиболее северное местообитание в пределах Хангайской горной страны имеет также эпифитный лишайник Pseudevernia cirrhata (Fr.) Schub. et Klem. (рис. 47). Ареал этого лишайника охватывает Дальний Восток (Приморский край), Японию, п-ов Корея, Китай, южную часть Северной Америки, Центральную Америку, западные районы Южной Америки (Рассадина, 1965б). Изолированное, далеко отстающее от восточно-азиатской части ареала местообитание этого вида находится в Северо-Западной Монголии, в хр. Хан-Хухэй и в окрестностях Тургэн сомона (Schubert, Klement, 1971).
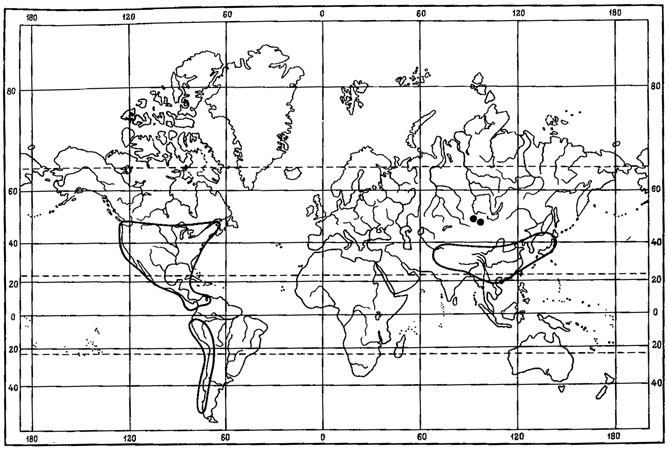
Рис. 47. Распространение на земном шаре Pseudevernia cirrhata (Fr.) Schub et Klem. (По: Рассадина, 1965б, с дополнениями автора)
По данным Л. Г. Бязрова и др. (1983), в Хангае, в Булганском аймаке, на юге Дашинчилен сомона, на горе Цэцэрлэг, на скалах им был собран Parmelia rudecta Ach. Это местонахождение также далеко оторвано от основной части азиатского ареала вида, которая связана с лесами Дальнего Востока (Приморского края) и Японией. Кроме того, этот вид известен из Австралии, Африки (Натал (ЮАР), Мадагаскар, мыс Доброй Надежды), Северной и Южной Америки (Окснер, 1934; Рассадина, 1971). Он относится к группе видов Parmelia borreri, объединяющей 3 вида - P. borreri (Sm.) Turn., P. rudecta Ach., P. subrudecta Nyl. В настоящее время эта группа видов вместе с другой группой пармелий - P. flaventior Stirt. - объединены Крог в род Punctelia Krog (Krog, 1982). Причем, как показала Крог, центром видового многообразия этого рода являются Африка и Америка, в то время как в Азии известны только 3 вида из этой систематической группы лишайников (виды группы P. borreri). Эти же виды встречаются и в Австралии. Как и P. rudecta Ach., P. borreri (Sm.) Turn, и P. subrudecta Ach. также с широкими мультирегиональными ареалами, охватывающими и Американские, и Африканский континенты, и, видимо, генетически связаны с флорами одного из районов. А. Н. Окснер (1974) относил P. rudecta Ach. к элементам третичной флоры во флоре Приморского края. Однако впоследствии он причислил P. rudecta Ach. к группе реликтовых видов в лесах третичного типа Приморской области, "климатические потребности которых находятся в некотором противоречии с современными климатическими отношениями наших широт" (Окснер, 1974, с. 233). При этом А. Н. Окснер отмечал, что пока еще не ясно, все ли эти реликты относятся к третичному времени, или они должны быть синхронизированы с номской теплой эпохой уже четвертичного времени. А. С. Лазаренко (1939), рассматривая вопрос о реликтовых видах мхов в бриофлоре Дальнего Востока, считал виды мхов, характеризующиеся широкими пантропическими и палеотропическими ареалами, поздними пришельцами во флоре Дальнего Востока, инвазию которых он связывает с коралловым веком Японии. В то же время Н. В. Седельникова (1977б) относила, например, Lobaria retigera (Bory) Trevis. к числу реликтов неморальной флоры Кузнецкого рефугиума, возраст которых ею определялся плиоценом.
Во флоре Монголии вышеуказанную группу тропического распространения - Lobaria isidiosa (Mull. Arg.) Vain., L. retigera (Bory) Trevis., Parmelia rudecta Ach., Pseudevernia cirrhata (Fr.) Schub. et Klem. - мы относим к реликтам флоры, существовавшей на территории Северной Монголии во время климатического оптимума голоцена (суббореальный период). Широкие мультирегиональные ареалы этих видов, приуроченные главным образом к тропикам и субтропикам Старого и Нового Света, свидетельствуют о древнем, возможно третичном, возрасте этих видов. Можно предполагать, что они сохранились в составе флоры Дальнего Востока также с третичного времени. Однако вряд ли эти термофильные и мезофильные элементы могли существовать непрерывно начиная с третичного времени в районах современного их произрастания на территории Монголии. Эти крупнолистоватые термофильные лишайники, характерные представители влажных, тенистых местообитаний (замшелые скалы, поросшие мхами стволы деревьев в лесах), по нашему мнению, не могли выжить как в суровых условиях резко континентального холодного климата плейстоцена, так и в условиях сухого и жаркого климата ксеротермической эпохи. Их иммиграция на территорию МНР, видимо, из восточноазиатского флористического центра могла произойти в одну из теплых и влажных фаз голоцена.
В составе группы "атлантических реликтов" - Arthopyrenia grisea (Schleich,) Koerb., A. rhyponta (Ach.) Massal., Parmelia exasperata DNot., P. glabratula (Lamy) Nyl., Pertusaria henrici (Harm.) Erichs. В экологическом отношении эта группа довольно однородна, она включает лесные эпифитные формы, обитание которых связано главным образом с деревьями лиственных пород. В географическом отношении эти виды отличает та особенность, что их основные ареалы приурочены к Западной Европе, иногда к восточным районам Северной Америки, а по направлению к востоку на Евразиатском материке они имеют "затухающие" ареалы. На территории Азии известны отдельные изолированные местонахождения этих видов, причем наиболее восточные из них приходятся на территорию Алтае-Саяно-Хангайской горной страны.
Так, Arthopyrenia grisea (Schleich.) Koerb. рассеянно встречается по всей территории Европы, где растет в лесах на коре березы, ольхи, рябины и деревьев других лиственных пород (Макаревич, 1977). Ареал этого вида простирается от Фенноскандии, Кольского полуострова, Карелии на севере до Пиренейского полуострова и горных районов Средней Европы на юге и от Британских островов на западе до Польши, Прибалтики, Белоруссии на востоке (Окснер, 1956; Макаревич, 1977). Далее, к востоку, известны изолированные местообитания этого вида - на Украине, в Подольской возвышенности (Тернопольская область) (Окснер, 1956), в Волжско-Камском заповеднике (Голубкова, Малышева, 1980); на территории Азии - в Монголии, Хангае (Бязров и др., 1983). Второй вид этого рода - A. rhyponta (Ach.) Massal., обитающий на гладкой коре деревьев лиственных пород - тополя, ясеня, ольхи, осины, характеризуется более обширным ареалом, охватывающим Европу, Северную Африку, Северную Америку (Массачусетс) (Окснер, 1956). Основная, европейская, часть его ареала простирается от Британских островов, Фенноскандии, Кольского полуострова, захватывая Францию, Бельгию, Швейцарию, Австрию, ФРГ, Польшу, Венгрию, Чехословакию, Румынию, Болгарию, на юге - Пиренейский полуостров и Италию. Восточная граница европейского ареала этого лишайника проходит на европейской части СССР, в Прибалтике, Карпатах. Далее, восточнее, имеются изолированные местонахождения этого вида в Крыму, на Кавказе (Азербайджанская ССР) (Бархалов, 1975), в Татарии, в Волжско-Камском междуречье (Голубкова и др., 1979; Голубкова, Малышева, 1980). После обширной дизъюнкции этот вид встречается в Монголии, где был найден в восточной части Хангая, на высоте 1950 м над ур. м., в травяном лиственничнике на валеже (Бязров и др., 1983). Эти виды артопирений связаны с западноевропейской флорой не только географически, но, видимо, и генетически. Как отмечал А. Н. Окснер (1946 а, 1956), род Arthopyrenia, насчитывающий около 85 видов, развился в умеренных широтах северного полушария, преимущественно в неморальной зоне, особенно в районах лучше сохранивших тургайскую флору, за исключением полутора десятков видов, связанных с тропическими и субтропическими областями земного шара. Областями наибольшего видообразования рода являются Средняя и Атлантическая Европа. Причем среди артопирений немало молодых видов, дифференциация и формирование которых происходили в конце третичного - начале четвертичного периода. Некоторые из артопирений, как отмечал А. Н. Окснер (1946б), несут черты тургайцев, уходящих корнями к тропическим областям. A. grisea (Schleich.) Koerb. и A. rhyponta (Ach.) Massal. - близкородственные виды, относящиеся к секции Polymeridiae Oxn. Вместе с третьим видом - A. persooni Massal. - они образуют один систематический ряд. A. persooni Massal. - вид европейского распространения, основной ареал которого связан с лесами Средней и Атлантической Европы; за пределами этих районов он известен в лесостепных районах Украины и в Закавказье (Азербайджанская ССР). Центральное место в этом ряду, по-видимому, занимает A. rhyponta (Ach.) Massal., который, возможно, является и наиболее древним, о чем свидетельствует его ареал, характеризующийся североатлантическим и средиземноморским разъединением. Обширный дизъюнктивный ареал этого вида, южная часть которого приурочена к средиземноморским районам, дает основание предполагать его доледниковый возраст, а также его возможные генетические связи с тургайской флорой. Два других вида, вероятно, его дериваты. Слабая морфологическая дифференциация этих двух видов как между собой, так и от A. rhyponta (Ach.) Massal. свидетельствует об их молодости.
У видов рода Arthopyrenia, характеризующихся эндофлеодными или слаборазвитыми слоевищами и погруженными в таллом или в субстрат перитециями, прослеживается своеобразная экологическая специализация. Большинство из них, в том числе и рассматриваемые нами виды, растут на гладкой коре деревьев лиственных пород - березы, осины, тополя, ольхи, ясеня, очень редко развиваясь на гниющих остатках растений, перегнойной почве, каменистом субстрате. Таким образом, географическая приуроченность центров их видообразования к районам консервации тургайской флоры, на что указывал А. Н. Окснер (1946б), и наличие в составе этой систематической группы большого количества систематически близких, слабо отграниченных в морфологическом отношении видов позволяют предполагать, что дифференциация и формирование ряда видов этого рода могли произойти в результате распада и переработки древних термофильных тургайских форм в более умеренные холодостойкие формы в конце третичного времени в связи с наступившем похолоданием и в плейстоцене. Специфические экологические особенности артопирений, их связь с мелколиственными древесными породами (березой, осиной) дают возможность предположить наибольший расцвет этой группы лишайников в позднеледниковье (аллеред), когда в результате быстрого и сильного потепления наблюдалось интенсивное развитие березовых лесов на территории Евразии, а также раннего послеледниковья, или времени березы и сосны. Возможно, что именно в конце ледниковой эпохи и в начале послеледникового времени происходили формирование ареалов многих видов этой систематической группы лишайников и их расширение как к северу, так и к востоку. Таким образом, современное географическое распространение A. grisea (Schleich.) Koerb. и A. rhyponta (Ach.) Massal. и генетические связи этих видов позволяют высказать предположение об их иммиграции на территорию Монголии с запада; возможно, это могло произойти в первую половину голоцена.
О проникновении Pertusaria henrici (Harm.) Erichs. на территорию Азии "с западной оконечности Евразиатского материка" также свидетельствует его современный ареал. Как было показано выше, этот вид характеризуется ареалом, приуроченным к листопадным лесам Европы. По направлению к востоку известны лишь отдельные разрозненные местонахождения этого лишайника - в Крыму, на Кавказе, в Алтае, на территории Монголии, где он был найден в Хангае, в верховье р. Хух-Сумэин-Гол и в северо-западной части этой страны, в окрестностях Сагил сомона, в лиственничных лесах на коре лиственницы (Schubert, Klement, 1971; Бязров, 1974а, 1974 б; Бязров и др., 1983).
К этой же группе реликтовых элементов во флоре Монголии мы относим неморальные виды рода Parmelia - P. exasperata DNot. (рис. 48) и P. glabratula (Lamy) Nyl., ареалы которых подробно обсуждались выше. Как было показано, для P. exasperata DNot. и P. glabratula (Lamy) Nyl. характерно североатлантическое разъединение ареалов. Изолированные местонахождения этих видов на территории Азии, в том числе и в МНР, по-видимому, реликтовые. Эти виды относятся к секции Vainioellae (Gyeln.) Essl. подрода Melanoparmelia (Hue) Essl. (Esslinger, 1977), в настоящее время рассматриваемого Эсслингером (Esslinger, 1978) как отдельный род. Ареал подрода Melanoparmelia (Hue) Essl. охватывает северное и южное полушария (Esslinger, 1977). Основная часть ареала этого подрода связана с Голарктикой и циркумполярно охватывает арктические, бореальные и умеренные районы этого региона.
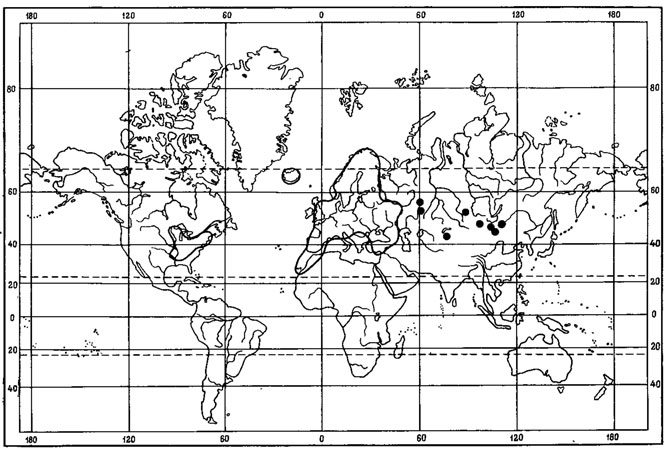
Рис. 48. Распространение на замном шаре Parmelia exasperata DNot. (Ориг.)
Секция Vainioellae занимает центральное место в этом подроде. Из 40 видов подрода Melanoparmelia (Hue) Essl. 29 относятся к этой секции. Как отмечал Эсслингер (Esslinger, 1977), эта группа довольно гомогенна и ее представители отличаются от видов других секций подрода морфологическими и экологическими особенностями. Большинство видов секции Vainioellae - эпифитные лишайники: они обитают на стволах и ветвях деревьев, иногда переходя к обитанию на гниющую древесину, в то время как представители других секций главным образом эпилитные лишайники. Центр современного многообразия секции, как и всего подрода Melanoparmelia (Hue) Essl., находится в Голарктике, где встречается 19 видов из этой систематической группы лишайников, 8 видов - представители австралийской и южноамериканской флоры; ареалы двух видов охватывают северное и южное полушария. В Голарктике наибольшая видовая насыщенность этой систематической группы связана с Северной Америкой: из 29 видов здесь встречается 18. Североамериканская флора наиболее богата эндемиками из этой группы лишайников: 6 видов секции Vainioellae - эндемики ее; один вид, помимо Северной Америки, встречается на Канарских островах. В Европе известно 10 видов из этой группы, из них один вид - эндемик европейской флоры, в Азии - 11 видов, 2 вида - эндемики азиатской флоры. К числу наиболее древних форм этой группы лишайников, видимо, можно отнести фертильные формы, характеризующиеся половым размножением. Анализируя их распространение на основании данных монографии Ахти (Ahti, 1966), посвященной изучению неизидиозных и несоредиозных видов этой группы пармелий, можно видеть, что из 9 видов 7 встречается в Северной Америке, причем из них 4 вида являются эндемиками североамериканской флоры, 3 вида (P. olivacea (L.) Ach., P. septentrionalis (Lynge) Ahti, P. glabra (Schaer.) Nyl) характеризуются широкоголарктическими ареалами, последние 2 вида распределяются между европейской и азиатской флорами. Таким образом, Северная Америка является центром видового разнообразия секции Vainioellae и ее флора наиболее богата эндемичными видами из этой группы лишайников, и, кроме того, здесь концентрируются и наиболее древние типы этой систематической группы лишайников, что позволяет искать исходный центр формирования секции на Североамериканском континенте, причем наиболее богата видами этой секции флора приатлантических районов Северной Америки, т. е. область развития типичных третичных лесов (Ильин, 1941). Североатлантическое разъединение ареалов P. exasperata DNot. и P. glabratuia (Lamy) Nyl. свидетельствует о том, что становление этих видов и их расселение, видимо, происходили до отделения Северной Америки от Европы. О доледниковом возрасте этих видов также говорят южные отрезки их европейских ареалов, связанные с средиземноморскими районами и продолжающиеся в средиземноморские районы Северной Африки. Изолированные местонахождения этих видов в, Монголии и других районах Азии являются свидетельством их более обширных ареалов на Евразиатском материке в прошлом, возможно, во время максимального развития широколиственных лесов, с которыми связано обитание этих видов, во влажную и теплую фазу голоцена (климатического оптимума). "Затухающие" по направлению к востоку ареалы этих пармелий на территории Евразии и отсутствие этих видов в восточных районах Сибири и на Дальнем Востоке позволяют предположить их иммиграцию на территорию Монголии с западной оконечности Евразии.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'