Особенности эндемизма
Флора лишайников Монголии не имеет в своем составе эндемичных семейств и родов. Эндемизм среди лишайников проявляется на уровне вида. Эндемичных видов, ареал которых ограничен территорией Монголии, всего 3 (около 0.4% от общего числа видов) - Aspicilia changaica (Klem.) Golubk., Toninia gobica Golubk., Peltula zabolotnoji (Elenk.) Golubk. (рис. 39, 40). Более многочисленны в составе флоры лишайников МНР субэндемичные виды, ареалы которых ограничены пределами естественных флористических районов. Среди них 18 видов, ареалы которых связаны с территорией Монгольской провинции Центральной Азии, - Rhizoplaca baranowii (Poelt) Golubk., Endopyrenium perminutum (H. Magn.) Golubk., E. mongolicum (H. Magn.) Golubk., E. minutum (H. Magn.) Golubk., Endocarpon mongolicum H. Magn., Psorotichia dispersa H. Magn., Peccania polyspora H. Magn., Collema minutum H. Magn., Peltula minuta (H. Magn.) Golubk., Acarospora suprasendens H. Magn., A. sarcogynoides H. Magn., A. umbrina H. Magn., A. verruculosa H. Magn., Buellia mongolica H. Magn., Toninia olivaceoatra H. Magn., Sarcogyne picea H. Magn., Lecanora subminuta H. Magn., Squamarina kansuensis (H. Magn.) Poelt.
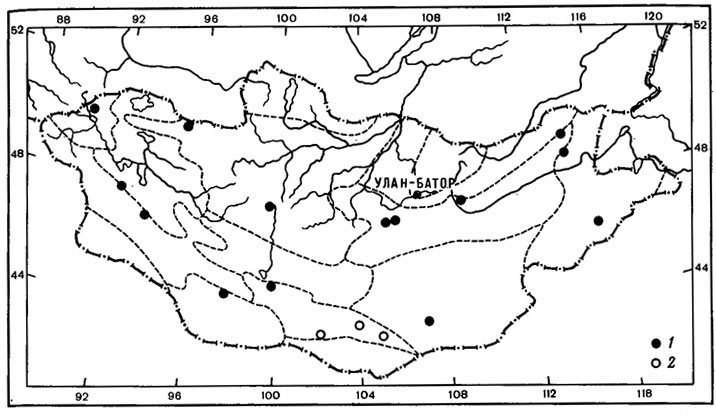
Рис. 39. Распространение на территории Монголии Peltula zabolotnoji (Elenk.) Golubk. (1) и Toninia gobica Golubk. (2). (Ориг.)
К числу субэлементов можно отнести также 5 видов - Acarospora jenisejensis Н. Magn., A. fulva Golubk., A. angusta Н. Magn., Usnea capitata Ras., Evernia terrestris (Tomin) Golubk., распространение которых связано главным образом с территорией Монголии, но они заходят в Забайкалье, Прибайкалье, Туву, Южный Алтай. Общая численность эндемичных и субэндемичных видов лишайников во флоре МНР, таким образом, достигает 27 видов (3.5%).
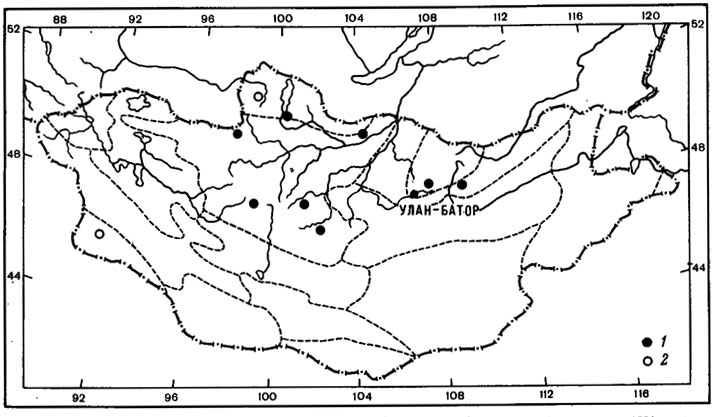
Рис. 40. Распространение на территории Монголии Aspicilia changaica (Klem.) Golubk. (1) и Squamarina pamirica Golubk. (2). (Ориг.)
В. И. Грубов (Грубов, Юнатов, 1952; Грубов, 1959, 1963) неоднократно подчеркивал самостоятельность и самобытность центральноазиатской флоры. Вслед за М. Г. Поповым (1927, 1931) он рассматривал Центральную Азию как особую подобласть Древнего Средиземья, занимающую в основном территорию внутреннего стока Азиатского материка, а именно царство холодных пустынь, пустынных и высокогорных степей. При этом он отмечал высокий родовой и видовой эндемизм среди цветковых растений. В. И. Грубов (1959) указывал, что древнейшей провинцией Центральноазиатской подобласти Древнего Средиземья является Монгольская. Территория Монголии, как свидетельствуют геологические данные, представляла собой сушу уже с верхнего палеозоя (карбон - пермь), начиная с мелового периода здесь установился аридный климат. "Именно здесь мы находим наиболее древние, морфологически своеобразные и систематически обособленные, и даже загадочные "осколки" прошлой пустынной флоры" (Грубов, 1959, с. 14).
Во флоре Центральной Азии известно большое число эндемиков в составе целого ряда систематических групп лишайников (Magnusson, 1940, 1944). Как было показано, 7.2% лихенофлоры МНР составляют центральноазиатские эндемичные виды. Помимо выше перечисленных монгольских эндемичных и субэндемичных видов, во флоре Монголии среди лишайников можно выделить еще 2 группы центральноазиатских эндемиков.
Первая группа видов - виды, распространенные в пределах Монгольской и Тибетской провинций Центральной Азии. Она насчитывает 12 видов - Endopyrenium atrocinereum (Н. Magn.) Golubk., Е. kansuense (Н. Magn.) Golubk., Prosotichia mongolica H. Magn., P. nigra H. Magn., P. kansuensis H. Magn., Lecidea hedinii H. Magn., Buellia hedinii H. Magn., B. sarcogynoides H. Magn., Aspicilia determinata (H. Magn.) Golubk., Lecanora semipallida H. Magn., L. isabellina H. Magn. Вторая группа центрально-азиатских эндемиков включает виды с ареалами, простирающимися в пределах Монгольской, Тибетской и Джунгаро-Туранской провинций Центральной Азии. Группа состоит из 20 видов - Endocarpon sinense Н. Magn., Peltula impressula (H. Magn.) Golubk., Lecidea asiaecentralis H. Magn., Acarospora bohlinii H. Magn., A. gobiensis H. Magn., A. superans H. Magn., Endopyrenium bohlinii (H. Magn.) Golubk., E. inaequale (H. Magn.) Golubk. и др.
Преобладающее число эндемичных центральноазиатских видов, в их числе и эндемики, и субэндемики монгольской флоры, относятся к аридному элементу и, таким образом, являются в экологическом отношении ксерофитными лишайниками. Лишь один субэндемичный байкало-хангайский вид - Usnea capitata Ras. - является лесным мезофильным лишайником, и еще один - хангае-алтайский вид - Evernia terrestris (Tomin) Golubk. - относится к высокогорному элементу. Большинство центрально-азиатских эндемичных видов обитает в пустынно-степных районах центральноазиатской части страны. А. А. Юнатов (1954) писал, что эндемизм среди бореальной флоры Монголии в общем невысок, что нельзя сказать о видах, населяющих большую часть страны, лежащей к югу от мирового водораздела и относимой нами к центральноазиатской подобласти Древнего Средиземья. Та же закономерность в отношении эндемизма проявляется и во флоре лишайников Монголии.
Сделаем попытку выяснить, какова же природа центральноазиатского, и в частности монгольского, эндемизма. Относится ли он по своей природе к консервативному, реликтовому, эндемизму или является молодым, прогрессивным? Во флоре лишайников МНР наиболее богаты центральноазиатскими эндемиками следующие семейства: Acarosporaceae (15 видов), Verrucariaceae (9), Lichinaceae (5), Lecideaceae (4), Heppiaceae, Aspiciliaceae, Physciaceae (по 3 вида каждое), Collemataceae (2), Candelariaceae, Teloschistaceae (по одному виду).
Рассмотрим генетические связи эндемиков центральноазиатской, в том числе и монгольской, лихенофлоры.
Как можно было видеть, наибольшее число центральноазиатских эндемиков приходится на сем. Acarosporaceae (15 видов). Это политипное семейство насчитывает более 400 видов (Poelt, 1973), широко распространенных на земном шаре, но в основном обитание их связано с аридными и горными районами. Большинство представителей акароспоровых лишайников относится к числу пустынно-степных и нагорных ксерофитов. При этом виды этого семейства преимущественно эпилитные лишайники, лишь немногие из них предпочитают в качестве субстрата карбонатные и глинистые почвы, растительные остатки, слоевища лишайников, гниющую древесину и кору деревьев.
В составе лихенофлоры МНР сем. Acarosporaceae насчитывает 57 видов, из них 14 видов относится к числу центральноазиатских эндемиков. В родовом отношении они распределяются следующим образом: 12 видов принадлежат к роду Acarospora, один вид - к роду Polysporina и один вид - к роду Sarcogyne.
В составе рода Acarospora центральноазиатский эндемизм проявляется в различных систематических группах. Из них в монгольской флоре 2 вида - A. fulva Golubk., A. gobiensis Н. Magn. - относятся к подроду Xanthothallia Н. Magn. - желтоокрашенным акароспорам. Остальные виды - Acarospora sarcogynoides Н. Magn., A. suprasedens Н. Magn., A. umbrina Н. Magn., A. verruculosa Н. Magn., A. invadens Н. Magn., A. superans Н. Magn., A. glypholecioides Н. Magn., A. jenisejensis Н. Magn., A. sibirica Н. Magn., A. angusta Н. Magn., A. bohlinii Н. Magn., A. brevilobata Н. Magn. принадлежат к группе коричневоокрашенных акароспор, подроду Acarospora.
Рассмотрим группу желтоокрашенных акароспор и место центральноазиатских эндемичных видов в ее составе. Подрод Xanthothallia насчитывает около 80 видов. Как отмечал Сантессон (Santesson, 1944), желтоокрашенные акароспоры распространены в основном в областях, характеризующихся жарким и засушливым климатом. Лишь немногие виды этой систематической группы лишайников проникают далеко на север. Так, в Арктике встречается лишь один арктовысокогорный вид - Acarospora chlorophana (Wahlenb.) Massal. На земном шаре выделяются два центра видового разнообразия желтоокрашенных акароспор. Один из них находится в районе субтропических пустынь Южной и Юго-Западной Африки - в пустынях Калахари, Намиб и Карро, где обитает 14 эндемичных видов из этой группы лишайников. Вторым очагом видового многообразия желтоокрашенных акароспор являются горно-пустынные районы Северной Америки и Мексика, где также известно 14 эндемичных видов из этой группы лишайников. Для некоторых из них характерны сонорско-древне-средиземноморские ареалы, например для Acarospora heufleriana Koerb. (рис. 41). Богато представлены виды подрода Xanthothallia в районе Средиземноморской области; здесь встречается 17 видов, из них 6 - эндемики флоры этого района. В Азии по направлению с запада на восток происходит постепенное убывание числа видов этой группы лишайников: в Монголии встречается 9 видов, в Средней Азии - 5, в Южной Сибири - 4, на Дальнем Востоке - 2 (Голубкова, 1978, 19806; Голубкова, Шапиро, 1978). Возможно, что лишайники подрода Xanthothallia относятся к числу представителей древней ксерофильной флоры лишайников. Во флоре Монголии эта группа представлена 9 видами (Голубкова, 1980б, 1981). Acarospora fulva Golubk. по своему систематическому положению относится к секции Xanthothallia, объединяющей виды с ареолированными или чешуйчатыми слоевищами. Представители секции проявляют главным образом гондванское распространение, будучи наиболее широко представлены во флорах Южной и Юго-Западной Африки, Австралии, Южной Америки (Magnusson, 1929), в Голарктике - в Средиземноморских районах и южных районах Северной Америки. A. fulva Golubk. генетически наиболее близок к A. xanthophana (Nyl.) Jatta, с которым морфологически очень сходен. Ареал A. xanthophana (Nyl.) Jatta связан главным образом с Новым Светом, где он встречается в горных районах Южной Америки (Боливия, Колумбия, Аргентина, Чили), проникая на севере в южные районы Северной Америки (Аризона), а на юге - в Антарктические районы (Magnusson, 1929). Этот вид также указывается Магнуссоном (Magnusson, 1940) для территории Внутренней Монголии (Китай), хотя это указание вызывает сомнение. Возможно, в этот район проникает A. fulva Golubk.
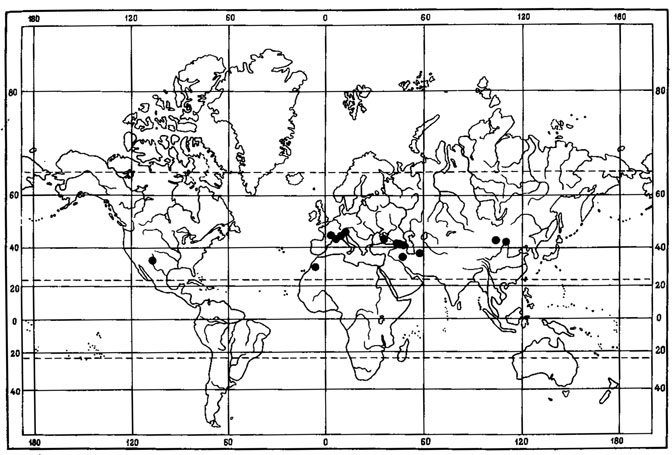
Рис. 41. Распространение на земном шаре Acarospora heufleriana Koerb. (Ориг.)
Ареал Acarospora fulva Golubk. приурочен главным образом к сухим и горным степям Средней Халхи, где он довольно широко распространен, а также к горным степям южного Хангая, долины р. Керулен, Монгольского Алтая; этот вид проявляет инвазии в Прибайкалье (Голубкова, 1979, 1981). Среди видов рода Acarospora этот лишайник является по-своему уникальным.
Как уже было указано выше, представители подрода Xanthothallia и подрода Acarospora различаются главным образом окраской слоевища. Представители первого подрода имеют желтоокрашенные слоевища, второго - коричневоокрашенные. Видимо, ни один подрод не может считаться исходным для другого. Эти группы, возможно, эволюционировали независимо, параллельно друг другу, проявляя сходные черты изменчивости в развитии слоевища и плодовых тел. В том и другом подроде развитие слоевища шло от однообразно накипного (бородавчатого, чешуйчатого или ареолированного) до диморфного (в центральной части слоевища накипного, по периферии - с развитыми лопастями). Возможно, что оба эти подрода произошли от одной вымершей предковой формы, объединяющей признаки этих групп лишайников. Немногочисленные данные по изучению химических веществ в слоевищах этих лишайников показывают, однако, что есть различия и в содержании лишайниковых кислот у представителей этих двух подродов. В слоевищах желтоокрашенных акароспор найдены главным образом вещества шикимово-кислотного происхождения, производные тетроновой кислоты, из которых для них наиболее характерна ризокарповая кислота, реже в слоевищах этих лишайников встречаются другие кислоты, например акараноевая, акареноевая. Характерным веществом коричневоокрашенных акароспор является гирофоровая кислота, которая относится к другой группе лишайниковых веществ - веществам ацетатно-малонатного происхождения, фенолкарбоновым кислотам (Asahina, 1959; Culberson, 1969; Poelt, Steiner, 1971; Голубкова, Шапиро, 1976, 1978; Huneck, 1980, и др.).
Acarospora fulva Golubk. объединяет признаки двух подродов акароспоровых лишайников - содержит в слоевище обе группы лишайниковых веществ (Голубкова, 1979), тем самым проявляя близость к исходным формам этой группы лишайников. Это обстоятельство, а также древние гондванские генетические связи, ареал вида в пределах северной окраины Центральной Азии позволяет высказать предположение, что этот лишайник - один из представителей древней прастепной флоры Монголии.
Субэндемичный вид Acarospora gobiensis Н. Magn. относится к секции Epithallia (Trev.) Zahlbr., представители которой характеризуются диморфными слоевищами. Этот вид принимает заметное участие в образовании лишайниковых эпилитных группировок в пустынно-степных и горных районах юго-западных и центральных областей Монголии, отсутствуя в Восточной Монголии. Он часто встречается по склонам низкогорных хребтов в Восточной, местами в Заалтайской Гоби, а также в Джунгарской Гоби, в Гобийском, Монгольском Алтае, в Котловине Больших Озер, Хангае, Средней Халхе. За пределами МНР ареал этого вида охватывает на территории Китая - Синьцзян, Ганьсу, Цинхай и Внутреннюю Монголию, а в СССР - Тянь-Шань. Севернее из районов Монголии он проникает в Туву, Прибайкалье, Забайкалье. В систематическом отношении он проявляет филогенетические связи с монтанным видом Acarospora oxytona (Ach.) Massal., тяготеющим в своем распространении к средиземноморским районам, а также с арктовысокогорным видом A. chlorophana (Wahlenb.) Massal., образуя с ними один систематический ряд. Довольно слабая морфологическая дифференциация этих видов, их совместное обитание на территории Монголии позволяют предполагать их происхождение от одной исходной предковой формы и их относительную молодость. Возможно, что дифференциация этих горно-ксерофитных форм и их формирование в самостоятельные виды могли происходить на рубеже третичного и четвертичного периодов в связи с изменением физико-географических условий в результате активизации орогенического процесса. A. gobiensis Н. Magn., видимо, можно отнести к числу прогрессивных эндемиков, расширяющих и в настоящее время свой ареал.
В составе группы коричневоокрашенных акароспор 12 видов - центральноазиатские эндемики и субэндемики. Остановимся на некоторых из них.
Заслуживает внимания в этой группе видов Acarospora sarcogynoides Н. Magn., так как относится к наиболее примитивной в филогенетическом отношении в подроде Acarospora группе видов серии Lecideoides Н. Magn. секции Acarospora. Эта группа наиболее близка к роду Polysporina Vezda, эволюционно наиболее примитивному в сем. Acarosporaceae. Как и представители рода Polysporina Vezda, виды группы Lecideoides характеризуются гирозным диском апотециев и сохраняют в онтогенезе древние черты, характерные для графидианового типа - образование в эксципуле и эпигимении черной углистой ткани. Видам этой серии свойственны также примитивные по строению эндолитные слоевища, иногда в виде порошковидной или слабо ареолированной корочки. По-видимому, строение этих лишайников можно рассматривать как один из наиболее примитивных типов среди акароспор, приближающихся к наиболее древнему, исходному типу. Эта группа предковых форм в роде Acarospora по своему составу относится к числу олиготипных, она насчитывает всего 5 видов. Распространение 4 видов этой группы - A. scutula (Stiz.) Hue, A. sarcogynoides Н. Magn., A. tuberculifera Н. Magn., A. gyrosa Golubk. - связано с территорией Древнего Средиземья. A scutula (Stiz.) Hue - эндемик Северной Африки, флоры Алжира (Magnusson, 1929). Три остальных вида - центральноазиатские эндемики. Из них A. sarcogynoides Н. Magn. является монгольским эндемичным видом. На территории МНР этот лишайник - один из характерных представителей флоры степных районов Монголии (Средняя Халха, Восточная Монголия, Прихинганье) (Голубкова, 1981). За пределами МНР этот вид встречается в Китае, во Внутренней Монголии (Magnusson, 1944). A. tuberculifera Н. Magn. обитает в горных районах Ганьсу (Magnusson, 1940) и на Памире (Голубкова, 1973а, 19736), A. gyrosa Golubk. - эндемик Памира (Голубкова, 1972, 1973а). Пятый вид этой группы лишайников - A. tyro- liensis Н. Magn. - эндемик Тирольских Альп, хотя, как отмечал Магнуссон (Magnusson, 1936а), имевший для исследования очень скудный материал, систематическое положение этого вида не совсем ясно.
Таким образом, принадлежность Acarospora sarcogynoides Н. Magn. к группе наиболее примитивных морфотипов рода Acarospora позволяет говорить о его филогенетической древности и отнести его к числу древних элементов монгольской флоры.
В составе центральноазиатских видов коричневоокрашенных акароспор, по-видимому, можно различить и филогенетически молодые виды, например Acarospora suprasedens Н. Magn. и A. invadens Н. Magn., относящиеся к серии Aspicilioides Н. Magn. - группе видов, обозначенной Магнуссоном (Magnusson, 1929) как Mediocres Н. Magn. Она представлена во флоре МНР 13 видами. По обилию видов она довольно многочисленна и насчитывает 70 видов; из них 30 видов ограничены в своем распространении Циркумбореальной областью. Особенно богаты эндемами из этой группы лишайников горные районы Средней Европы. В Древнесредиземноморском подцарстве встречается 24 вида из этой группы лишайников, из них 10 эндемичны для него, а в Сонорском подцарстве - 12 видов, из них 9 эндемиков. В то же время ареал группы выходит далеко за пределы Голарктики: отдельные представители встречаются в Палеотропическом, Неотропическом, Капском и Голантарктическом царствах. Это свидетельствует о древнем происхождении этой группы в целом. Большинство видов (10), присущих флоре Монголии, относится к группе Hypachroeae Н. Magn., которую характеризуют отсутствие на слоевище нижнего корового слоя и бледноокрашенный внутренний слой коры. Среди них можно выделить два систематических ряда - виды с коричневоокрашенными слоевищами, без налета на поверхности, и виды, на поверхности слоевища которых образуется беловато-сероватый налет. Первый систематический ряд образуют виды A. veronensis Massal., A. praeruptarum Н. Magn., A. invadens Н. Magn., A. suprasedens Н. Magn., A. compacta Н. Magn. Центральное место в этом ряду принадлежит A. veronensis Massal. - евритопному лишайнику, обитающему на скалах как гранитных, так и вулканических и карбонатных пород. Ареал этого вида простирается от арктической Европы и Северной Азии до средиземноморских районов Франции (Эро, Вар, о-в Корсика), Италии (Лигурия, Венеция), Греции (о-в Корфу), достигая на юге Северной Африки (Алжир) (Magnusson, 1929). В Азии этот вид встречается, помимо северных районов Сибири, в Казахстане, на Алтае, в Красноярском крае, в Монголии (южный Хэнтэй, Хангай, Хобдоский район, Монгольский Алтай, Котловина Больших Озер, Гобийский Алтай, Восточная Гоби), в Турции, Сирии, Гималаях (Джомолунгма), в Северной Америке, США - в штатах Калифорния и Мэн (Magnusson, 1929; Голубкова, 1978). Такой размах ареала вида, его евротопность и некоторая вариабельность в морфологическом строении - все это позволяет предположить древний возраст этого лишайника. Остальные виды этого систематического ряда морфологически близки к нему и, возможно, его дериваты, о чем свидетельствуют их слабая морфологическая обособленность и обитание в пределах ареала исходной формы.
Для Acarospora suprasedens Н. Magn. и A. invadens Н. Magn. характерно то, что в отличие от близкородственных видов они растут не на обнаженных каменистых субстратах, а развиваются на слоевищах других лишайников. Можно предположить, что этот переход от обитания на каменистой поверхности в более благоприятные условия произошел в результате активизировавшейся аридизации климата, которая проходила на территории Монголии начиная с неогена. A. suprasedens Н. Magn. - монгольский эндемичный вид, описанный с территории Китая - Внутренней Монголии (Magnusson, 1944) и довольно обычный на территории МНР, в степных восточных районах страны (Голубкова, 1981); он найден также в Гобийском Алтае, в хр. Хурху (Голубкова, Цогт, 19746). A. invadens Н. Magn. - более широко распространенный центральноазиатский лишайник. Ареал этого вида охватывает Цинхай, Ганьсу, Внутреннюю Монголию (Magnusson, 1940, 1944), Алашаньскую, Восточную Гоби, Среднюю Халху, на севере достигает Хобдоского бассейна и центральной части Хангая (Schubert, Klement, 1971; Голубкова, 1971а, 1981). В свою очередь эти виды близки к ирано-туранскому лишайнику A. compacta Н. Magn., также встречающемуся на территории МНР, в горах Монгольского и Гобийского Алтая и в юго-западной части Хангая (Schubert, Klement, 1971; Голубкова, 1981). Помимо Монголии, этот лишайник известен на Восточном Памире (Голубкова, 1973а) и в Иране (Szatala, 1957). A. praeruptarum Н. Magn. - монтанный лишайник, встречающийся главным образом в горах Европы, на высоте 300 - 2000 м над ур. м. Его ареал охватывает почти всю Европу, от Скандинавии до средиземноморских районов Франции и от Британских островов на западе до Болгарии, Чехословакии, Польши, Венгрии на востоке. Находки A. praeruptarum Н. Magn. в МНР - в Хангае (хр. Хан-Хухэй), Монгольском Алтае (хр. Сутай-Ула) - первые на территории Азии. Произрастание его на территории Азии свидетельствует о связи флоры горных районов Западной Монголии с монтанной флорой Европы. Возможно, ареал вида недостаточно изучен.
Таким образом, слабая морфологическая дифференциация Acarospora suprasedens Н. Magn. и A. invadens Н. Magn. от близкородственных форм и их обитание в пределах ареалов этих видов позволяют причислить их к числу неоэндемиков.
У центральноазиатского эндемика рода Polysporina - P. sinensis (Н. Magn.) Golubk. - в морфологическом строении просматриваются филогенетически наиболее примитивные признаки. В основной линии филогенеза акароспоровых лишайников прослеживаются черты постепенного изменения в строении репродуктивных органов с целью усиления процесса спорообразования, что проявляется прежде всего в увеличении численности спор в сумках (до 400) и уменьшении их размеров. В отличие от большинства представителей рода P. sinensis (Н. Magn.) Golubk. характеризуется довольно крупными для этой систематической группы спорами и сравнительно небольшим их числом (до 50) в сумках. Этот вид относится к группе ксерофитов: его ареал охватывает пустыню Галбын-Гоби и южную часть Монгольского Алтая; за пределами Монголии: в СССР - Тянь-Шань, в Китае - провинцию Ганьсу. Филогенетически P. sinensis (Н. Magn.) Golubk. наиболее близок к иранскому виду Sarcogyne (Polysporina) polackiana (Mull. Arg.) H. Magn. Примитивные морфологические признаки этого центральноазиатского эндемичного вида, приближающие его к исходным морфотипам рода, а в связи с этим несколько обособленное систематическое положение в группе акароспоровых лишайников, отдаленные иранские генетические связи позволяют предполагать его реликтовую природу.
В роде Sarcogyne к числу монгольских эндемиков относится S. picea Н. Magn. Этот горно-степной лишайник часто встречается на поверхности щебнисто-галечного панциря и скалах в горных и сухих степях Средней Халхи, Восточной Монголии, Прихинганья, проникая к северу в степи Монгольско-Даурского района, Хангая и Прихубсугулья. За пределами МНР его ареал охватывает территорию Внутренней Монголии (Китай). Филогенетически этот вид близок к S. privigna (Ach.) Anzi с широким голарктическим ареалом, также обитающему в степных ценозах Монголии. По-видимому, S. picea Н. Magn. является молодым прогрессивным эндемиком. О филогенетической молодости этого вида свидетельствуют нерезкая морфологическая дифференциация и однородная с исходным видом экология, а также распространение в пределах его ареала.
Таким образом, на основании рассмотренных примеров можно сделать вывод, что среди акароспоровых лишайников различаются древние и самобытные ксерофитные формы, о чем свидетельствует их примитивное морфологическое строение, в котором прослеживаются черты, приближающие эти формы к исходным морфотипам рассматриваемых родов, а также их отдаленные (гондванские) филогенетические связи с представителями пустынных флор других районов земного шара (Южной Африки, Северной и Южной Америки). Но в этой систематической группе также наблюдается в составе лихенофлоры Монголии и молодой, прогрессивный эндемизм.
На втором месте по числу центральноазиатских эндемичных видов в лихенофлоре Монголии стоит сем. Verrucariaceae. Это политипное семейство охватывает более 600 видов, характеризующихся накипными, чешуйчатыми, листоватыми и карликово-кустистыми слоевищами. Представители этих пиренокарповых лишайников широко распространены на земном шаре, главным образом в районах умеренной зоны, где обитают на каменистых субстратах, реже на почве и растительных остатках в горных, арктических и пустынно-степных районах. В составе этого семейства центральноазиатский эндемизм проявляется в родах Endopyrenium и Endocarpon.
Род Endopyrenium насчитывает около 45 видов, распространенных главным образом в Голарктике (Окснер, 1956); в палеотропиках и неотропиках известно всего около 10 видов из этой группы лишайников. Особенно богата видами этого рода средиземноморская область (Zahlbruckner, 1926). В экологическом отношении эти чешуйчатые лишайники преимущественно эпилитные и эпигейные формы, обитающие в горных, степных и пустынных районах, хотя некоторые из них проникают далеко на север, вплоть до арктических районов. В составе монгольской флоры 7 видов этого рода является центральноазиатскими эндемиками, что свидетельствует о том, что Центральная Азия относится к числу районов, с которыми связано видообразование в этой систематической группе лишайников.
Большинство центральноазиатских эндемичных видов относится к секции Endopyrenium, представители которой имеют светло окрашенный эксципул перитециев; среди них Е. atrocinereum (Н. Magn.) Colubk., Е. bohlinii (Н. Magn.) Golubk., E. kansuense (H. Magn.) Golubk., E. minutum (H. Magn.) Golubk., E. mongolicum (H. Magn.) Golubk.; 2 вида - E. inaequale (H. Magn.) Golubk., E. perminutum (H. Magn.) Golubk. - принадлежат к секции Catopyrenium (Stizb.) Oxn. и характеризуются черным эксципулом. Есть одна характерная морфологическая особенность у центральноазиатских видов эндопирениумов - все они принадлежат к формам с примитивным морфологическим строением таллома - в виде бугорчатых, трещиноватых бугорчато-чешуйчатых однообразных корочек. Этот морфологический признак свидетельствует о низкой морфологической организации центральноазиатских эндопирениумов, сближающей их с филогенетически более примитивным родом веррукариевых лишайников - родом Verrucaria. Как известно, более филогенетически продвинутые формы рода Endopyrenium характеризуются чешуйчатыми и даже мелколистоватыми слоевищами. Это обстоятельство дает основание говорить об их достаточной филогенетической древности и близости к исходному типу рода.
На территории Монголии эти лишайники довольно часто встречаются в горных, сухих и пустынных степях, в пустынях по низкогорным хребтам, где обитают на гумусированных участках скал, в их расщелинах, непосредственно на каменистой поверхности скал, иногда на слоевищах других лишайников.
В составе секции Endopyrenium 3 вида - Е. bohlinii (Н. Magn.) Golubk., Е. kansuense (Н. Magn.) Golubk., Е. mongolicum (Н. Magn.) Golubk. - образуют один систематический ряд, являются близкородственными видами и отличаются друг от друга незначительными морфологическими признаками. Центральное место в этом ряду занимает Е. bohlinii (Н. Magn.) Golubk., который относится к числу широко распространенных видов эндопирениумов горных и пустынно-степных районов Южной Монголии. Ареал этого вида охватывает Заалтайскую, Восточную Гоби, Гобийский, Монгольский Алтай, Котловину Больших Озер и Долину Озер; этот лишайник был также найден в Дархатской Котловине в Прихубсугулье. За пределами Монголии он довольно широко встречается в провинции Ганьсу и во Внутренней Монголии на территории Китая, а также в СССР - на Восточном Памире. Как отмечает Магнуссон (Magnusson, 1940), генетически этот вид наиболее близок к пустынному североафриканскому виду Dermatocarpon (Endopyrenium) deserti Zahlbr. Примитивное морфологическое строение этого вида, отдаленные филогенетические связи и обширный ареал в пределах Центральной Азии - все это позволяет отнести его к числу палеоэндемиков и говорить о его связях с первично-пустынными предковыми формами. Два других вида этого систематического ряда - Е. kansuense (Н. Magn.) Golubk., Е. mongolicum (Н. Magn.) Golubk., видимо, дериваты реликтовой формы, о чем свидетельствуют их слабая морфологическая обособленность от исходной формы и обитание в пределах ее ареала (Magnusson, 1940; Schubert, Klement, 1971).
К числу палеоэндемиков, по-видимому, относится и Е. atrocinereum (Н. Magn.) Golubk., также характеризующийся примитивным строением слоевища, в виде толстой потрескавшейся серой корочки, обширным ареалом (Гобийский Алтай, Долина Озер, Ганьсу, Восточный Памир) и филогенетическими связями с североафриканскими пустынными формами. Как указывает Магнуссон (Magnusson, 1940), этот вид наиболее близок^ к североафриканскому виду Dermatocarpon (Endopyrenium) aegyptiacum Zahlbr.
Endopyrenium minutum (H. Magn.) Golubk. является монгольским эндемичным видом; его ареал связан в основном с южными районами Монголии (Джунгарская, Заалтайская Гоби, Гобийский, Монгольский Алтай, Долина Озер), где он встречается в пустынях по низкогорным хребтам, в полупустынях, пустынных и горных степях, обитая на гумусированных участках скал и их основаниях. За пределами Монголии встречается только в Китае в Хэси (Magnusson, 1940). Однако систематическое положение вида не совсем ясно и о характере его эндемизма судить трудно, хотя примитивное строение и ареал его в пределах гобийских районов Монголии позволяют предполагать достаточную древность и близость этого лишайника к первично-пустынным формам.
У представителей секции Catopyrenium (Stizbg.) Oxn. также прослеживаются отдаленные родственные связи с пустынными формами Северной Африки (Magnusson, 1940). Ареал Endopyrenium inaequale (Н. Magn.) Golubk. охватывает Гобийские пустыни (Джунгарская, Заалтайская, Восточная Гоби), Гобийский, Монгольский Алтай, Долину Озер и Среднюю Халху, где этот вид часто встречается в горных, сухих и пустынных степях, в низкогорных хребтах в расщелинах и гумусированных участках скал. Этот лишайник проникает в горные степи Хангая. За пределами МНР он встречается в Китае (Ганьсу) и в СССР - на Восточном Памире (Magnusson, 1940; Голубкова, 19716, 1973а). Как и в случае Е. minutum (Н. Magn.) Golubk, примитивное морфологическое строение этого лишайника, его обширный ареал в пределах Центральной Азии и отдаленные генетические связи свидетельствуют о его филогенетической древности.
Систематическое положение второго центральноазиатского вида этой секции - Endopyrenium perminutum (Н. Magn.) Golubk. - не вполне ясно. Этот вид характеризуется очень крупными спорами, что приближает его, как отмечал Магнуссон (Magnusson, 1940), к видам рода Verrucaria, но в отличие от видов этого рода Е. perminutum (Н. Magn.) Golubk. имеет хорошо развитый коровой слой слоевища, что свойственно видам рода Endopyrenium. Подобными крупными спорами, указывал Магнуссон, характеризуются и некоторые другие виды рода Endopyrenium, например Dermatocarpon (Endopyrenium) trabutii В. de Lesd. из Алжира, Dermatocarpon (Endopyrenium) granulosum В. de Lesd. из штата Нью-Мексико (США), Dermatocarpon (Endopyrenium) verruculosum Miill. Arg. из Ирана. Ареал вида, возможно, недостаточно изучен; он был найден на территории Монголии только в хр. Хурху Гобийского Алтая, где обитал на скалах. За пределами Монголии этот лишайник известен из одного местонахождения в провинции Ганьсу и округа Хэси (Китай) (Magnusson, 1940). Примитивные признаки в строении Е. perminutum (Н. Magn.) Golubk. и его обособленное систематическое положение в составе рода говорят в пользу реликтовой природы этого монгольского эндемика.
В составе рода Endocarpon в монгольской лихенофлоре известны два центральноазиатских эндемичных вида - Е. mongolicum Н. Magn. и Е. sinense Н. Magn. Род Endocarpon насчитывает около 30 видов, из них 2/3 видов связаны в своем распространении с территорией Древнего Средиземья, и лишь 1/3 видов растет за пределами Голарктики, в тропических и субтропических областях Америки, Африки, Австралии (Окснер, 1956). Представители этого рода преимущественно эпигейные и эпилитные лишайники, они обитают в полупустынях, степях и горно-ксеротических местообитаниях. Е. mongolicum Н. Magn. и Е. sinense Н. Magn. часто встречаются на территории Монголии по склонам гор, в равнинных и горных степях, где обитают на почве среди камней, у основания и в расщелинах скал. Ареал Е. mongolicum Н. Magn. охватывает Среднюю Халху, Долину Озер, Гобийский Алтай, Джунгарскую и Заалтайскую Гоби, достигая на востоке Восточной Монголии, а на юге заходя на территории Внутренней Монголии (Китай). Инвазии этого вида отмечены в горных степях Хангая, на севере Дархатской котловины (Прихубсугулье). Е. mongolicum Н. Magn. относится к секции Paracarpidium (Mull. Arg.) Zahlbr., включающей виды эндокарпонов, характеризующихся светлыми перегородками спор. Эта секция относится к числу олиготипных систематических групп лишайников; ее немногочисленные представители встречаются лишь в тропических и субтропических областях земного шара. Обособленное систематическое положение Е. mongolicum Н. Magn. во флоре Монголии, его тропические филогенетические связи свидетельствуют о том, что перед нами один из представителей палеоэндемиков.
Endocarpon sinense Н. Magn. принадлежит к другой секции рода - Endocarpon, которая охватывает большинство видов рода. Представители этой секции характеризуются темноокрашенными перегородками спор. Как отмечал Магнуссон (Magnusson, 1940), этот вид проявляет тесные филогенетические связи с Е. pusillum Hedw. - эпигейным лишайником с широким голарктическим распространением, в том числе и на территории Монголии. Слабая морфологическая дифференциация этих двух видов, обитание Е. sinense Н. Magn. в пределах ареала Е. pusillum Hedw. позволяют отнести его к числу неоэндемиков. Ареал Е. sinense Н. Magn. довольно обширен: помимо территории Монголии (Прихубсугулье, Хангай, Монгольский, Гобийский Алтай, Средняя Халха, Котловина Больших Озер, Долина Озер, Заалтайская Гоби) (Голубкова, 1981; Цогт, 1981), он охватывает округ Хэси и горы Нань-Шань (Китай), а также Центральный Тянь-Шань (Бредкина, 1979, 1981а, 19816).
В составе монгольской флоры имеется 7 видов сем. Lecanoraсеае - центральноазиатские эндемики и субэндемики. В их числе Rhizoplaca baranowii (Poelt) Golubk. - монгольский эндемичный вид; 4 вида рода Lecanora - L. subminuta Н. Magn., L. invadens H. Magn., L. semipallida H. Magn., L. isabellina H. Magn., и 2 вида рода Squamarina - S. kansuensis (H. Magn.) Poelt, S. pamirica Golubk.
Rhizoplaca baranowii (Poelt) Golubk. - неприкрепленный кочующий лишайник с эгагропильной формой слоевища (табл. VI, 1). Ареал этого вида ограничен северо-западной частью Монгольского Алтая, хр. Сайлюгем и нагорьем Сангилен, где он обитает на почве в холоднополынно-типчаковых горных степях. R. baranowii (Poelt) Golubk. относится к олиготипному роду Rhizoplaca, основное ядро которого составляют арктовысокогорные виды с характерной умбиликатной формой накипного слоевища. Генетически R. baranowii (Poelt) Golubk. близок к арктовысокогорному виду R. chrysoleuca (Smith) Zopf (Leuckert е. a., 1976; Голубкова, 1980), широко распространенному не только в высокогорном поясе гор Монголии, но и в горных степях и степных районах страны.
R. baranowii (Poelt) Golubk., видимо, вычленился из популяции R. chrysoleuca (Smith) Zopf, и его отклонение и развитие как особой кочующей формы могли произойти в условиях горных степей под влиянием нарастающей ксерофитизации в периоды усиления континентальности климата в одну из эпох плейстоцена. На молодой эндемизм этого вида указывают его слабая систематическая изоляция и распространение в пределах ареала исходной формы, а также приуроченность узколокального ареала этого лишайника к крайним северо-западным районам современной Монголии, где в начале четвертичного периода шла коренная перестройка физико-географических условий в связи с активностью орогенических процессов.
Род Lecanora по числу видов (около 400) - один из крупнейших (Poelt, 1973). Как отмечал А. Н. Окснер (1946а), этот мезофильный род является заметно односторонним в флорогенетическом отношении, что выражается в преобладающей роли тропического элемента (около 1/3 видов); велика также роль в составе рода неморального элемента. Представители рода обитают на самых разнообразных субстратах, при этом наибольшую группу видов среди леканор образуют лесные эпифитные формы. Центральноазиатские эндемики из этой группы лишайников преимущественно эпилитные лишайники, в том числе и виды монгольской лихенофлоры.
Горно-ксерофитные лишайники Lecanora invadens Н. Magn. и L. semipallida Н. Magn. относятся к секции Lecanora, представители которой характеризуются слабо развитыми накипными слоевищами. Ареал L. invadens Н. Magn. связан главным образом с Тибетской провинцией Центральной Азии, где он встречается в горах Нань-Шаня (Magnusson, 1940) и на территории афганской части Памира (Poelt, Wirth, 1968). В Монголии этот вид относится к числу редких и был найден только в Монгольском Алтае, где обитал в полупустыне на высоте 1950 м над ур. м. (Schubert, Klement, 1971). Генетически этот вид близок к эпилитному, широко распространенному на земном шаре лишайнику L. dispersa (Pers.) Rohl., встречающемуся и на территории Монголии, в степях Средней Халхи. Как и в предыдущем случае, слабая морфологическая обособленность центральноазиатского эндемика от исходной формы, обитание в пределах ее ареала, сходная экология - все это свидетельствует о филогенетической молодости вида. Вероятно, это одна из ксероморфных форм L. dispersa (Pers.) Rohl., вычленившихся из исходной формы под влиянием прогрессирующей аридизации климата, о чем свидетельствует наличие густого налета на поверхности плодовых тел этого лишайника.
Ареал Lecanora semipallida Н. Magn., эпилитного горно-ксерофитного лишайника, также в основном связан с территорией Тибетской провинции - хр. Рихтгофена и некоторые другие районы (Magnusson, 1940), а также с южной частью Монголии (Гобийский Алтай), где он обитает на скалах в горных степях. Как отмечал Магнуссон (Magnusson, 1940), эта эпилитная форма относится к группе видов Lecanora hagenii Ach. и по своим морфологическим особенностям наиболее близка к этому голарктическому виду, встречающемуся и в Монголии, и в других районах Центральной Азии преимущественно на растительных остатках. Слабая морфологическая дифференциация L. semipallida Н. Magn. от других видов группы L. hagenii Ach. и обитание в пределах ареала исходной формы позволяют причислить его к неоэндемикам. Дифференциация этого ксерофитного вида от исходной мезофильной формы и переход к обитанию на каменистый субстрат могли произойти под влиянием прогрессирующей ксерофитизации. О ксероморфных чертах в строении этого лишайника свидетельствует, например, наличие многочисленных гранул, образованных неидентифицированным веществом, на поверхности корового слоя и эпитеция в плодовых телах этого лишайника.
Lecanora subminuta Н. Magn. и L. isabellina Н. Magn. относятся к секции Placodium, объединяющей виды леканор с диморфными талломами. Представители этой секции преимущественно нагорные ксерофитные лишайники, они особенно часто встречаются на обнаженных каменистых субстратах в горных районах, изредка переходя к обитанию на карбонатную почву, мхи и очень редко на древесину. Виды этой систематической группы лишайников особенно широко распространены в пределах территории Древнего Средиземья, от Средиземноморских районов Европы и Африки, Передней Азии до горных районов Средней и Центральной Азии. Некоторые из них встречаются в горных районах Средней Европы, Южной Сибири, отдельные виды проникают в Арктику. Центральноазиатские эндемики - L. subminuta Н. Magn., L. isabellina Н. Magn. - вместе с L. baicalensis Zahblr. и L. kukunorensis H. Magn. - образуют серию азиатских близкородственных горно-ксерофитных видов (Poelt, 1958). Центральное место в ней принадлежит, видимо, L. baicalensis Zahlbr., характеризующемуся обширным ареалом, охватывающим всю территорию Монголии, Забайкалье, Внутреннюю Монголию (Китай), Каракорум, Памир, Узбекистан. Обширный ареал этого вида позволяет предполагать его третичный возраст. Ареал L. kukunorensis Н. Magn. также обширен; он связан главным образом с территорией Монголии (Голубкова, 1981), где этот вид обитает на скалах по склонам гор, в горных и равнинных степях, заходя в высокогорья. За пределами ' МНР он известен в Китае - Цинхай (оз. Кукунор) (Magnusson, 1940), на Памире (Голубкова, 1973а) и в Туркмении (Бредкина, Макарова, 1971). L. subminuta Н. Magn. и L. isabellina Н. Magn. относятся к числу редких лишайников. Первый из них приводится для территории Внутренней Монголии (Китай), где он обитает на известняковых скалах (Magnusson, 1940; Poelt, 1958), а также для территории Монголии (Хангай) (Schubert, Klement, 1971). Второй вид указан для гор Нань-Шань и Внутренней Монголии (Китай) и для южного Хангая, где обитает на скалах (Magnusson, 1940, 1944; Poelt, 1958; Schubert, Klement, 1971). По-видимому, эти виды можно отнести к числу молодых, прогрессивных эндемиков, о чем свидетельствуют их слабая морфологическая дифференциация как друг от друга, так и от других близкородственных видов этой серии, совпадение их ареалов и однородная экология.
Горно-ксерофитный вид Squamarina kansuensis (Н. Magn.) Poelt, ареал которого охватывает Ганьсу (Китай) и Монголию (юго-западный Хангай) (Magnusson, 1940; Poelt, 1958; Schubert, Klement, 1971), где он растет на карбонатной почве, также относится к числу неоэндемиков. Генетически он наиболее близок к S. crassa (Huds.) Poelt (Magnusson, 1940; Poelt, 1958), характеризующемуся обширным древнесредиземноморским ареалом. О молодости центральноазиатского эндемика говорят его слабая морфологическая обособленность от исходной формы, сходная с ней экология и его распространение в пределах ее ареала.
Squamarina pamirica Golubk. - эпигейный лишайник, также относящийся к числу горных ксерофитов. Основной ареал этого вида связан с холодными высокогорными пустынями Восточного Памира, где он играет заметную роль в составе синузий эпигейных лишайников (Голубкова, 1972, 1973а; Бредкина, Голубкова, 1977); встречается также в горных степях Центрального Тянь-Шаня (Бредкина, 1981а) и на территории МНР, в пустыне Джунгарская Гоби (Голубкова, 1981; Цогт, 1981). Изолированный участок ареала этого вида находится в западном Прихубсугулье, в Дархатской Котловине, в бассейне оз. Дод-Нур (Цогт, 1976б) (рис. 40, 2). В составе рода Squamarina этот вид занимает несколько обособленное положение благодаря своеобразному строению апотециев и характеру их образования (Голубкова, 1972). Несмотря на тщательное изучение морфологии этого лишайника и других представителей этого рода, нам не удалось установить его систематического положения среди видов этой систематической группы лишайников. Обособленное систематическое положение Squamarina pamirica Golubk. в составе рода, а также дизъюнктивный ареал его на территории Монголии позволяют отнести этот вид к числу палеоэндемиков.
Таким образом, в составе леканоровых лишайников лихено-флоры Монголии в группе центральноазиатских эндемиков и субэндемиков преобладают неоэндемики, относящиеся к горным ксерофитам. У большинства из них прослеживаются средиземные флорогенетические связи.
Сем. Heppiaceae, как и сем. Acarosporaceae, можно отнести к числу панэмерейских систематических групп лишайников (Wetmore, 1970; Rogers, 1977). Виды сем. Heppiaceae особенно богато представлены во флоре пустынных районов юго-западной части Северной Америки, Мексики, Африки, Центральной Азии и в Средиземноморской области. Из лишайников этого семейства для пустынно-степных районов Центральной Азии характерны виды рода Peltula. В составе флоры этого района известны 5 эндемиков из этой группы лишайников. Во флоре МНР встречается 7 видов из этой группы лишайников, из них 2 вида - Peltula impressula (Н. Magn.) Golubk. и P. minuta (H. Magn.) Golubk. - центрально-азиатские эндемичные виды; один вид - P. zabolotnoji (Elenk.) Golubk. - монгольский эндемик (рис. 39, 1).
Peltula zabolotnoji (Elenk.) Golubk. обитает на корневищах Seliginella involvens, на почве, растительных остатках, на гумусированных участках и в расщелинах скал. Вид часто встречается в степных и пустынных районах Монголии - в Средней Халхе, Восточной Монголии, Котловине Больших Озер, Долине Озер, Восточной и Заалтайской Гоби, изредка в горных степях Монгольского Алтая и Хангая. P. zabolotnoji (Elenk.) Golubk. можно отнести к числу наиболее примитивных типов в этом роде в связи с наличием на нижней стороне чешуек ризоидов, состоящих из одного ряда клеток (Еленкин, 1905). Этот признак приближает этот вид к видам рода Heppia, филогенетически наиболее примитивного в сем. Heppiaceae. Примитивизм в строении P. zabolotnoji (Elenk.) Golubk., в связи с чем этот вид занимает несколько обособленное положение в роде, свидетельствует в пользу его древнего происхождения и близости к исходному типу рода.
В сем. Lecideaceae центральноазиатский эндемизм представлен в родах Toninia и Lecidea.
Род Toninia насчитывает 115 видов, распространенных по всему земному шару, но главным образом в засушливых областях (Окснер, 1968). Во флоре Монголии эта систематическая группа лишайников представлена 8 видами, из них 2 вида - Toninia gobica Golubk. и Т. olivaceoatra Н. Magn. - являются монгольскими эндемиками.
Toninia gobica Golubk. - эндемик восточной окраины Гобийского Алтая (рис. 39, 2; табл. VI, 2) Он обитает на гумусированных участках скал, известен лишь из 3 местонахождений - ущелья Елын-Ама в хр. Гурван-Сайхан, ущелья Алтын-Ама в хр. Хурху и хр. Ноен. Т. gobica Golubk. относится к подроду Thalloedema, в составе которого 30% видов - средиземноморские. Т. gobica Golubk. - эндемик среднеазиатского родства. Он наиболее близок к Т. flavida Tomin, который был описан М. П. Томиным (1950) с Алайского хребта и найден нами на территории Монголии в Котловине Больших Озер. Эти два вида очень сходны по внешнему облику, но отличаются друг от друга рядом морфолого-анатомических признаков и химическими реакциями на действие реактивов (Голубкова, 1973в). Близость этих видов позволяет предположить, что, видимо, они произошли при дифференциации на обширном ареале единого предка.
Другой монгольский эндемичный вид - Toninia olivaceoatra Н. Magn. относится к секции Toninia, представители которой характеризуются плоскими или слабо выпуклыми слоевищными чешуйками и поперечно-многоклеточными спорами. Т. olivaceoatra Н. Magn. является характерным лишайником степей Восточной Монголии и Прихинганья, где довольно часто встречается на поверхности силикатных и базальтовых скал, в их расщелинах и на гумусированных участках. Западная граница его ареала проходит в восточных районах Средней Халхи и юго-восточных хребтах Гобийского Алтая (Голубкова, 1981). За пределами МНР этот вид известен на территории Китая - Внутренняя Монголия (Magnusson, 1944). Он проявляет филогенетические связи с европейским горно-ксерофитным видом Т. cinereovirens Massal., который произрастает в основном в средиземноморских районах и горах Средней Европы, проникая к востоку до Крыма, Куйбышевской и Ульяновской областей и Южного Урала, где он обитает на сухих и хорошо прогреваемых участках (Окснер, 1968; Poelt, 1969). Морфологическая близость этих двух видов и однородность их экологии говорят о том, что они могли произойти при дифференциации на обширном ареале единой миоцен-плиоценовой предковой степной формы, распад ареала которой мог произойти уже в четвертичный период в связи с похолоданием климата и физико-географическими перестройками.
Что касается других видов лецидиевых лишайников - Lecidea hedinii Н. Magn., L. subconcava H. Magn., характеризующихся центральноазиатским распространением, то их систематическое положение еще не совсем ясно.
В составе лихенофлоры Монголии 4 эндемичных и субэндемичных видов - Aspicilia changaica (Klem.) Golubk., A. mikhnoi (Zahlbr.) Oxn., Usnea capitata Ras., Evernia terrestris (Tomin) Golubk. - занимают несколько обособленное положение, так как распространение их связано с районами байкало-саяно-хангае-алтайского горного сооружения.
Aspicilia changaica (Klem.) Golubk. - эпилитный лишайник, эндемик горных степей Северной Монголии, с ареалом в пределах районов Прихубсугулья, Хэнтэя, Хангая (рис. 40, 1). Изредка этот лишайник встречается в высокогорном поясе и на скалах в лиственничных лесах. Вид принадлежит к политипному роду Aspicilia, обильно представленному во флоре Монголии. Виды этого рода принимают заметное участие в растительном покрове всего региона. A. changaica (Klem.) Golubk. генетически близок к A. transbaicalica Oxn. (Klement, 1965), также встречающемуся в горных степях Северной Монголии. Эти виды очень близки и отграничены друг от друга незначительными морфолого-анатомическими признаками. Оба вида отличаются от других аспицилий карликово-кустистой формой роста накипного слоевища (в виде вертикально направленных выростов), сформировавшейся, видимо, в условиях степей в связи с обитанием на плоских горизонтальных поверхностях скал, едва выступающих над уровнем почвы. Эти виды образуют один систематический ряд с арктовысокогорный видом A. mastrucata (Wahlenb.) Ach. и высокогорным Lecanora (Aspicilia) nunatakkorum Poelt (Klement, 1965). Aspicilia changaica (Klem.) Golubk. можно отнести к числу молодых прогрессивных эндемиков, о чем свидетельствует его слабая морфологическая обособленность от A. transbaicalica Oxn. и распространение его в пределах ареала этого вида. Можно полагать, что дифференциация и оформление этой горно-ксерофитной формы могли произойти в горных районах современной Северной Монголии в криоксеротических условиях позднего плейстоцена-раннего голоцена.
Субэндемичный вид Aspicilia mikhnoi (Zahlbr.) Oxn. характеризуется забайкальско-хангайским ареалом. В Хангае он является сравнительно редким видом, растущим на выходах горных пород в степных и высокогорнолуговых сообществах на склонах и вершинах сопок, гребнях хребтов (Бязров и др., 1983). За пределами МНР обитает в южном Забайкалье, в степях р. Ага, где встречается, по свидетельству Цальбрукнера (Zahlbruckner, 1911), довольно часто. Филогенетически этот вид наиболее близок к гипо-арктомонтанному евразиатскому лишайнику Aspicilia cupreoatra (Nyl.) Arnold (Zahlbruckner, 1911). Этот гипоарктомонтанный вид часто, встречается в горных и равнинных степях Монгольско-Даурского района и Восточной Монголии, где входит в число доминантов в составе эпилитных лишайниковых группировок. Нечеткая морфологическая дифференциация Aspicilia mikhnoi (Zahlbr.) Oxn. от филогенетически наиболее близкой формы, однородная с ней экология и обитание в пределах ее ареала свидетельствуют о молодости этого субэндемичного вида.
Субэндемик Usnea capitata Ras. характеризуется прибайкаль- ско-хангайским ареалом. В Хангае он встречается довольно редко, обитая на затененных скалах, валунах под пологом леса в средне- горном поясе (Бязров и др., 1983). В районе оз. Байкал этот вид обитает по скалистым горам западного побережья, в проливе Ольхонские ворота (Рассадина, 1936). В систематическом отношении Usnea capitata Ras. принадлежит к подсекции Comosae Mot. секции Barbatae Mot., группе видов Usnea glabrescens (Nyl.) Vain. Представители этой секции главным образом лесные эпифитные формы, широко распространенные в лесной зоне и горных лесах Голарктики. Нечеткая морфологическая дифференциация видов группы Usnea glabrescens (Nyl.) Vain., совпадение их ареалов свидетельствует об их филогенетической молодости. Usnea capitata Ras. является одной из эпилитных форм в составе этих лесных эпифитных лишайников. Возможно, ее обособление и оформление в самостоятельный вид произошли в результате перехода к обитанию на новый субстрат.
Evernia terrestris (Tomin) Golubk. относится к роду Evernia Ach. ex Luyken, который A. H. Окснер (1946a) относил к числу автохтонных тургайских родов, в дальнейшем подвергшегося бореализации. В составе его 13 видов (Флоровская, 1952), распространенных преимущественно в Голарктике; лишь один из них встречается в Южной Америке (Чили). Из них 8 видов произрастают в Азии, 6 - в Восточной Азии. Во флоре Монголии род Evernia Ach. ex Luyken представлен 6 видами - Е. esorediosa (Mull. Arg.) DR., E. perfragilis Llano, E. terrestris (Tomin) Golubk., E. mesomorpha Nyl., E. divaricata (L.) Ach., E. arenaria (Retz.) Fr. Первые 4 вида, в их числе и Е. perfragilis Llano, - близкородственные виды, представители секции Archevernia Th. Fr.; они образуют один систематический ряд - Esorediosa Flor. (Флоровская, 1952). Центральное место в этом ряду занимает Е. esorediosa (Miill. Arg.) DR. - лесной эпифитный восточно-азиатский вид, который, по мнению Е. Ф. Флоровской, является одним из древних в роде. Возникновение исходных форм этого вида она относит к верхнему мелу. Об этом свидетельствует как ряд примитивных признаков в анатомо-морфологическом строении вида, так и образование апотециев, т. е. более примитивный способ размножения - с помощью спор, нехарактерный для представителей этой группы кустистых лишайников, в большинстве размножающихся вегетативно - соредиями, изидиями или фрагментами слоевища. На территории Монголии этот восточноазиатский лишайник в районе Прихубсугулья имеет западную границу своего распространения. Три других вида этого систематического ряда - Е. mesomorpha Nyl., Е. perfragilis Llano, Е. terrestris (Tomin) Golubk. - дериваты этой исходной тургайской формы, обособление которых, видимо, произошло в конце плиоцена - начале плейстоцена в связи с наступившим общим похолоданием климата. Как полагал Н. А. Миняев (1940), например, бореальный эпифитный вид Е. mesomorpha Nyl., морфологически очень близкий к Е. esorediosa (Miill. Arg.) DR., но размножающийся вегетативно - соредиями и изидиями, видимо, сформировался лишь в начале четвертичного периода. Два других вида - Е. terrestris (Tomin) Golubk., Е. perfragilis Llano - криоксерофитные кочующие лишайники, являются примерами форм, которые прошли этап ксерофитизации из мезофильных элементов. Их обособление от исходной мезофильной формы, возможно, произошло в периферической части ее ареала в начале плейстоцена, во время деградации древних лесных третичных типов, и их процесс формирования в самостоятельные виды проходил в условиях криофитных степей плейстоцена в горных районах Северной Монголии и Южной Сибири. О молодости этих видов свидетельствует слабая морфологическая обособленность как от исходной формы, так и друг от друга. Под влиянием бореализации Е. terrestris (Tomin) Golubk. перешел к обитанию на замшелую почву и тем самым освоил новую экологическую нишу, проник за пределы лесного пояса в подгольцовый и высокогорный пояса. При этом он утратил способность к половому размножению, приобретя способность к вегетативному размножению с помощью фрагментов слоевища. В Монголии Е. terrestris (Tomin) Golubk. был собран в высокогорных злаковотипчаковых степях в окрестности оз. Хух-Нур на Хангае, в хр. Хасагт-Хайрхан-Ула Монгольского Алтая, в злаковых горных степях на высоте 2970 м над ур. м.; он известен также из окрестностей оз. Холбо-Нур (Томин, 1933), откуда был описан как форма Е. esorediosa (Mull. Arg.) DR. Западнее этот вид был отмечен на Алтае (Рассадина, 1938), где он обитал на ледниковых моренах и в альпийской тундре в истоках р. Канаса, а также в Казахстане в долине р. Каджар (Томин, 1933).
Таким образом, на основе анализа эндемизма лишайников МНР можно сделать следующие выводы. Прежде всего необходимо отметить, что собственно эндемы лихенофлоры МНР имеют очень небольшой удельный вес, составляя всего 0.5% состава лихенофлоры. Более заметна во флоре лишайников МНР роль субэндемиков (3.5% состава лихенофлоры), среди которых значительна доля монгольских эндемиков (2.3% состава флоры). Центральноазиатские эндемики охватывают 7.2% ее состава.
Среди эндемиков и субэндемиков лихенофлоры МНР и центральноазиатских эндемов преобладают пустынно-степные, горностепные и горно-ксерофитные формы, приуроченные главным образом к центральноазиатским районам МНР, что свидетельствует о том, что аридная лихенофлора Монголии, а также в целом Центральной Азии достаточно самобытна.
Большинство центральноазиатских эндемиков проявляет филогенетические связи с представителями флор территории Древнего Средиземья, из чего можно сделать вывод об их принадлежности к средиземной группе флорогенетических элементов. В их составе можно выделить древние и самобытные ксерофитные формы - Acarospora fulva Golubk., A. sarcogynoides Н. Magn., Peltula zabolotnoji (Elenk.) Golubk., Polysporina sinensis H. Magn., - которые, видимо, должны рассматриваться как реликтовые, находящиеся в условиях консервации благодаря благоприятному для них пустынно-степному режиму южных районов Монголии. Об этом свидетельствует их примитивное морфологическое строение, в котором прослеживаются черты, приближающие эти формы к исходному морфотипу рода, их отдаленные филогенетические связи с представителями пустынных флор других районов земного шара (Южной Африки, Северной и Южной Америки), нередко обособленное систематическое положение в составе рода, а также экологические особенности - обитание в пустынно-степных ценозах Монголии. Все это дает основание предполагать их филогенетические связи с древними первичноксерофитными пустынными формами, с ксерофитно-пустынными гондванскими и древнесредиземноморскими элементами. Некоторые из них в условиях прогрессирующей аридности климата Центральной Азии дали начало новым циклам форм, главным образом из числа горных ксерофитов, - Acarospora gobiensis Н. Magn., Sarcogyne picea H. Magn., Endocarpon sinense H. Magn., Endopyrenium mongoli- cum (H. Magn.) Golubk. и др., которые могут быть отнесены к числу молодых прогрессивных эндемиков.
Довольно слабо в лихенофлоре МНР представлен автохтонный монгольско-даурский горно-степной элемент - Aspicilia changaica (Klem.) Golubk. и A. mikhnoi (Zahlbr.) Охn. Горно-степной монгольско-даурский эндемизм является молодым, прогрессивным, о чем свидетельствует слабая обособленность этих эндемичных форм от близкородственных видов, обитающих совместно с ними в аналогичных экологических условиях бореальной части МНР. Филогенетические связи монгольско-даурских эндемиков с арктовысокогорными и гипоарктомонтанными формами позволяют отнести их к ксерофилизованным дериватам бореальной (голарктической) флоры и предполагать их возможные связи с элементами плейстоценового флористического комплекса.
В составе субэндемиков различаются также формы, которые можно причислить к группе криоксерофилизованных дериватов мезофильных лесных флор (Evernia terrestris (Tomin) Golubk.). Об этом свидетельствуют филогенетические связи этих криоксерофитных высокогорных хангайско-алтайских форм с неморально- лесными формами (Evernia esorediosa (Mull. Arg.) DR.).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'