Географические элементы лихенофлоры
Основной единицей географического анализа флоры лишайников МНР служит зональный географический элемент флоры, который выделяется в зависимости от растительно-климатической зоны, где этот элемент наиболее распространен. В отечественной лихенологии большинство классификационных систем географических элементов флоры основано на зональном принципе (Макаревич, 1963; Трасс, 1970; Окснер, 1974; Бархалов, 1975; Питеранс, 1982, и др.). Как отмечал крупнейший советский биогеограф А. Н. Окснер (1974), изучение распространения лишайников показывает, что формирование большинства их ареалов тесно связано с определенными растительно-климатическими зонами. Лишайники вследствие большей, чем у цветковых растений, ригидности генотипической структуры, как организмы, которые образуют, видимо, в меньшем количестве и более медленно новые биотипы, значительно лучше по сравнению с цветковыми растениями сохраняют свою первоначальную связь с растительно-климатическими зонами (Окснер, 1948). А. Н. Окснер указывал, что этим лишайники в некоторой степени отличаются от цветковых растений, у которых связь ареалов с растительно-климатическими зонами менее заметна вследствие более энергичного видообразования, более быстрой и резкой реакции их на условия среды как в минувшие эпохи, так и в настоящее время, в результате чего они гораздо чаще нарушают границы определенных зон.
М. Ф. Макаревич (1968) подчеркивала, что при рассмотрении современных ареалов растений в пределах растительно-климатических зон мы не отрываем растение от условий его существования, поскольку каждая растительно-климатическая зона характеризуется строго определенным комплексом экологических факторов, во взаимодействии с которыми происходят развитие и расселение растительных организмов. Зональное распределение растений на земном шаре - явление историческое, поэтому современные ареалы растений в пределах определенных растительно-климатичеческих зон в известной степени отражают их исторические связи.
Во флоре лишайников Монголии нами выделены следующие зональные географические элементы - бореальный, неморальный, аридный. Кроме элементов с выраженным зональным характером обобщенного ареала, выделяются географические элементы с азональным распространением - арктовысокогорный, гипоарктомонтанный, монтанный, высокогорный, субокеанический и мультизональный.
Нами не выделяется мультирегиональный (плюрирегиональный) элемент, как это принималось в некоторых классификационных системах при анализе лихенофлор Памира, горного Зеравшана, лихенофлоры Латвии (Голубкова, 1977; Кудратов, 1979; Питеранс, 1982). Этот элемент в отличие от остальных географических элементов флоры выделялся не на зональной, а на региональной основе. Как отмечал А. Н. Окснер (1974), уже одни формальные основания - особенности строения ареалов, а также соображения общего ботанико-географического порядка, история миграции видов - заставляют вычленить в особый элемент виды, встречающиеся на трех континентах и более. Однако в данном случае, видимо, можно согласиться с X. X. Трассом (1970), который указывал, что поскольку многие виды мультирегионального элемента имеют центры массового развития именно в Голарктике в сочетании с хорошо выраженным зональным распространением, а кроме того, так как наши знания в отношении распространения многих видов лишайников вне Голарктики явно недостаточны, в целях анализа локальной флоры правильнее рассматривать их как принадлежащие к тому или иному зональному элементу и при этом выделять их в особый субэлемент. Взгляды X. X. Трасса впоследствии разделили М. Ф. Макаревич и другие исследователи (Макаревич, 1971а; Кудратов, 1979).
При детализации классификации ареалов, как и в других системах (Макаревич, 1968; Окснер, 1974; Бархалов, 1975, и др.), нами используются особенности широтно-регионального распространения лишайников, на основе чего выделяются типы ареалов. Внесены некоторые изменения в названия элементов. Так, вместо "арктоальпийский" и "альпийский" элементы соответственно используются - арктовысокогорный и высокогорный. Это связано с тем, что высокогорная растительность Монголии представлена преимущественно кобрезиевниками - особым типом высокогорной растительности, свойственной горным массивам внутриконтинентальных районов Евразии. Состав кобрезиевников, условия их существования и развития значительно отличаются от типично альпийской растительности в горнопоясных системах океанического типа, и поэтому по отношению к ним нецелесообразно использовать такие понятия, как "альпийская растительность" (Карамышева, Банзрагч, 1976). В отличие от вышеупомянутых систем географических элементов ксеромеридиональный элемент мы называем аридным. Нами также введен мультизональный элемент.
Остановимся на характеристике географических элементов флоры лишайников Монголии.
I. Арктовысокогорный элемент. Объединяет 108 видов (14.3%). Основное распространение видов связано с Арктикой, иногда с Антарктикой, и с высокогорьями земного шара. В составе этого элемента представители 21 семейства лишайников. Наибольшее число арктовысокогорных видов относится к семействам Lecideaceae (12 видов), Aspiciliaceae (12), Parmeliaceae (10), Umbilicariaceae, Usneaceae и Lecanoraceae (по 8 каждое), Peltigeraceae (6 видов).
Ареал большинства арктовысокогорных лишайников ограничен на территории Монголии высокогорными районами; эти виды встречаются фрагментарно в высокогорном поясе Монгольского Алтая, Хэнтэя, Хангая, а также в Прихубсугулье. Отдельные виды, например Thamnolia vermicularis (Sw.) Ach., Dactylina madreporiformis (Sw.) Ach., Umbilicaria virginis Schaer., по горным хребтам Монгольского Алтая проникают далеко на юг в высокогорные степи северо-западных хребтов Гобийского Алтая. В высокогорном поясе арктовысокогорные лишайники принимают значительное участие в растительном покрове, играя заметную роль в составе кобрезиевников, высокогорных степей, горных тундр, в петрофитных сообществах. Однако некоторые арктовысокогорные виды, например Phaeophyscia constipata (Norrl. et Nyl.) Moberg, Hypogymnia subobscura (Vain.) Poelt, обитают в лесных и степных ценозах и по каменистым склонам нижне- и среднегорного поясов, видимо, являясь в составе лесной и горно-степной флоры Монголии гляциальными реликтами.
Господствующее положение среди лишайников арктовысоко- горного элемента занимают плагиотропные жизненные формы, причем наибольшее число их принадлежит к накипным (60% от общего числа видов), и только 26% - к листоватым. Ортотропные жизненные формы охватывают всего 14% всего состава экобио-морф. Характерно также наличие в составе экобиоморф свободно-живущих жизненных форм, представленных такими видами, как Evernia perfragilis Llano, Dactylina madreporiformis (Sw.) Ach., Cornicularia divergens Ach.
А. Голарктический арктовысокогорный субэлемент. Объединяет 71 вид; ареалы этих лишайников связаны с арктическими и высокогорными районами Голарктики.
1. Чукотско-монгольский тип ареала характерен для 2 видов - Aspicilia peripterodes (Nyl.) Oxn. и Arthopyrenia humicola Vain. О распространении этих лишайников имеется мало данных. До находок Aspicilia peripterodes (Nyl.) Oxn. в Монголии, в горной степи на перевале хребта Тургэн-Ула (Schubert, KJement, 1971), он был известен лишь на Чукотском полуострове, на побережье Кониамского залива (Nylander, 1888). Arthopyrenia humicola Vain, был описан Вайнио (Vainio, 1909) по образцам, собранным на северном побережье Чукотского полуострова, и до сих пор этот вид был известен только из классического местонахождения. В Монголии этот лишайник встречен однажды в Хангае, южнее Тэвшрулэха, на высоте 2390 м над ур. м., где он обитает на почве и мхах в высокогорных кобрезиевых лугах (Бязров и др., 1983).
2. Таймырско-монгольский тип ареала присущ одному виду - Aspicilia fimbriata (Н. Magn.) Охп. Ареал этого вида также изучен недостаточно. A. fimbriata (Н. Magn.) Охп. известен был лишь на п-ове Таймыр, мысе Челюскин (Окснер, 19716). В Монголии был найден в Хангае, в петрофитной степи, на гребне хребта, на высоте 1700 м над ур. м., северо-восточнее госхоза Тэвшрулэх (Бязров и др., 1983).
3. Американо-азиатский тип ареала характерен для 4 видов - Asahinea chrysantha (Tuck.) Culb. et Culb., Umbilicaria caroliniana Tuck., Evernia perfragilis Llano, Cetraria laevigata Rassad.; основное распространение этих видов связано с арктическими и высокогорными районами Азии и Северной Америки.
Виды с подобным распространением, в том числе и выше указанные, отнесены Крог (Krog, 1968) к азиатско-североамериканскому элементу, к амфиберенгийской группе видов, включающей арктоальпийские растения с ареалами, простирающимися по обе стороны от Берингова моря и Берингова пролива. При этом, как отмечала Крог, одни из этих видов имеют ограниченные ареалы, охватывающие, например, острова Берингова пролива, ареалы других простираются от бассейна р. Лены на западе до бассейна р. Маккензи на востоке, в то время как третьи широко распространены от Уральских гор до Девисова пролива. Кроме того, некоторые виды могут иметь изолированные местонахождения в европейской Арктике или западной Гренландии.
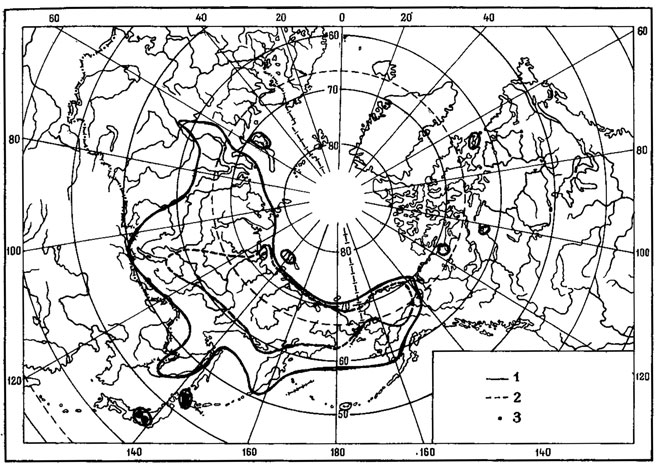
Рис. 6. Распространение на земном шаре видов рода Asahinea. (По: Culberson, Culberson, 1965). 1 - A. chrysantha (Tuck.) Culb. et Culb.; 2 - A. schojanderi (Llano) Culb. et Culb.; 3 - A. kurodakensis (Asah.) Culb. et Culb
Примером амфиберенгийского распространения может служить как ареал. Asahinea chrysantha (Tuck.) Culb. et Culb. (рис.6), так и ареал всего олиготипного рода Asahinea, охватывающий Тихоокеанское побережье и сибирские районы Азии, а также северо-западные районы Северной Америки. Основное распространение А. chrysantha (Tuck.) Culb. et Culb. связано с арктическими и горными районами Сибири, где этот вид встречается от Урала до Камчатки и Берингова пролива и от Ново-Сибирских островов и берегов Северного Ледовитого океана до Южной Сибири и Монголии, хребтов Сихотэ-Алиня и Приморского края, обитая на почве, среди лишайников и мхов в северных и горных тундрах, на каменистых россыпях, в лиственничном редколесье. В высокогорных районах Японии, на островах Хоккайдо и Хонсю, находится самое южное местонахождение этого вида на горе Фудзияма (Окснер, Рассадина, 1960; Culberson, Culberson, 1965). На территории МНР этот лишайник встречается лишь в районе Прихубсугулья и в Хэнтэе (Савич, 1924; Цогт, 1976а, 1981; Голубкова, 1981), где обитает на почве в высокогорном поясе, изредка в лиственничных лесах. Причем его местонахождение в Южном Хэнтэе, в окрестностях г. Улан-Батора, на горе Богдо-Ула - одно из самых южных в Восточной Сибири, находится на пределе лесной растительности, на границе с зональными степями Монголии. В арктических районах A. chrysantha (Tuck.) Culb. et Culb. дает инвазии в европейскую Арктику, встречаясь спорадически на о-вах Новая Земля, Кольском полуострове, в Скандинавии. В Северной Америке распространение этого лишайника связано с Аляской, Алеутскими островами и северными районами Канады. Он широко встречается на Аляске, на ее северном побережье, на п-ове Сьюард, в хр. Брукс, в бассейне р. Юкон, на Аляскинском хребте и Тихоокеанском побережье Аляски (Culberson, Culberson, 1965; Krog, 1968; Moser е. a., 1979; Thomson, 1979). В Канаде этот вид обычен в бассейне р. Маккензи, в долине р. Пил и Артик-Ред-Ривер (Bird е. а., 1980); в районе Северо-Западных Территорий он встречается в долинах рек Коппермайн (Thomson, 1970) и Телон (Scotter, Thomson, 1966).
Как отмечал Кернефелт (Karnefelt, 1979), Cetraria laevigata Rassad. относится к числу арктоальпийских тундровых видов с обширным амфиберенгийским ареалом. Современный центр распространения этого вида расположен в беренгийском районе. Так, на Аляске, например, С. laevigata Rassad. является одним из обычнейших напочвенных кустистых лишайников, далее к востоку от дельты р. Маккензи и вдоль арктического берега Канады его встречаемость постепенно снижается, и в районе залива Св. Лаврентия этот вид очень редок, на западе о-ва Ньюфаундленд и в юго-западной Гренландии известно всего несколько местонахождений этого лишайника. Южнее имеются изолированные местообитания С. laevigata Rassad. высоко в горах на территории США - в Белых горах в штате Нью-Гэмпшир, в штатах Нью-Йорк, Вермонт, Мэн, Северная Каролина. На территории Азии ареал этого лишайника охватывает арктическую Азию, от Чукотского полуострова и о-ва Врангеля до Урала (Рассадина, 1943, 1950; Llano, 1950). При этом центр массовости в развитии этого вида в арктической Азии, видимо, также приурочен к беренгийскому району: в районе Чукотки С. laevigata Rassad. относится к числу часто встречающихся видов (Андреев, 1979; Макарова, 1979а), в то время как на Таймыре этот лишайник встречается редко (Пийн, 1979а, 1979б). Южнее С. laevigata Rassad. распространен в горах Восточной и Южной Сибири, Монголии, Дальнего Востока, Камчатки, о-ва Сахалин, Японии, Северного Китая, п-ова Корея, Северо-Западной Индии и Непала (Рассадина, 1950; Karnefelt, 1979]. На территории Монголии этот лишайник встречается в Прихубсугулье, Хэнтэе, Хангае, Монгольском Алтае, где он обитает в высокогорном поясе, на кобрезиевых пустошах, реже в лиственничных лесах, в ерниках. В западной части ареала С. laevigata Rassad. его инвазии обнаруживаются в восточных районах Европы (Архангельская область) (Рассадина, 1943, 1950). К. А. Рассадина (1943, 1950) полагает, что в Европе этот вид замещается С. ericetorum Opiz. Однако Кернефелт (Karnefelt, 1979) утверждал, что это не совсем так, ибо С. ericetorum Opiz ассоциирует с хвойными лесами, а С. laevigata Rassad. относится к тундровым видам. Он отмечал также, что общее распространение С. laevigata Rassad. позволяет предположить, что этот вид имел немного более широкое распространение до плейстоцена, о чем свидетельствуют его изолированные местонахождения в Гималаях и в Новой Англии (США). Северосибирская и основная североамериканская популяции, вероятно, пережили плейстоценовые оледенения в беренгийском районе, где существовали обширные, свободные ото льда рефугиумы, в частности на Аляске.
Umbilicaria caroiiniana Tuck, характеризуется обширным дизъюнктивным ареалом, охватывающим горные районы Восточной Азии, арктические и горные районы Северной Америки. На территории Азии этот лишайник встречается в Японии, где обитает в горах на высоте 3000 м над ур. м. (Llano, 1950), на Камчатке (Городков, 1935), в Амурской области - р. Зея (Llano, 1950), в Становом хребте (Еленкин, Савич, 1911; Suza, 1925), на западном побережье оз. Байкал (Рассадина, 1936), в гольцовом поясе Западного Саяна и Кузнецкого нагорья (Седельникова, Седельников, 1979), в Монголии (Хангай) (Бязров и др., 1983). Вторая часть ареала этого лишайника связана с Аляской, где он приводится для п-ова Сьюард, мыса Барроу, хр. Брукс, бассейна р. Юкон и других районов, а также с Северо-Западной Канадой (долина р. Пил) (Llano, 1950; Krog, 1968; Moser е. а., 1979; Thomson, 1979; Bird е. а., 1980). Третья часть ареала Umbilicaria caroliniana Tuck, приурочена к восточным районам США, штатам Теннесси и Северная Каролина, где этот лишайник довольно часто встречается в горах Аппалачи (Llano, 1950; Bird е. а., 1980). На территории Монголии U. caroliniana Tuck, является редким лишайником и был собран лишь однажды в Хангае, в дол. р. Цэцэрлэг, на высоте 1850 м над ур. м., на выходах горных пород в лиственничном лесу (Бязров, 1983).
Американские ученые (Bird е. а., 1980) характеризуют ареал этого вида как амфиберенгийско-западноарктическо-аппалачский. Они считают, что его дизъюнктивный ареал свидетельствует о том, что перед нами азиатско-восточно-североамериканский арктотретичный вид, имеющий изолированный участок ареала на Аляске и в Северо-Западной Канаде.
Evernia perfragilis Llano - криоксерофитный, эпигейный, кочующий лишайник (рис. 7). Одна часть ареала этого вида связана с горными районами Северной Монголии, где он обитает преимущественно в высокогорном поясе, на типчаково-осоково-кобрезиевых пустошах, в горных степях, изредка в лиственничных лесах в Хангае, в Хобдоском горно-лесостепном районе, в Монгольском Алтае, а также в низовьях р. Хунгуй, в Котловине Больших Озер (Голубкова, 1981; Цогт, 1981). В соседних с Монголией районах был найден в Тувинской АССР, в нагорье Сангилен, где был собран Н. В. Седельниковой в истоках р. Каргы, на высоте 2000 м над ур. м., в щебнистых тундрах (Гербарий БИН АН СССР). Вторая часть ареала этого вида связана главным образом с азиатским и американским секторами Арктики. В азиатской Арктике встречается на Новосибирских островах - о-в Котельный (Городков, 1956) и на Чукотском полуострове, где обитает на почве в горных каменистых лишайниково-моховых тундрах в Анюйском нагорье, на побережье залива Лаврентия и в хр. Искатень (Локинская, 1970; Макарова, 1979а; Андреев, 1980). В Северной Америке приводится для Аляски, Северо-Западных Территорий и Баффиновой Земли (Llano, 1951; Krog, 1968; Thomson, 1970, 1979; Bird, 1974; Bird e. a., 1980). Evernia perfragilis Llano также проникает на о-ва Новая Земля (Еленкин, Савич, 1912; Lynge, 1928). В условиях Монголии он, помимо высокогорного пояса, изредка обитает и в лесных ценозах. Видимо, исходный ареал этого вида находится в пределах Хангае-Алтайской горной страны. Арктическая часть ареала, по-видимому, вторичная. В арктические районы Азии Е. perfragilis Llano мог проникнуть во время фазы максимального, самаровского, оледенения верхнего плейстоцена, когда установились флористические связи между горными системами Сибири и Арктикой в связи с широким распространением перигляциальных пространств, на которых происходило формирование ареалов арктовысокогорных видов.
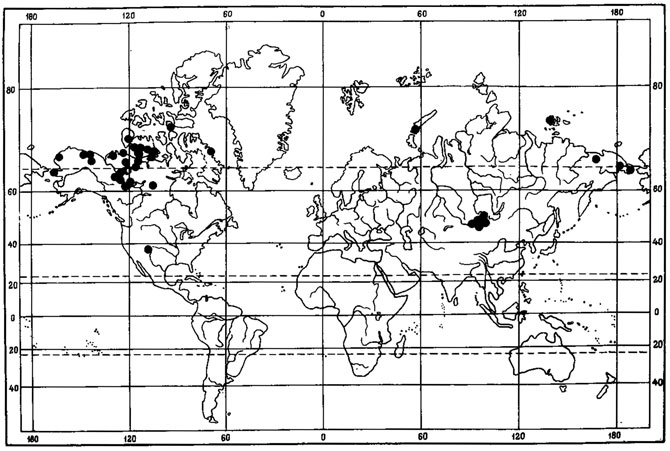
Рис. 7. Распространение на земном шаре Evernia perfragilis Liano (Ориг.)
4. Евразиатский тип ареала характерен для 13 видов, произрастающих в арктических и высокогорных районах Европы и Азии. В их числе Placidiopsis cervinula (Nyl.) Zahlbr., Peltigera lygnei Gyeln., Catillaria subnitida Hellb., Rhizocarpon phalerosporum (Vain.) H. Magn., Aspicilia supertegens Arnold, A. lapponica (Zahlbr.) Oxn., A. verruculosa Krempelh., A. aliena (Zahlbr.) Oxn., A. aquatica Koerb., A. cingulata (Zahlbr.) Oxn., A. chidinensis (H. Magn.) Oxn., A. perradiata (Nyl.) Hue, A. polychroma Anzi.
Как можно видеть, среди арктовысокогорных лишайников с евразиатским распространением выделяется большая группа аспицилий. Представители рода Aspicilia преимущественно ксерофитные лишайники, обитающие главным образом на каменистых субстратах, реже на почве и растительных остатках в арктических, горных и пустынно-степных районах Голарктики. Род насчитывает 116 видов, из них 40% является арктическими, 20% - арктовысокогорными, 20% - монтанными, 36% видов связано в своем распространении с пустынно-степными и горноаридными районами. К сожалению, эта систематическая группа лишайников изучена далеко не достаточно, так как аспицилии являются чрезвычайно трудными для идентификации лишайниками. Поэтому данные о распространении целого ряда видов этого рода отрывочны. Например, Aspicilia cingulata (Zahlbr.) Oxn. до настоящего времени считалась эндемиком о-вов Новая Земля, теперь же найдена в Монголии, в горных степях Хангая (Бязров и др., 1983). В этом же районе Монголии была собрана A. aliena (Zahlbr.) Охп., до сих пор известная только в Швеции и на о-вах Новая Земля, а также A. polychroma Anzi, приводимая ранее также для о-вов Новая Земля и горных районов Северной Италии- Ломбардии (Окснер, 1971б; Бязров и др., 1983). Из аркто-высокогорных аспицилий евразиатского распространения наибольший интерес представляет A. aquatica Koerb., которая относится к интересной экологической группе лишайников, так называемых подводных лишайников, обитающих на камнях, валунах и скалах в горных речках, озерах, ручьях. Основной ареал A. aquatica Koerb. охватывает арктическую и Северную Европу, а также горы Средней Европы. Распространение этого вида в Азии, видимо, недостаточно изучено. До его находок в Монголии, в Хангае, в верховьях р. Хух-Сум (Бязров и др. 1983), он был известен в арктических Приенисейских районах, а также на п-ове Камчатка (Окснер, 1971б).
Несомненно представляют интерес также находки в Монголии редкого пиренокарпового лишайника - Placidiopsis cervinula (Nyl.) Zahlbr. Род Placidiopsis насчитывает 17 видов (Бредкина, 1972), распространение которых связано главным образом со средиземноморской областью и районами Средней Европы. P. cervinula (Nyl.) Zahlbr., вероятно, единственный представитель рода арктовысокогорного распространения, хотя ареал этого вида изучен недостаточно. Долгое время он был известен лишь из классического местонахождения в Понойской Лапландии на Кольском полуострове (Nylander, 1864; Vainio, 1921; Домбровская, 1970а). Недавно этот лишайник был указан для флоры п-ова Таймыр (Пийн, Мартин, 1978), где был собран на почве на границе лиственничной редины и пятнистой тундры в бассейне р. Хатанги (р. Новая). В Монголии P. cervinula (Nyl.) Zahlbr. обитает в высокогорных степях в Хангае и Монгольском Алтае. В Хангае он был встречен в районе Главного Хангайского хребта, на высоте 2700 м над ур. м., в окрестностях оз. Хух-Нур, где обитал на замшелой почве у основания скал, в Монгольском Алтае - на перевале Улакчин-Даба, на высоте 3050 м над ур. м., в полынно-типчаково-подушечной степи (Голубкова, 1981; Цогт, 1981).
Арктическая часть ареала Rhizocarpon phalerosporum (Vain.) Н. Magn. охватывает о-ва Новая Земля (Магнуссон, 1927; Lynge, 1928, 1934b, 1936), Фенноскандию (Vainio, 1883, 1922; Магнуссон, 1927; Rasanen, 1943; Koskinen, 1961; Домбровская, 1970а, 1970б; Ulvinen е. а., 1978, и др.), Полярный Урал (Магнуссон, 1927), Чукотский полуостров, побережье Берингова пролива (Lynge, 1934b, 1936). В горных районах известен в Украинских Карпатах (Макаревич, 1963), в Кузнецком нагорье (Седельникова, 1978) и в Монголии, где был собран А. А. Еленкиным в окрестностях оз. Хубсугул (Голубкова, 1981).
5. Голарктический тип ареала свойствен большой группе лишайников - 55 видам, обитающим в арктических районах и высокогорьях Голарктики. К этой группе лишайников относятся виды родов Solorina - S. saccata (L.) Ach. (рис. 8, 1), S. crocea (L.) Ach., S. octospora Arnold; Caloplaca - C. cinnamomea (Th. Fr.) Oliv., C. epithallina Lynge, C. jungermanniae (Vahl) Th. Fr.; Lecidea - L. garovaglii Schaer., L. assimilata Nyl., L. limosa Ach.; Cetraria - C. tilesii Ach., C. nivalis (L.) Ach., C. cucullata (Bellardi) Ach., C. hepatizon (Ach.) Vain.; Parmelia stygia (L.) Ach., Peltigera scabrosa Th. Fr., Nephroma arcticum (L.) Torss., Dactylina madreporiformis (Wulf.) Tuck., Cornicularia divergens Ach., Ochrolechia inaequatula (Nyl.) Zahlbr., Pachyospora verrucosa (Ach.) Massal., Pertusaria panyrga (Ach.) Massal., Pannaria praetermissa Nyl., Umbilicaria torrefacta (Lightf.) Schrad., U. virginis Schaer. и др.
Одни виды этой группы имеют циркумполярное распространение в Арктике и в высокогорьях умеренной Голарктики. Другие часто встречаются в арктических районах и высокогорьях Европы, в то время как в Азии отмечаются отдельные изолированные местообитания их в Сибирской Арктике и ее горных районах. Некоторые же виды этой группы, наоборот, имеют центры массовости своего распространения на территории Азии и являются редкими в Европейской Арктике и в высокогорьях Европы и Северной Америки. Попытаемся проследить особенности распространения лишайников этой географической группы на некоторых примерах.
К числу видов, имеющих центры массовости своего развития на территории Азии, принадлежат арктовысокогорные виды рода Cetraria - С. tilesii Ach., С. cucullata (Bellardi) Ach., С. nivalis (L.) Ach. Так, например, К. А. Рассадина (1950), характеризуя ареал С. tilesii Ach., отмечала, что этот лишайник - представитель тундр, почти исключительно встречается в Сибири. Ареал С. tilesii Ach. (Рассадина, 1950, 1971) на территории Азии охватывает острова Северного Ледовитого океана (острова Вайгач, Северная Земля) и арктическое побережье Сибири, от Полярного Урала, п-ова Таймыра до низовьев Лены, Чукотского полуострова, а также Якутию, Магаданскую область, долину р. Анадыря, Северную Корякию, Камчатку, и далее, южнее - Алтай, Саяны, побережье оз. Байкал, хр. Сайлюгем, горы Северо-Западной Монголии. На территории Монголии по хребту Цаган-Шибэту и Главному Хангайскому хребту, вероятно, проходит южная граница ареала этого вида в пределах Азии. В Монголии этот лишайник встречается в Прихубсугулье, в хр. Цаган-Шибэту и в Центральном Хангае (окрестности оз. Цаган-Нур, Хух-Нур, перевал Эгин-Даба), где обитает в высокогорном поясе, на вершине гольцов, в кобрезиевых пустошах, высокогорных степях, изредка в горно-таежном поясе, в пределах высот 2100 - 3100 м над ур. м. (Голубкова, 1981; Цогт, 1981). В Северной Америке распространение этого лишайника связано с Аляской и северными районами Канады - бассейнами рек Юкон, Пил, Маккензи, с Северо-Западными Территориями и др. (Lynge, 1937, 1939; Krog, 1968; Thomson, 1979; Bird е. a., 1980, и др.). В Европе С. tilesii Ach. довольно широко распространен на Полярном Урале и Среднем Урале; на о-вах Новая Земля встречается только на севере (Lynge, 1928; Рассадина, 1950); спорадически - высоко в горах Скандинавии и Средней Европы (Альпы, Татры). В равнинных условиях этот лишайник найден на островах Готланд и Эланд (Швеция) и в Карелии (Vainio, 1940; Motyka, 1960; Домбровская, 1970а, Dahl, Krog, 1973). Этот вид известен также на Кавказе (Рассадина, 1950).
Характеризуя распространение С. cucullata (Bellardi) Ach., К. А. Рассадина (1971) отмечала также, что этот вид достигает более пышного развития в азиатской части СССР. Так, этот лишайник встречается в разнообразных типах тундр, как равнинных, так и горных, Азии, от Полярного Урала на западе до Чукотского полуострова, Анадыря, Камчатки, Сахалина, Приморского края на востоке и от Арктической Сибири на севере до Алтая, Саян, Хакассии, Северо-Западной Монголии на юге (Рассадина, 1950). В пределах Монголии С. cucullata (Bellardi) Ach. довольно широко распространен на кобрезиевых пустошах, в горных тундрах высокогорного пояса, изредка в лиственничных лесах в Прихубсугулье, Хэнтэе, Хангае, в Монгольском Алтае, в хребтах Цаган-Шибэту и Тургэн-Ула (Голубкова, 1981; Цогт, 1981). На территории Азии этот лишайник достигает южной части Тибета (Wei, Chen, 1974) и Гималаев (Wei, Jiang, 1981).
Основное распространение других видов этой группы арктовысокогорных лишайников связано с арктическими и горными районами Европы, в то время как на территории Азии они имеют изолированные местообитания. К числу таких лишайников относятся виды рода Lecidea - L. limosa Ach., L. assimilata Nyl., L. garovaglii Schaer.
Например, основной ареал L. limosa Ach. (Окснер, 1968) охватывает северные и горные районы Европы, где этот вид встречается в Фенноскандии, Карельской АССР, Великобритании, Альпах, Карпатах, Тироле, в горах ФРГ, Польши, Чехословакии. В Азии L. limosa Ach. является редким видом, найден лишь в восточной части Чукотского полуострова и в Анюйском нагорье (Vainio, 1909; Макарова, 1979а), южнее же - лишь в Монголии (Голубкова, 1981; Цогт, 1981) и в Кузнецком нагорье (Седельникова, 1978). Этот лишайник является редким также и в Северной Америке; он известен на Аляске на п-ове Сьюард и на о-ве Элсмир, приводится также для Гренландии (Thomson, 1979). В Монголии собран лишь однажды в долине р. Тургэн-Гол, на каменистом берегу в лиственничном редколесье, в интересном эпигейном сообществе лишайников, сочетающем арктовысокогорные и пустынно-степные виды, о чем будет сказано ниже.
Ареал другого эпибриофитного лишайника - L. assimilata Nyl. охватывает арктические и горные районы Европы, где он встречается на о-ве Медвежий, в Северной Фенноскандии, на Кольском полуострове, в горах Великобритании и в других западных районах Европы, в Тироле, в горных районах южной части ФРГ, в ГДР - в Саксонии, в горах Чехословакии, Польши, Венгрии, в СССР - в Карпатах (Окснер, 1968). В Северной Америке этот арктовысокогорный лишайник растет в арктических районах, от о-ва Элсмир до Аляски, южнее встречается в Канаде - в Квебеке и провинции Саскачеван, в США - в штатах Колорадо и Вашингтон (Thomson, 1979), известен также в Гренландии. На территории Азии отмечен на Таймыре (Пийн, 1979б), на Чукотском полуострове (Nylander, 1888; Malrne, 1932; Макарова, 1979а; Андреев, 1980), в восточной Якутии (Городков, 1956), в бассейне р. Индигирки (Афонина и др., 1979. 1980); южнее - в Монголии и Гималаях.
На территории МНР L. assimilata Nyl. - редкий вид, был собран лишь в Хангае, в окрестностях оз. Хух-Нур, на высоте 2700 м над ур. м., в змеевково-житняковых горных степях, и в верховьях р. Чулутын-Гол, на высоте 2200 м над ур. м., по склонам гор (Голубкова, 1981; Цогт, 1981).
Ареал эпилитного лишайника L. garovaglii Schaer. (Hertel, 1977) охватывает Арктику, горы умеренной Европы, где он встречается от Пиренеев до Альп и Карпат, а также горные районы Северной Америки. В Азии этот вид известен лишь в Красноярском крае, в окрестностях г. Минусинска (Vainio, 1896), и в Монголии, где был собран только в районе Прихубсугулья (Голубкова, 1981).
Изолированные местообитания на территории Азии имеет также эпигейный лишайник Solorina octospora Arnold. Он известен только на п-ове Таймыр (Пийн, 1979а, 19796), на Чукотском полуострове (Макарова, 1979а), в Якутии, на Алтае (Савич, 1975), а также в Монголии (Хангай, долина р. Чулутын-Гол) (Голубкова, 1981; Цогт, 1981). В то же время этот вид широко распространен в Европе, в Скандинавии, на Кольском полуострове, в Карельской АССР, в Прибалтике, в горах Средней Европы - Пиренеи, французское центральное плато, Альпы, Судеты, Карпаты, Балканы (Савич, 1975), известен также на Кавказе. В Северной Америке S. octospora Arnold широко распространен в арктических районах от о-ва Элсмир до Аляски, южнее - в горах штатов Колорадо и Нью-Мексико, а также в Скалистых горах (Thomson, 1979). Этот лишайник встречается также в Гренландии и на о-вах Шпицберген.
Другие виды группы арктовысокогорных солорин циркумполярно распространены как в арктических районах, так и в высокогорьях Голарктики, в том числе часто встречаются и на территории Азии. Так, ареал S. saccata (L.) Ach. охватывает арктические районы и высокогорья Европы, от о-вов Шпицберген и Британских островов до Апеннинского, Балканского полуостровов и от Пиренейского полуострова до Чехословакии, Венгрии, СССР - Карпаты. Он обитает также в высокогорьях Северной Африки и Кавказа. Этот вид широко распространен в арктических районах Северной Америки, на юге достигает северных штатов США - Вермонт, Висконсин, Южная Дакота, и Канады - провинций Альберта и Британская Колумбия (Thomson, 1979), известен также в Гренландии и на Фарерских островах. На территории Азии встречается на Таймыре (Пийн, 1979б), на Чукотском полуострове (Nylander, 1888; Макарова, 1973, 1979а), в Якутии (Окснер, 1939а; Городков, 1956), южнее - на Алтае, в Саянах, Прибайкалье, на Тянь-Шане, в Хабаровском крае и на Камчатке, а также в Монголии. Этот лишайник довольно широко распространен в северо-западных районах Монголии, в Прихубсугулье, Хэнтэе, Хангае, Монгольском Алтае, где обитает на карбонатной почве и мхах, у основания скал во влажных и тенистых местах по склонам гор, в лиственничных лесах, в высокогорном поясе (Голубкова, 1981; Цогт, 1981) (рис. 8, 1). Как и S. saccata (L.) Ach., S. сrосеа (L.) Ach. часто встречается в Европе, от арктических районов (острова Шпицберген, Исландия, Медвежий, Фенноскандия) до горных районов Великобритании, Франции, Пиренеев, Альп, Карпат, гор Италии, Австрии, ГДР, ФРГ, Польши, Чехословакии, Венгрии, а также Урала, известен на Кавказе. В Северной Америке этот лишайник широко распространен в арктических районах, от о-ва Элсмир до Аляски, проникает в более южные районы до п-ова Лабрадор, Квебека, Нью-Мексико и Вашингтона; он известен также в Гренландии. На территории Азии часто встречается в арктических тундрах от п-ова Таймыр до Чукотского полуострова, в Магаданской области, в восточной Якутии, южнее - на Алтае, в Прибайкалье, Саянах, в Казахстане (Заилийский Алатау), в Монголии, на Камчатке. В Монголии S. сrосеа (L.) Ach. - редкий лишайник, собранный лишь в западном Прихубсугулье, в долине р. Хотон-Гол, на высоте 2800 м над ур. м.
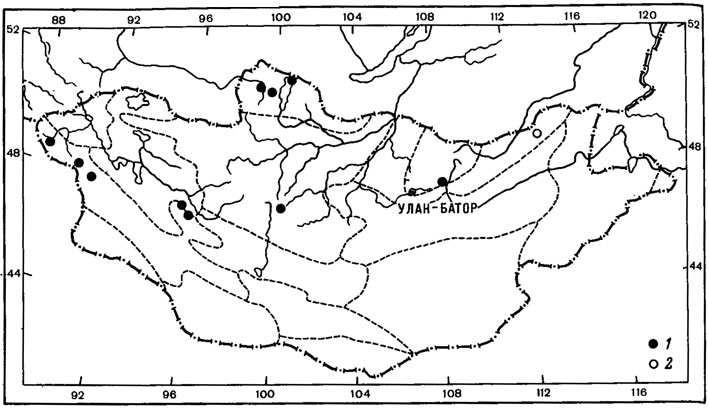
Рис. 8. Распространение на территории Монголии Solorina saccata (L.) Ach. (1) и Leptogium hildenbrandii (Garov.) Nyl. (2). (Ориг.)
Б. Омниарктовысокогорный субэлемент. Объединяет 33 вида лишайников, основное распространение которых связано с приполярными и высокогорными районами Голарктики и другими флористическими царствами. Ареалы этих видов характеризуются следующими типами.
1. Мультирегиональный тип ареала свойствен 14 видам. Среди них виды рода Umbilicaria - U. proboscidea (L.) Schard., U. cylindrica (L.) Del., U. vellea (L.) Ach., U. hyperborea (Ach.) Hoffm., Biatora leucophaea (Vahl) Flk., Lecanora pulicaris (Pers.) Ach., Parmelia omphalodes (L.) Ach., Alectoria ochroleuca (Hoffm.) Massal., Cladonia lepidota Nyl., Ochrolechia upsaliensis (L.) Massal., Acarospora badiofusca (Nyl.) Th. Fr. (рис. 9) и некоторые другие. Это виды с обширными дизъюнктивными ареалами, охватывающими арктические и высокогорные районы Голарктики и высокогорья других флористических царств.
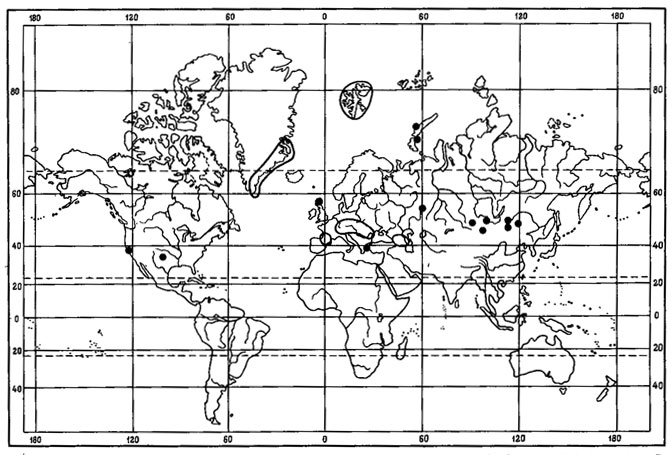
Рис. 9. Распространение на земном шаре Acarospora badiofusca (Nyl.) Th. Fr. (Ориг.)
В качестве примера может служить ареал Umbilicaria proboscidea (L.) Schrad. (Llano, 1950; Голубкова, Савич, 1978). Этот лишайник относится к числу криоксерофитных, эпилитных видов, обитающих на скалах, каменных глыбах, валунах и камнях по каменистым россыпям в арктических районах, в горах в альпийском и субальпийском поясах, иногда на открытых местах верхней границы лесного пояса. U. proboscidea (L.) Schrad. циркумполярно распространен в Арктике, на о-вах Шпицберген, в северной Скандинавии, на Кольском полуострове, Земле Франца-Иосифа, на островах Новая Земля, Вайгач, на Новосибирских островах, о-ве Врангель, побережье Сибири, Чукотском полуострове, островах Прибылова, Алеутских островах, на Аляске, в долине р. Юкон, в Северо-Западных Территориях, на островах Элсмир, Баффинова Земля, полуострове Лабрадор, в Ньюфаундленде, в западной Гренландии, на островах Исландия, Ян-Майен. В Европе, помимо арктических районов, встречается главным образом в горах Средней Европы, в Альпах, достигая высоты 3400 м над ур. м., в Судетах, Татрах, Карпатах, Вогезах и в других районах. Но имеются изолированные местообитания этого вида и в равнинных условиях - в Карелии, Архангельской, Ленинградской областях, в Эстонии и некоторых других. Этот лишайник встречается на Кавказе, Урале, в горах Южной Сибири, в Западной и Восточной Сибири, на Дальнем Востоке, на Камчатке, в горах Японии, в Монголии. На территории Северной Америки, помимо арктических районов, растет в высокогорьях Скалистых гор и Аппалачей, в штатах Вашингтон, Орегон, Колорадо, Нью-Йорк, Нью-Гэмпшир, Вермонт, Мэн (Llano, 1950). Кроме того, U. proboscidea (L.) Schrad. обитает в горах Центральной Америки и в Австралии. На территории Монголии этот вид был собран только в Прихубсугулье, в окрестностях оз. Хубсугул (Голубкова, 1981).
Большинство видов этой группы лишайников на территории Монголии является редкими, например Biatora leucophaea (Vahl) Flk., Lecanora pulicaris (Pers.) Ach., Parmetia omphalodes (L.) Ach. и др. К наиболее широко распространенным видам принадлежат Alectoria ochroleuca (Hoffm.) Massal., Ochrolechia upsaliensis (L.) Massal. В районе Прихубсугулья, а также в Хэнтэе, Хангае Alectoria ochroleuca (Hoffm.) Massal. играет определенную фитоценотическую роль в растительных ценозах высокогорного пояса. Этот кустистый лишайник вместе с видами родов Cetraria, Cladonia, Cladina принимает заметное участие в сложении эпигейных лишайниковых группировок в составе кобрезиевников, высокогорных тундр и других растительных сообществ высокогорного пояса. Ареал этого лишайника охватывает арктические районы и высокогорья Европы, Азии (Сибирь, МНР, КНР, Япония, Индия, Непал), Северной Америки. За пределами северного полушария Alectoria ochroleuca (Hoffm.) Massal. встречается в горах Центральной и Южной Америки (Анды, о-в Огненная Земля), Новой Зеландии.
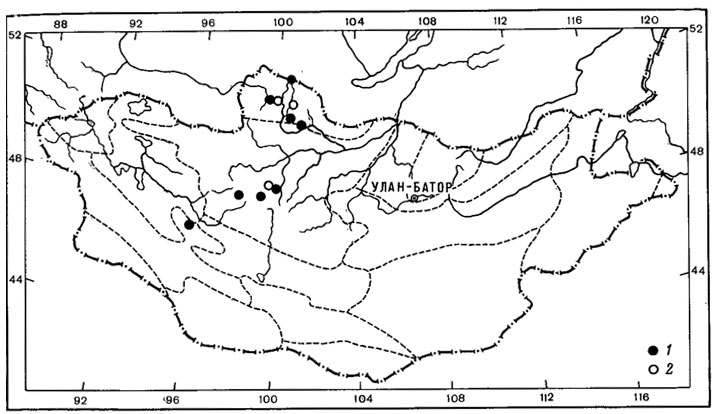
Рис. 10. Распространение на территории Монголии Ochrolechia upsaliensis (L.) Massal. (1) и O. inaequatula (Nyl.) Zahlbr. (2) (Ориг.)
Широко распространен в северо-западных районах Монголии (Прихубсугулье, Хангай, Монгольский Алтай) эпибриофитный лишайник Ochrolechia upsaliensis (L.) Massal. (рис. 10, 1). Этот лишайник, поселяющийся на мхах и растительных остатках встречается не только в высокогорном поясе, в составе высокогорных степей, кобрезиевых пустошей, но и в лесах, а также в горных степях среднегорного пояса. Некоторые виды из этой группы арктовысокогорных лишайников в условиях Монголии ведут себя как типичные "сниженные альпийцы", обитая в нижнегорных поясах горных районов. К их числу, например, относится Cladoniia lepidota Nyl. Обширный ареал этого вида охватывает северное и южное полушария (Окснер, 1968). Он циркумполярно распространен в Арктике, местами проникает в таежную зону и растет высоко в горах Европы, Азии и Северной Америки. На территории Азии, помимо арктических районов, этот лишайник встречается в Омской и Томской областях, Красноярском крае, в Якутии, на Алтае, в Саянах, в Читинской области, в Монголии, Японии, а также на о-ве Ява. За пределами Голарктики произрастает в горах Африки (острова Маскаренские, Реюньон, Мадагаскар), в Южной Америке (Перу) и Австралии. На территории; Монголии этот арктовысокогорный лишайник был встречен в пределах нижнегорного пояса в хр. Эрэн-Даба-Нуру, на высоте 1200 - 1300 м над ур. м., в сосновых лесах, а также в Хэнтэе, в верховьях р. Тола, где он обитал в пойме реки на почве среди мхов.
2. Биполярный тип ареала свойствен 19 видам - Sphaerophorus globosus (Huds.) Vain., Lecidea atrobrunnea (Ram.) Schaer., Haematomma ventosum (L.) Massal., Pseudephebe minuscula (Nyl.) Brodo et D. Hawksw., P.. pubescens (L.) Choisy, Bryoria chalybeiformis (L.) Brodo et D. Hawksw., Neuropogon sulphureus (Koenig) Hellb., Umbilicaria decussata (Vill.) Frey, Blastenia leucoraea (Ach.) Th. Fr., Lecanara badSa (HoSfm.) Achi, L. epibryon (Ach.) Ach., Ochrolechia frigida (Sw.) Lynge, Tremolecia atrata (Ach.) Hertel, Psoroma hypmorum (Vahl) S. Gray, Rhizoplaca melanophthalma (Ram.) Poelt et Leuck., Sporastatia testudinea (Ach.) Massal, Thamnolia vermicullaris (Sw.) Ach, Phaeorrhiza nimbosa (Fr.) Mayrh. et Poelt (рис. 11), St'ereocaulon alpinum Laur., Rhizocarpon superficiale (Schaer.) Vain., Buellia papillata (Sommerf.) Tuck. Они характеризуются обширным дизъюнктивным ареалом, одна часть которого связана с Арктикой, другая - с высокогорьями земного шара, третья - с Антарктикой.
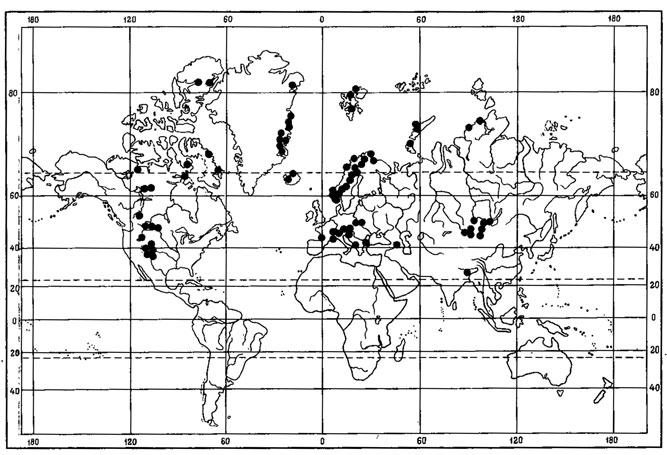
Рис. 11. Распространение на земном шаре Phaeorrhiza nimboza (Fr.) Mayrh. et Poelt. (По: Mayrhofer, Poelt, 1978, с дополнениями автора)
Типичным примером биполярного ареала может служить ареал Neuropogon sulphureus (Koenig) Hellb. (рис. 12, 1). Этот кустистый эпилитный лишайник был найден в, Монголии лишь однажды в Хангае, на горе Отгон-Тэнгэр (или Энх-Тайван) в нивальном поясе, на высоте 3550 м над ур. м., среди вечного снега на камнях андезитово-базальтовых пород.
Находка представителя рода Neuropogon, в Монголии, на склоне горы Отон-Тэнгэр Хангайского хребта, - первое местонахождение этого рода в высокогорьях северного полушария (Бязров, 1979). Ареял этого биполярного лишайника охватывает острова Вайгач, Земля Франца-Иосифа, Шпицберген, Исландия, Ян-Майен, Гренландия, Мелвилл, а также побережье Северного Ледовитого океана; после огромной дизъюнкции этот вид встречается в высокогорьях Эквадора, Чили, Перу, Аргентины и на побережьях Антарктики (Motyka, 1936; Lamb, 1964; Голубкова, Савич, 1969). Как предполагает А. Н. Окснер (1944), продвижение антарктических лишайников, в том числе и Neuropogon sulphureus (Koenig) Hellb., в северное полушарие, как и вообще миграции биполярных лишайников, могло происходить только через венесуэльско-колумбийские цепи и Антильский (ныне погруженный) складчатый район в основном в конце третичной эпохи и в квартере. Проникнув в горы Северной Америки, вплоть до районов, примыкающих к арктической области, N. sulphureus был снижен наступившим горным оледенением в неоледеневшие равнины американской Арктики; оттуда по неоледеневшим пространствам полярной области проник на запад до о-ва Беннетта, а на восток - к о-ву Мелвилл и далее в Гренландию и Атлантический сектор Арктики вплоть до Новой Земли (Окснер, 1944). Видимо, не исключена возможность миграции этого вида вместе с волнами отступавших с севера арктийцев во времена плейстоцена далеко к югу через пространства Сибири вплоть до горных районов Южной Сибири, откуда он мог проникнуть и в высокогорья Монголии. Как указывал А. А. Юнатов (1950), отдельные высокогорья Монголии, в настоящее время совершенно изолированные, ранее, в эпоху оледенения, без сомнения, имели связь с горными цепями северо-востока Сибири; горно-тундровая растительность последних далеко проникала к югу. Можно предположить, что изолированный ареал этого вида в горах Хангая является реликтовым, свидетельствующим о более обширной голарктической части ареала этого лишайника в плейстоцене. К сожалению, высокогорья Восточной Сибири в лихенологическом отношении изучены очень слабо, но в дальнейшем не исключена находка этого лишайника и в других горных районах региона.
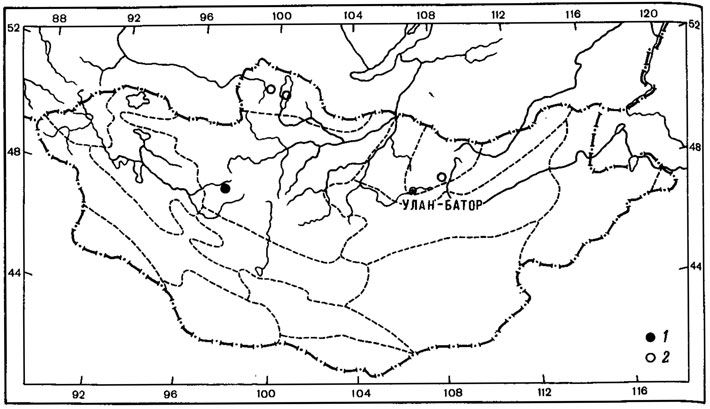
Рис. 12. Распространение на территории Монголии Neuropogon sulphureus (Koenig) Hellb. (1) и Bryoria lanestris (Ach.) Brodo et D. Hawksw. (2) (Ориг.)
В заключение анализа арктовысокогорного элемента следует отметить следующее.
Как можно видеть, арктовысокогорный элемент играет заметную роль в составе лихенофлоры Монголии, охватывая 1/7 часть общего числа видов. Это несомненно представляет интерес, если учесть, что высокогорные пустоши и горные тундры представлены на территории МНР весьма скудно, занимая всего 3% от общей площади Монголии (Юнатов, 1954). Однако при преобладающем развитии на территории Монголии степного (65% всей площади страны) и пустынного (15%) типов растительности арктовысокогорный элемент как бы выпадает из общей специфики лихенофлоры этой страны, которой в целом свойственны пустынно-степные черты. Большинство арктовысокогорных лишайников является редкими на территории Монголии, и местообитания их ограничены лишь высокогорным поясом горных районов северозападной части страны, где, однако, некоторые из них играют заметную роль в растительном покрове. Характерным для растительного покрова Монголии является широкое участие арктовысокогорных видов лишайников в составе лесных, горно-степных, а также равнинно-степных растительных сообществ Монголии. Широкое участие арктовысокогорных видов в составе лесных и степных ценозов является еще одним свидетельством того, что как лесная, так и степная растительность Монголии в наибольшей степени, чем где-либо в Северной Евразии, сохранила черты комплекса холодной плейстоценовой лесостепи (Крашенинников, 1954а).
Ареалогический анализ видов арктовысокогорного элемента показывает, что 51.4% видов характеризуется широкоголарктическими ареалами, 12.1 - евразиатскими, 2.8% - азиатско-американскими; они отражают широкие флористические связи между горными системами Сибири и Северной Монголией, Арктикой, а также между другими горными системами Голарктики, существовавшие, видимо, в плейстоцене. Время вхождения арктовысокогорного элемента в лихенофлору Монголии, по-видимому, относится к плейстоцену.
В генетическом отношении арктовысокогорный элемент лихенофлоры Монголии представляет собой гетерогенную группу видов. Одни из них горного голарктического генезиса, другие арктического происхождения, генезис третьих связан с высокогорными и полярными районами южного полушария. К числу систематических групп, имеющих горное происхождение, по мнению А. Н. Окснера (1956), относится, например, род Solorina. Из числа других лишайников он указывает на горный голарктический генезис Lecanora epibryon Ach. (Окснер, 1944). Генезис рода Neuropogon, по мнению А. Н. Окснера (1944), связан с высокогорными и полярными районами южного полушария.
Видимо, многие представители арктовысокогорных видов лишайников, найденные в составе флоры Монголии, азиатского горного генезиса. К числу таких лишайников, например, относятся арктовысокогорные виды родов Asahinea, Cetraria, Evernia и другие. Например, можно предполагать, что областью первоначального развития представителей рода Asahinea были, по-видимому, горные районы Ангариды. Олиготипный род Asahinea насчитывает 3 вида - Asahinea chrysantha (Tuck.) Culb. et Culb., A. scholanderi (Llano) Culb. et Culb., A. kurodakensis (Asah.) Culb. et Culb. (Culberson, Culberson, 1965). Этот олиготипный род имеет берингийское распространение. Ареал Asahinea chrysantha (Tuck.) Culb. et Culb. (рис. 6) подробно обсуждался выше. Второй вид этого рода - A. scholanderi (Llano) Culb. et Culb. - характеризуется ярко выраженным берингийским ареалом, будучи распространенным в Азии только в Восточной Сибири, к востоку от р. Лены и до побережья Тихого океана. На территории МНР этот лишайник не встречается, южная граница его ареала в Азии проходит в районе оз. Байкал (Окснер, Рассадина, 1960; Будаева, 1976), однако не исключены его находки в горных районах Северной Монголии. В Северной Америке распространение A. scholanderi (Llano) Culb. et Culb. ограничено Северной Аляской и северозападными районами Канады, где восточная граница его ареала проходит в районе бассейна р. Бак и оз. Бейкер-Лейк (Culberson, Culberson, 1965; Thomson, 1979). Вид этого же рода - A. kurodakensis (Asah.) Culb. et Culb. - является эндемиком островов Японии - Хонсю и Хоккайдо (Culberson, Culberson, 1965).
Как полагает А. Н. Окснер и К. А. Рассадина (1960), Asahinea chrysantha (Tuck.) Culb. et Culb. - вид горного генезиса, существовавший как высокогорный, вероятно, уже в условиях горных сооружений Ангариды и сохранивший до настоящего времени такие же позиции в горных хребтах южной части Восточной Сибири, которые хотя и испытывали частичное оледенение, но, несомненно, являлись одним из районов консервирования третичных форм. По протянувшимся в долготном направлении хребтам Восточной Сибири A. chrysantha (Tuck.) Culb. et Culb. продвинулся далеко к северу и, оказавшись в пределах Якутского сектора Арктики, начал распространяться циркумполярно, но встреченные им, находившиеся под ледяным покровом территории полярной области не позволили его ареалу замкнуть арктическое кольцо. Дойдя в своем расселении на запад до Полярного Урала, он спускается по горным хребтам к югу и проникает в эпоху последнего оледенения в пределы Южного Урала. Как отмечают А. Н. Окснер и К. А. Рассадина (1960), обращает на себя внимание, что распространение A. chrysantha (Tuck.) Culb. et Culb. в Арктике незначительно выходит за пределы неоледеневших областей и повторяет в общем тип ареала эоарктических видов (Толмачев, 1932). Но, разделяя с эоарктийцами историю их расселения в Арктике, A. chrysantha (Tuck.) Culb. et Culb. резко отличается от них историей формирования своего ареала, своим горным генезисом. Горное, а не арктическое происхождение этого вида подтверждается не только особенностями южной части его ареала, но и его морфологическим обликом. Среди арктических по происхождению лишайников нет относящихся к широколистоватому типу, ярчайшим представителем которого является не только A. chrysantha (Tuck.) Culb. et Culb., но и другие виды этого рода. Морфологическое строение представителей рода Asahinea, характеризующихся крупнолистоватыми дорзивентральными слоевищами, дает основание высказать предположение, что исходные формы этой систематической группы лишайников, видимо, были связаны с горно-лесным поясом. A. scholanderi (Llano) Culb. et Culb. и A. kurodakensis (Asah.) Culb. et Culb. по морфологическим признакам, наличию изидиев и содержанию лишайниковых веществ очень близки между собой (Culberson, Culberson, 1965). Возможно, что они произошли от одной исходной формы в результате распада ее обширного ареала и развития A. kurodakensis (Asah.) Culb. et Culb. в условиях географической изоляции на островах Японии. A. scholanderi (Llano) Culb. et Culb., по-видимому, довольно молодой вид, возникший во второй половине четвертичного периода (Окснер, Рассадина, 1960). Но этот вид, вероятно, должен был пережить эпоху последнего оледенения, иначе трудно представить и понять его северо-восточный ареал, ограниченный на западе р. Леной, хорошо очерченный и захватывающий как горную, так и арктическую часть Дальнего Востока.
Таким образом, современное географическое распространение, морфологические особенности представителей рода Asahinea, их обитание на замшелых скалах и на почве среди других видов лишайников и мхов в горных и равнинных тундрах - все это позволяет высказать предположение, что областью первоначального развития этой группы лишайников были горные районы Азии. Это согласуется с мнением А. И. Толмачева (1958), который полагал, что исходными для формирования элементов высокогорных флор являются преимущественно флоры умеренных, континентальных областей, в частности умеренной Азии. Он отмечал, что на протяжении длительного времени в умеренных, широтах, в том числе в Азии, имелись предпосылки для формирования высокогорных растений. В умеренной Азии этому способствовал как муссонный климат этих районов, так и наличие с доисторических, времен более или менее выраженного рельефа и связанное с этим разнообразие типов местообитания, т. е. те биоценотические условия, которые существовали здесь с конца мелового периода и по меньшей мере по миоцен. Как было показано выше, на примере рода Asahinea, в горах, уже существовавших в третичном периоде, в районе умеренной Азии могли сформироваться более специализированные горные представители азиатских по своему происхождению лишайников.
Помимо этого рода, другим примером арктовысокогорных лишайников, генезис которых связан с горами умеренной Азии, могут служить кустистые виды рода Cetraria - С. tilesii Ach., С. cucullata (Ballardi) Ach., С. nivalis Ach.
При характеристике ареалов этих арктовысокогорных видов подчеркивалось, что центр массовости развития этих видов находится в Азии. На горный генезис Cetraria nivalis Ach. и С. cucullata (Bellardi) Ach. указывала К. А. Рассадина (1948, 1950), монограф этой систематической группы лишайников. Она отмечала, что распространение видов рода Cetraria на земном шаре показывает, что исходный центр происхождения Cetraria надо искать в Азии, в горах Южной Сибири, в Гималаях, Китае. При этом следует подчеркнуть, что исходные формы этой группы лишайников, как и в случае рода Asahinea, видимо, были листоватыми и обитали в горных лесах. Так, К. А. Рассадина (1948, 1950) указывала, что большинство представителей Южной и Юго-Восточной Сибири, а также Приморского края относится к числу листоватых цетрарий (секция Platysma) и встречается главным образом на стволах и ветвях различных древесных пород. Можно предположить, что кустистые виды рода Cetraria - более поздние дериваты исходных листоватых форм. У ряда арктовысокогорных кустистых видов этой систематической группы лишайников прослеживаются генетические связи с лесными листоватыми формами. Например, К. А. Рассадина (1948) указывала на филогенетические связи арктовысокогорного вида Cetraria tilesii Ach. с эпифитным листоватым лишайником С. juniperina (L.) Ach., разновидностью которого он долгое время рассматривался.
Таким образом, как и в случае цветковых растений, у лишайников, по-видимому, также исходными типами многих ореофитов послужили микротермные и гелиофитные представители лесных третичных флор (Толмачев, 1958). Как отмечал А. И. Толмачев (1958), все ореофиты "альпийского типа" являются не только растениями, приспособленными к существованию в условиях минимального и ограниченного кратким периодом времени тепла, но также и гелиофитами. Причем наряду с ограниченной потребностью в тепле приверженность к местообитаниям, обеспечивающим достаточное освещение, была унаследована большинством ореофитов от их лесных предков. Возможно, как считал А. И. Толмачев, искони связанная с условиями неморального комплекса и порожденная ими "весенняя фракция" третичных неморальных флор дала начало ряду элементов высокогорных флор, складывавшихся первоначально как продукт преобразования неморального флористического комплекса в связи с поднятием гор.
II. Гипоарктомонтанный элемент. Объединяет 59 видов (7.8%), основное распространение которых связано с Гипоарктикой; после значительной дизъюнкции эти виды встречаются в хвойно-лесном поясе гор Голарктики (голарктический гипоарктомонтанный субэлемент) и других флористических царств (омнигипоарктомонтанный субэлемент).
Эта географическая группа лишайников охватывает представителей 16 семейств. Из них наиболее богаты по численности гипоарктомонтанных видов следующие семейства: Cladoniaceae (9 видов), Lecideaceae (8), Peltigeraceae, Parmeliaceae, Umbilicariaceae (пo 5 каждое), Physciaceae (4 вида).
А. Голарктический гипоарктомонтанный субэлемент. Охватывает 45 видов. Этим видам свойственны следующие типы ареалов.
1. Американо-азиатский тип ареала характерен для 3 видов - Leptogium crenulatum (Nyl.) Vain., Lasallia pensylvanica (Hoffm.) Llano, Umbilicaria muehlenbergii (Ach.) Tuck.
Распространение Leptogium crenulatum (Nyl.) Vain, связано главным образом с районом Берингова пролива, где он встречается на островах, и северным побережьем Чукотского полуострова (Vainio, 1909). Находка этого вида в Монголии, в окрестностях оз. Хубсугул, является первой находкой в горных районах Азии. Этот лишайник был собран на западном побережье оз. Хубсугул, к северу от Хатгала, на косе Модот-Булан, на высоте 1650 м над ур. м., где обитал на мхах на прибрежном лугу у леса (Голубкова, 1981; Цогт, 1981).
Два других вида - Lasallia pensylvanica (Hoffm.) Llano и Umbilicaria muehlenbergii (Ach.) Tuck. - широко распространены на территории как Азии, так и Северной Америки. Так, Lasallia pensylvanica (Hoffm.) Llano довольно часто встречается от арктической Сибири, Чукотского полуострова до гор Южной Сибири и Монголии и от Урала до Дальнего Востока и Японии. В Западной Сибири этот лишайник приводится для Тюменской и Томской областей (Семенов, 1922; Савич, 1926), для хр. Кузнецкий Алатау (Баранов, Смирнов, 1931; Седельникова, 1978). Он известен также из горных районов Восточной Сибири - для Восточного Саяна (Еленкин, 1902, 1903а, 19046), Прибайкалья (Рассадина, 1936) и Забайкалья (Окснер, 1926), Читинской области (Zahlbruckner, 1911), довольно широко распространен на Северо-Востоке СССР - в Восточной Якутии (Окснер, 1939а), в Магаданской области (Локинская, 1970), на Чукотском полуострове (Локинская, 1970) и на Анадыре (Рассадина, 1934). На Дальнем Востоке этот вид приводится для Амурской области (Еленкин, 1901 в) и Приморского края (Еленкин, 1912), известен также в Японии. На территории Монголии проходит, видимо, один из участков южного предела его распространения на территории Азии. В Монголии L. pensylvanica (Hoffm.) Llano - редкий лишайник, найденный на южном склоне Хангайского хребта, в долине р. Худжирт-Гол, где он обитал в лиственничном лесу, на высоте 2100 м над ур. м., а также в Монгольском Алтае, в долине р. Кобдо (Голубкова, 1981). В западной части ареала инвазии этого вида отмечены на территории европейской части СССР, в Ульяновской области (Еленкин, 1904а); он известен также на Кавказе. В Северной Америке основное распространение L. pensylvanica (Hoffm.) Llano связано с восточными штатами США - горами Аппалачи (Llano, 1950), а также с Аляской и западными районами Канады - Северо-Западными Территориями (Llano, 1950; Krog, 1968; Thomson е. а., 1969; Thomson, 1970, 1979; Bird е. а., 1980). В других районах этого континента имеются его изолированные местообитания: в США - в штатах Северная Дакота, Аризона, Калифорния, а также в Мексике (Llano, 1950). L. pensylvanica (Hoffm.) Llano известен также в западной Гренландии (Llano, 1950). Западно-арктическо-аппалачская дизъюнкция ареала этого лишайника (Brid е. а., 1980) свидетельствует о древнем, доледниковом его поселении в пределах Северной Америки. Его третичный возраст подтверждает как приуроченность основной части его ареала к восточным районам Северной Америки, так и широкое распространение его в горных районах Южной Сибири и Дальнего Востока.
Основной ареал Umbilicaria muehlenbergii (Ach.) Tuck, также связан с арктическими и горными районами Азии. Этот вид приводится для арктической Азии (Stizenberger, 1876), восточной Якутии (Окснер, 1939а; Куваев, 1956), Магаданской области (Андреев, 1978), западной Чукотки (Локинская, 1970). Южнее он известен в горных районах Казахстана, в хр. Кузнецкий Алатау (Баранов, Смирнов, 1931; Седельникова, 1978), в Красноярском крае (Vainio, 1896), Саянах (Еленкин, 1904; Савич, 1936), Прибайкалье (Савич, 1930; Рассадина, 1936), Забайкалье (Еленкин, 1907; Zahlbruckner, 1911). На Дальнем Востоке этот лишайник указан для побережья Охотского моря (Nylander, 1888) и Приморского края (Еленкин, 1912; Томин, 1926). В Монголии Umbilicaria muehlenbergii (Ach.) Tuck. - редкий лишайник. Он был найден в Монгольско-Даурском районе, в долине р. Онон, на северном склоне хребта Эрэн-Даба-Нуру, на скалах в сосновом лесу, а также в районе Средней Халхи, на сланцевых россыпях в долине р. Элдык (Голубкова, 1981). Видимо, в этих районах проходит южная граница его ареала в пределах Азии. На территории Северной Америки U. muehlenbergii (Ach.) Tuck, широко распространен в восточных районах Канады и США (Llano, 1950); в западных районах Северо-Западных Территорий и на Аляске имеются его изолированные местонахождения (Llano, 1950; Krog, 1968). Обширный дизъюнктивный ареал этого вида, его широкое распространение в горных районах Южной Сибири и Дальнего Востока и приуроченность основной части ареала к восточным районам Северной Америки дают основание предполагать третичный возраст этого лишайника.
2. Голарктический тип ареала свойствен 41 виду, обитающему в гипоарктической зоне и лесном поясе гор Голарктики. В составе этой географической группы виды родов Peltigera - P. aphthosa (L.) Willd., P. leucophlebia (Nyl.) Gyeln., P. collina (Ach.) Schrad., P. venosa (L.) Baumg.; Cladonia - C. bacilliformis (Nyl.) Gliick., C. polycarpoides Nyl., C. acuminata (Ach.) Norrl., C. norrlinii Vain.; Hypogymnia - H. bitteri (Lynge) Ahti, H. austerodes (Nyl.) Ras. (рис. 13, 1); Parmelia - P. disjuncta Erichs., P. infumnta Nyl., Leptogiurn tenuissimum (Dicks.) Koerb., Xylographa abietina (Pers.) Zahlbr., Rinodina conradii Koerb., Arthrorhaphis citrinella (Ach.) Poelt., Hymenelia lacustris (With.) Choisy, Aspicilia caesiocinerea (Nyl.) Arnold, Verrucaria imbrinula Nyl., V. aethiobola Wahlenb., Physconia mustigena (Ach.) Poelt, Cetraria juniperina (L.) Ach. и др.
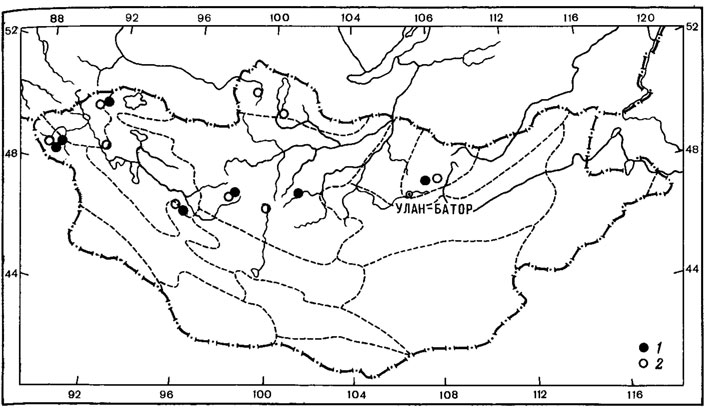
Рис. 13. Распространение на территории Монголии Hypogymnia austerodes (Nyl.) Ras. (1) и Pachyospora verrucosa (Ach.) Massal. (2). (Ориг.)
Б. Омнигипоарктомонтанный субэлемент. Объединяет 14 видов. Им присущ один тип ареала.
1. Мультирегиональный тип ареала характеризует лишайники, обитающие в гипоарктической зоне и лесном поясе гор северного и южного полушарий. Среди них можно назвать некоторые виды рода Cladonia - С. amaurocraea (Flk.) Schaer., С. deformis (L.) Hoffm., С. macroceras (Delise) Ahti, C. cornuta (L.) Hoffm.; Cladina stellaris (Opiz) Brodo, Icmadophila ericetorum (L.) Zahlbr., Lecidea vorticosa (Flk.) Koerb., Hypogymnia vittata (Ach.) Gas., Parmelia saxatilis (L.) Ach., Umbilicaria polyphylla (L.) Baumg. и др.
Таким образом, среди гипоарктомонтанного элемента лихенофлоры Монголии преобладают виды с голарктическими ареалами (42 вида, или 71.1% от общего числа видов этого элемента). Ареалами, простирающимися за пределы Голарктики, характеризуется 14 видов (23.7%). Более 5% видов имеет более узкие американо-азиатские ареалы.
Гипоарктомонтанные лишайники широко распространены на территории Монголии, причем большинство их приурочено к горным районам, к Прихубсугулью, Хэнтэю, Хангаю, Монгольскому Алтаю, где они часто встречаются в высокогорном поясе в составе высокогорных степей, кобрезиевых пустошей, горных тундр.
Однако характерной особенностью растительного покрова Монголии является широкое участие гипоарктомонтанных видов лишайников в составе лесных, а также степных ценозов, при этом некоторые из них играют определенную фитоценотическую роль в лесных и степных сообществах. К числу таких видов, например, относится Hypogymnia bitteri (Lynge) Ahti. Этот листоватый лишайник является одним из доминантных видов эпифитных лишайниковых синузий, развивающихся на стволах лиственниц, кедров и других пород деревьев в лиственничных, кедрово-лиственничных и других типах лесов Монголии (Бязров, 1974а, 1974б, 1976, 1977). В составе эпигейных синузий лиственничных лесов заметная роль принадлежит видам рода Peltigera, в их числе и гипоарктомонтанному виду Peltigera leucophlebia (Nyl.) Gyeln. (Бязров, 1974a), а в сосновых лесах Монгольско-Даурского района - гипоарктомонтанному виду Cladonia amaurocraea (Flk.) Schaer. Некоторые виды гипоарктомонтанного элемента принимают заметное участие в степных сообществах Монголии. Среди них прежде всего следует назвать листоватый лишайник Physconia mustigena (Ach.) Poelt. Этот вид чрезвычайно широко распространен на замшевелой почве, дерновинках мхов, растительных остатках у основания и в расщелинах скал в различных степных и высокогорнолуговых ценозах в Прихубсугулье, Хангае, Монгольском Алтае, а также в Котловине Больших Озер и Гобийском Алтае, в пределах высот 1200 - 2970 м над ур. м. В составе эпилитных синузий горно-степных ценозов в Монгольско-Даурском районе и в Восточной Монголии заметную роль играет накипной гипоарктомонтанный лишайник Aspicilia cupreoatra (Nyl.) Arnold. Время вхождения гипоарктомонтанного элемента в состав флоры лишайников Монголии, видимо, можно отнести к плейстоцену.
III. Бореальный элемент. В понимании этого элемента мы придерживаемся мнения X. X. Трасса (1970), который к бореальному элементу относил виды, распространенные (и имеющие центры массовости) в бореальной зоне хвойных лесов Голарктики, и, кроме того, бореальные виды, поднимающиеся в горы (даже до альпийского пояса). Специфические природные особенности Монголии, ее высокое положение над уровнем океана, приуроченность лесной растительности к горным поднятиям - все это способствовало расселению в ее пределах видов горно-бореального распространения. По своему видовому составу бореальный элемент лихенофлоры Монголии имеет много общего с видовым составом горно-бореальных групп распространения бореального и мультирегионального элементов, выделяемых М. Ф. Макаревич (1963) при анализе лихенофлоры Украинских Карпат. Характеризуя горно-панбореальную группу видов, М. Ф. Макаревич отмечала, что в этой группе наиболее выявляются как характерные закономерности распространения бореальных видов, так и экологические особенности их. Распространившись в бореальной зоне Голарктики и нередко заходя в южную часть Арктики, они везде встречаются в соответствующих равнинных условиях, поднимаясь в горах иногда и до альпийского пояса. В основном это виды хвойных лесов и их дериватов, которые поселяются на коре и древесине хвойных деревьев и их спутников, песчаных грунтах бореальных пустошей, на болотах, часто на мхах и растительных остатках, нередко на обработанной и гниющей древесине. К бореальному элементу мы вслед за X. X. Трассом (1970) присоединяем также виды лишайников, встречающихся, помимо Голарктики, в холодно- умеренных областях южного полушария, т. е. виды нотобореального элемента в понимании А. Н. Окснера (1944). Виды лишайников с подобным распространением мы объединяем в омнибореальный субэлемент.
В систематическом отношении лишайники бореального элемента относятся к 20 семействам. Наибольшее число бореальных видов во флоре Монголии насчитывают следующие семейства: Cladoniaceae (28 видов), Usneaceae (22), Lecideaceae (18), Physciaceae (13), Parmeliaceae (10), Caliciaceae (7), Peltigeraceae (6 видов), Stereocaulaceae и Nephromataceae (пo 5 каждое). Бореальный элемент охватывает 145 видов (19.2%).
А. Голарктический бореальный субэлемент объединяет 82 вида. Этим видам свойственны следующие типы ареалов.
1. Сибирско-монгольский тип ареала присущ 2 видам - Rinodina sibirica Н. Magn., R. exiguella (Vain.) H. Magn. (рис. 14), растущим в хвойных лесах Сибири и северных районах Монголии.
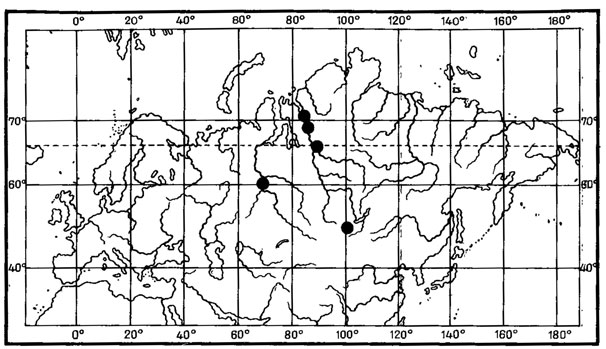
Рис. 14. Распространение на земном шаре Rinodina exiguella (Vain.) H. Magm. (Ориг.)
Как отмечал Магнуссон (Magnusson, 1947), R. sibirica Н. Magn. (рис. 15), обитающая на гладкой коре деревьев лиственных и хвойных пород, по-видимому, является широко распространенным видом на территории Сибири; в исследованных районах этот вид довольно обычен. Западная часть ареала этого вида приурочена к Западно-Сибирской низменности, к долинам рек Оби, Иртыша и Конды. В Восточной Сибири этот лишайник известен из многочисленных местонахождений вдоль долины р. Енисея, от г. Енисейска до Дудинки, причем его встречаемость по направлению к северу постепенно падает, в Якутии, в долине р. Лены и в устье р. Алдана, в Иркутской области в Восточном Саяне (Magnusson, 1936b, 1947). На территории Монголии был найден в Хэнтэе, в хр. Барун-Борхын-Нуру, где обитал в кедрово-лиственничном лесу, а также в долине р. Улдза в березовом лесу, в Монгольско-Даурском районе (Голубкова, 1981).
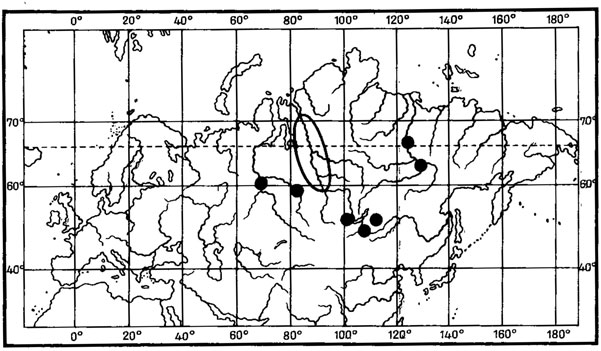
Рис. 15. Распространение на земном шаре Rinodina sibirica H. Magn. (Ориг.)
Распространение R. exiguella (Vain.) Н. Magn., как и R. sibirica Н. Magn., также, видимо, недостаточно хорошо изучено. Этот вид известен на территории Западно-Сибирской низменности, в долинах рек Иртыша и Конды и в Восточной Сибири, в нижнем течении р. Енисея (Vainio, 1928; Magnusson, 1947); в Монголии он был найден в северном Хангае (Голубкова, 1981).
Эти азиатские виды рода Rinodina в систематическом отношении относятся к секции Eurinodina Stiz., подсекции Pachysporina Н. Magn., к одной и той же серии - Archaeinae Н. Magn., представители которой широко распространены в, хвойных лесах Сибири и Монголии. Характер ареалов этих видов, основная часть которых приурочена в Сибири к районам, испытавшим в плейстоцене покровное оледенение, их слабая морфологическая обособленность ют других видов ринодин серии Archaeinae Н. Magn. - R. exigua (Ach.) S. Gray, R. jeniesejensis H. Magn. - дает основание предполагать относительную молодость этих видов.
2. Сибирско-восточноазиатский тип ареала характерен для 4 видов лишайников, основное распространение которых связано с восточноазиатской областью и восточносибирскими районами. В составе этой группы видов - Evernia esorediosa (Mull. Arg.i) DR., Hypogymnia fragillima (Hillm.) Rassad., H. iuplicatoides (Oxn.) Rassad., Cetraria komarovii Elenk.
Evernia esorediosa (Mull. Arg.) DR. - эпифитный лишайник, обитающий на стволах и ветвях лиственниц, кедров, пихт, иногда на деревьях других пород и кустарниках в лиственничных, кедрово-лиственничных, лиственничных лесах с примесью ели, березы, ольхи, сосново-березовых и в других типах леса. Ареал этого эпифитного лишайника охватывает Японию (о-в Хонсю), Дальний Восток, Бурятскую АССР, Читинскую область, Якутию - Лено-Алданский и Лено-Амгинский водоразделы, Верхоянский хребет, долину р. Индигирки, Иркутскую область - побережье оз. Байкал и Восточный Саян, Красноярский край, достигая на западе долины р. Енисея, а на севере - северной части Средне-Сибирского плоскогорья, гор Путорана (Рассадина, 1936; Савич, 1936; Будаева, 1976; Афоноина и др., 1979, и др.; Гербарий БИН АН СССР). Он приводится также для Северо-Восточного Китая :(Флоров,ская, 1962). В Монголии Е. esorediosa (Mull. Arg.) DR. встречается только в районе Прихубсугулья, по берегам и на островах оз. Хубсугул, где обитает в лесном поясе, иногда на его крайних пределах, на ветвях лиственниц и кедров. Этот вид указывался Крог (Krog, 1962, 1968) для Аляски, однако, как отмечал Бирд (Bird, 1974), материал, на котором было основано это сообщение, в действительности относится к Е. perfragilis Llano. В систематическом отношении Е. esorediosa (Mull. Arg.) DR. относится к секции Archevernia Th. Fr. Вместе с другими представителями этой секции - Е. mesomorpha Nyl., Е. perfragilis Llano, Е. terrestris (Tomin) Golubk. - этот вид образует один систематический ряд близкородственных видов - Esorediosa Flor. Центральное место в этом ряду занимает Е,esorediosa (Mull. Arg.) DR., который, по мнению Е. Ф. Флоровской (1952), является наиболее древним. Ею высказано предположение, что в третичный период род Evernia был представлен несколькими видами, из которых наиболее древними являются Е. divaricata (L.) Ach., Е. esorediosa (Mull. Arg.) DR., E. prunastri (L.) Ach. О древнем происхождении E. esorediosa (Miill. Arg.) DR. свидетельствует прежде всего примитивный способ размножения - с помощью спор, ряд примитивных анатомо-морфологических признаков, а также его ареал, основная часть которого связана с Восточноазиатской областью и горными районами Юго-Восточной Сибири, с районами, богатыми третичными реликтами.
Cetraria komarovii Elenk. имеет дизъюнктивный ареал, одна часть которого связана с горами Южной Сибири - с Восточным Саяном (Тунктинский хребет, Тунктинская долина, хр. Пограничный), Становым нагорьем (Верхне-Ангарский и Байкальский хребты), Забайкальем (Малханский хребет, долина р. Булукун, Баргузинский хребет), на территории МНР - с горами Прихубсугулья и Хангая (Голубкова, 1981; Макрый, 1981; Будаева, 1982). Вторая часть ареала этого вида приурочена к Восточноазиатской области Приморскому краю и хр. Сяоу-Тайшань (Гурулева, Княжева, 1972; Княжева, 1973; Макрый, 1981). В Монголии этот лишайник является редким и обитает на замшелых скалах, однако, как отмечала Т. В. Макрый (1981), он растет также на стволах деревьев, на поверхности камней и почве в лесном поясе, иногда поднимаясь в высокогорья, при этом предпочитает хорошо увлажненные, прогреваемые местообитания в долинах горных рек и ручьев с более мягким микроклиматом. Этот вид восточно-азиатских филогенетических связей, он относится к секции Platysma Koerb. рода Cetraria, представители которой - эпифитные листовые формы - встречаются главным образом в Восточной и Юго-Восточной Азии (Рассадина, 1950). Наиболее близкородственные виды этого лишайника - С. wallichiana Mull. Arg. и С. pseudocomplicata Asah. - восточного и юго-восточного азиатского распространения.
Основное распространение Hypogymnia duplicatoides (Oxn.) Rassad. связано с Японией и Дальним Востоком. На территории СССР он был найден в Приморском крае, на берегу залива Посьет, в южной части хр. Сихотэ-Алинь (Рассадина, 1956), в Хабаровском крае в долине р. Мухень, притока р. Садома (Окснер, 1938). После значительной дизъюнкции этот вид встречается в Монголии, в Хангае, где является редким. Он обитает в лиственничных и кедрово-лиственничных лесах среднегорного пояса, на коре деревьев (Бязров и др., 1983).
Основной ареал Н. fragillima (Hillm.) Rassad. также связан с Восточноазиатской областью - с Японией, п-овом Корея, о-вом Сахалин, Северо-Восточным Китаем (провинция Хэйлунцзян), Дальним Востоком, где был найден на восточном склоне хр. Сихотэ-Алинь, в высокогорном ельнике в верховьях р. Талеш (Рассадина, 1953, 1956). На территории МНР имеется изолированное местообитание этого вида в Хангае. Здесь он был собран лишь однажды в верховьях р. Хух-Сум, на склонах северной экспозиции, в кедрово-лиственничном лесу, на высоте 2000 м над ур. м., на коре лиственницы (Бязров и др., 1983). Изолированные местообитания как этого вида, так и Н. duplicatoides (Oxn.) Rassad. на территории Монголии, видимо, носят реликтовый характер.
3. Евразиатский тип ареала присущ 12 видам - Buellia pharcidia (Ach.) Malme, B. margaritacea (Sommerf.) Lynge, Aspicilia verrucigera Hue, A. laevata (Ach.) Arnold, Cyphelium notarisii (Tul.) Blom. et Forss., Rinodina cinereovirens (Vain.) Vain, и др. Эти виды характеризуются дизъюнктивными ареалами, охватывающими подзону хвойных лесов и лесной пояс гор Европы и Азии.
4. Голарктический тип ареала характерен для видов с обширными ареалами, одна часть которых расположена в хвойнолесной подзоне, другая - в лесном поясе гор Голарктики. Эту группу лишайников составляют 69 видов; ряд видов родов Bryoria - В. furcellata (Fr.).Brodo et D. Hawksw., В. simplicior (Vain.) Brodo et D. Hawksw., B. implexa (Hoffm.) Brodo et D. Hawksw., B. capillaris (Ach.) Brodo et D. Hawksw., B. trichodes (Michx.) Brodo et D. Hawksw.; Usnea - U. filipendula Stirt., U. glabrescens (Nyl.) Vain., U. longissima Ach., U. glabrata (Ach.) Vain., U. hirta (L.) Wigg., Buellia - B. schaereri DNot., B. zahlbruckneri Steiner, B. triphragmoides Anzi; Spilonema revertens Nyl.; Parmelia - P. olivacea (L.) Ach. (рис. 16), P. septentrionalis (Lynge) Ahti, P. exasperatula Nyl., P. subargentifera Nyl.; Biatora - B. uliginosa (Schrad.) Fr., B. fusca (Borr. ex Schaer.) Th. Fr., B. helvola Koerb., B. granulosa (Ehrh.) Flot., B. symmicta (Ach.) Fr.; Cladonia - C. anomaea (Ach.) Ahti et P. James, C. turgida (Ehrh.) Hoffm., C. grayi Merr., и др.; Lecanora - L. piniperda (Boist.) Koerb., L. saligna (Schrad.) Zahlbr.; Hypogymnia - H. bitteriana (Zahlbr.) Ras., H. tubulosa (Schaer.) Hav.; Bilimbia - В. naegelii (Hepp) Krempelh., В. microcarpa (Th. Fr.) Lett.; Leptorhaphis epidermidis (Ach.) Th. Fr., Catillaria atropurpurea (Schaer.) Th. Fr., Scoliciosporum chlorococcum (Stenh.) Vezda, Evernia mesomorpha Nyl., E. divaricata (L.) Ach., Parmeliopsis ambigua (Wulf.) Nyl., Acarospora oligospora (Nyl.) Arnold, Bacidia inundata (Fr.) Koerb. и др.
Рис. 16. Распространение на земном шаре Parmelia olivacea (L.) Ach. (По: Ahti, 1966)
В качестве примера рассмотрим распространение некоторых видов рода Bryoria. Так, например, основное распространение В. simplicior (Vain.) Brodo et D. Hawksw. связано с северными лесами Европы, Северной Америки и, вероятно, Азии, где, однако, его ареал недостаточно изучен. Этот эпифитный лишайник обитает главным образом на коре деревьев хвойных пород (елях, лиственницах, пихтах и др.), иногда и на деревьях других пород (Ahlner, 1948; Poelt, 1969; Brodo, Hawksworth, 1977). Изредка этот вид проникает в субарктическую зону и даже в Арктику, где растет на скалах; так, например, он приводится для юго-западных районов Гренландии (Dahl, 1950). В Северной Америке В. simplicior (Vain.) Brodo et D. Hawksw. встречается в бореальной и субарктической зонах, от Ньюфаундленда до Аляски; южнее обитает в горах западных районов, проникая на юг до гор Колорадо, а также в горных районах Квебека (Brodo, Hawksworth, 1977). На территории Азии, помимо Монголии, приводится для Сибири, Камчатки и Японии (Ahlner, 1948). В Монголии этот вид довольно широко распространен в районе Прихубсугулья и в Хангае (Голубкова, 1981), где встречается в лиственничных лесах на коре лиственниц, гниющем дереве, иногда на скалах.
Некоторые виды этой группы лишайников вместе с горными хвойными лесами проникают далеко на юг, в южные районы Голарктики. Например, В. furcellata (Fr.) Brodo et D. Hawksw. часто встречается в хвойных лесах Европы, особенно в сосновых (Ahlner, 1948; Poelt, 1969), но в то же время приводится для о-ва Мадейра (J0rgensen, 1972). В Северной Америке он обычен в северной и средних частях хвойнолесной подзоны, от Ньюфаундленда до Аляски, но особенно обилен в районе Великих озер и в горах Аппалачи (Brodo, Hawksworth, 1977). На западе Северной Америки имеются изолированные местонахождения этого лишайника в Скалистых горах, а также в некоторых других горных районах США. Южнее он проникает в высокогорьях Мексики, а также в Центральную Америку в горы Коста-Рики. Его ареал на территории Азии недостаточно изучен. До его находок в МНР этот вид был известен в Гималаях (Bystrek, 1969) и Японии (Kurokawa, 1959). На территории Монголии один из широко распространенных видов бриорий: довольно широко встречается в Прихубсугулье, Хангае и Монгольско-Даурском районе (Голубкова, 1981), где обитает в лиственничных, сосновых и березовых лесах на коре деревьев.
Б. Омнибореальный субэлемент. Объединяет 59 видов лишайников, которым присущ один тип ареала.
Мультирегиональный тип ареала характерен для видов с обширным дизъюнктивным ареалом, одна часть которого расположена в подзоне хвойных лесов Голарктики, другая - в холодно-умеренных широтах южного полушария. В составе этой географической группы многие виды рода Cladonia - С. digitata (L.) Hoffm., С. coccifera (L.) Willd., С. pleurota (Flk.) Schaer., C. bacillaris (Ach.) Nyl., C. cariosa (Ach.) Spreng., C. phyllophora Hoffm., C. gracilis (L.) Willd., C. botrytes (Hagen) Willd. и др., Cladina rangiferina (L.) Harm., Hypogymnia physodes (L.) Nyl., Buellia punctata (Hoffm.) Massal., Stereocaulon paschale (L.) Hoffm., S. condensatum Hoffm., S. glareosum (Savicz) H. Magn., Peltigera spuria (Ach.) DC., P. malacea (Ach.) Funck, Biatora turgidula (Fr.) Nyl., B. vernalis (L.) Fr., B. symmictera (Nyl.) Ras., Lecanora varia (Ehrh.) Ach., L. poiytropa (Ehrh.) Rabenh., Catillaria sphaeroides (Dicks.) Schui., Cornicularia acu- leata (Schreb.) Ach., Cetraria pinastri (Scop.) S. Gray, Calicium abietinum Pers., C. trabinelium Ach., C. sphaerocephalum (L.) Ach., Coniocybe furfuracea (L.) Ach., Leptogium saturninum (Dicks.) Nyl., Nephroma helveticum Ach., N. laevigatum Ach., Buellia geophila (Flk.) Lynge, Bryoria fuscescens (Gyeln.) Brodo et D. Hawksw., B. nadvornikiana (Gyeln.) Brodo et D. Hawksw. и др.
В этой группе лишайников представляют интерес виды рода Nephroma. Этот род содержит около 45 видов, распространенных по всему земному шару, главным образом в умеренных и горных областях (Окснер, 1956). На территории Монголии встречаются 5 видов этого рода, но большинство из них редкие в этом районе лишайники. В этом отношении исключение составляет Н. helveticum Ach. (рис. 17). Этот вид довольно широко распространен в горно-таежном поясе в районе Прихубсугулья, а также в Хэнтэе, Хангае, Монгольско-Даурском районе, где обитает в лиственничных и березовых лесах на замшелых скалах, гниющих стволах деревьев, мхах. В Азии N. helveticum Ach. встречается на побережье оз. Байкал, в Забайкалье, Саянах, в Красноярском крае, на Алтае, в Томской области, а также на Камчатке, п-ове Корея, в Японии, Китае, Индии, Непале, на о-ве Ява (Домбровская, 1975). За пределами Азии ареал этого вида охватывает хвойные леса и горы Европы (Фенноскандия, Франция, Италия, ФРГ, Балканский полуостров, Венгрия), на территории европейской части СССР - Карелию, Архангельскую область, Эстонию, Кировскую область, а также Урал и Кавказ (Окснер, 1956). Кроме того, этот лишайник встречается в Северной и Центральной Америке, в Новой Зеландии.
Рис. 17. Распространение на территории Монголии Nephroma helveticum Ach. (I) и N. isidiosum Ach. (2). (Ориг.)
Второй бореальный вид рода - Nephroma laevigatum Ach, - является редким на территории Монголии лишайником. Он был найден только в Хангае (Бязров и др., 1983), где обитает на основаниях стволов деревьев, валеже и скалах в лесах среднегорного пояса. Ареал этого вида охватывает хвойнолесную область и горы Европы (Фенноскандия, Великобритания, Франция, Италия, Австрия, ФРГ, Балканский полуостров, Чехословакия, Венгрия, Польша) (Окснер, 1956). На территории европейской части СССР этот лишайник обычен в Карелии, Архангельской и Ленинградской областях, Эстонии, Белоруссии, в Карпатах и Крыму, в Кировской области, на Урале. В Азии он известен в Омской области, на Алтае, в Прибайкалье, на Камчатке, в Японии. В Северной Америке N. laevigatum Ach. ведет себя как океанический вид и встречается главным образом вдоль Тихоокеанского побережья, от Аляски, Британской Колумбии до Калифорнии, и на Атлантическом побережье - от п-ова Лабрадор до штата Массачусетс (Wetmore, 1960). Кроме того, этот вид растет в горах Северной Африки (Алжир, Тунис, Марокко), а также на Канарских островах и о-ве Мадейра, в Южной Америке, на о-ве Ява, в Австралии и Новой Зеландии.
В Монголии бореальные виды лишайников широко представлены в лесных ценозах в Прихубсугулье, Хэнтэе, Хангае, Монгольском Алтае и в Монгольско-Даурском районе. При этом в лиственничных, кедрово-лиственничных, сосновых и березовых лесах они играют заметную фитоценотическую роль, доминируя в составе эпифитных, эпилитных, эпигейных и эпиксильных лишайниковых синузий. В числе таких видов можно назвать Hypogymnia physodes (L.) Nyl. (f. labrosa Ach.), Evernia mesomorpha Nyl., часто встречающихся на коре лиственницы, кедра, березы, сосны в горнотаежном поясе и поясе леса и степи горных районов Монголии. В составе эпигейных лихеносинузий заметное участие принимают бореальные виды родов Cladonia - С. digitata (L.) Hoffm., С. саrriosa (Ach.) Spreng., C. phyllophora Hoffm., C. gracilis (L.) Willd. и др.; Peltigera - P. malacea (Ach.) Funck, P. spuria (Ach.) DC. ap. Lam. et DC., P. praetextata (Flk.) Vain, и другие виды. В составе эпиксильных лишайниковых синузий - виды рода Biatora - В. symmicta (Ach.) Fr., В. turgidula (Fr.) Nyl. и др., а также Hypogymnia physodes (L.) Nyl. Некоторые виды бореальных лишайников, например Cornicularia aculeata (Schreb.) Ach., С. odontella (Ach.) Rohl., часто встречаются на почве среди мхов и других растений в горных степях, поднимаясь до высокогорного пояса, где достигают широкого развития на кобрезиевых пустошах. Ряд видов этого элемента заходит в горные и сухие степи Монголии. Например, Spilonema rеvertens Nyl. является одним из типичных лишайников горных степей Прихинганья, где образует на скалах очень характерное сообщество с Psorula rufonigra (Tuck.) G. Schneider. Этот чешуйчатый лишайник обычно развивается на дерновиках карликово-кустистого слоевища слизистого лишайника Spilonema revertens Nyl. В степных районах Восточной Монголии на поверхности мелких камней щебнисто-галечного панциря довольно часто встречается бореальный лишайник Acarospora oligospora (Nyl.) Arnold.
Таким образом, как можно было видеть, в составе бореального элемента преобладают виды, характеризующиеся циркумполярным распространением в Голарктике, - 46.8% видов бореального элемента, с мультирегиональными ареалами - 40.8% видов. На долю видов с евразиатскими ареалами приходится 8.2%, с азиатским - всего 4.1%.
Виды бореального элемента очень разнообразны в историческом и генетическом отношении. Как отмечала М. Ф. Макаревич (1963), генезис бореальных видов очень сложный и географическая дифференциация этого элемента относится к глубокой древности. Вслед за А. Н. Окснером и А. С. Лазаренко она полагает, что родину древних бореальных видов следует искать на северо-востоке Азии и северо-западе Северной Америки. К числу представителей древней ранней бореальной флоры А. Н. Окснер (1944), например, относил Cornicularia aculeata (Schreb.) Ach., Parmeliopsis aleurites (Ach.) Nyl. В числе молодых бореальных видов можно назвать Evernia mesomorpha Nyl., который, как полагает Н. А. Миняев (1940), сформировался, видимо, в начале четвертичного периода, а также сибирско-монгольские виды Rinodina sibirica Н. Magn., R. exiguella H. Magn. Как полагал Хультен (Hulten, 1937), значительное ядро современной евразиатской бореальной флоры имело максимальное распространение в эпоху "великого межледниковья" (миндель - рисс), когда ареалы многих растений этого типа простирались непрерывно от "Великого" до Атлантического океанов. Однако во время "великого оледенения" (рисс) эти ареалы уже не представляли непрерывного пояса, а были составлены из целого ряда изолированных, элементарных ареалов. Как полагает Хультен, по мере отступания льда радианты из этих ареалов начали расселяться и заполнили прорывы, другие же формы по каким-то причинам не успели заполнить разрывы ни в рисс-вюрм, ни в послеледниковое время, поэтому мы имеем самые разнообразные конфигурации дизъюнкции. Но так как при этом происходили не только дефект ареалов, но и морфологическая дифференциация различного объема, часто хорошо подчеркнутая продолжительной географической изоляцией, то возникали многочисленные примеры близкородственных систематических рядов или видовых серий. Примером систематического ряда близкородственных видов, возникших, видимо, в результате морфологической дифференциации единого предкового бореального вида и распада его ареала в ледниковые фазы плейстоцена являются азиатские систематически близкие виды рода Rinodina - R. exiguella Н. Magn., R. sibirica H. Magn., R. jenisejensis H. Magn. В силу экологической пластичности лишайников процесс восстановления ареалов бореальных лишайников в послеледниковье, видимо, происходил довольно активно. Об этом свидетельствует хотя бы такой факт, что среди представителей бореального элемента в лихенофлоре Монголии преобладают, как было показано выше, виды с циркумполярным распространением в Голарктике. Как отмечал Ахти (Ahti, 1977), циркумполярному расселению лишайников в зоне хвойных лесов способствуют относительное однообразие экологических условий, слабое влияние флорогенетических изоляций и замедленное протекание эволюционных процессов компонентов флоры.
В генетическом отношении бореальный элемент не представляет однородного комплекса. В формировании бореального элемента лихенофлоры Монголии, очевидно, значительная роль принадлежит восточноазиатскому центру. В частности, с районами Восточной Азии связан генезис многих видов рода Hypogymnia, представители которого играют важную фитоценотическую роль в хвойных лесах Евразии, в том числе и в Монголии. Как указывалось выше, в лесных ценозах Монголии такие виды, как Н. bitteri (Lynge) Ahti и Н. physodes f. labrosa Ach., входят в число доминантов в составе эпифитных лишайниковых синузий, развивающихся на стволах лиственниц, кедров, сосен, елей и на деревьях других пород. Как отмечала К. А. Рассадина (1965а), представители рода Hypogymnia тесно связаны с хвойными породами деревьев и в изобилии покрывают стволы и ветви елей, пихт, лиственниц, часто переходя на березу, а также на другие субстраты, кору лиственных пород деревьев, обработанное дерево, почву, каменистые субстраты. По ее данным (Рассадина, 1965а), этот род насчитывает 58 видов, из них более половины - 32 вида - обитает в Азии, причем наибольшее число видов встречается на Дальнем Востоке, в Китае и Японии. 17 видов являются эндемиками азиатской флоры. Остальные виды рода Hypogymnia распространены на земном шаре следующим образом. Во флоре Северной Америки известно 17 видов гипогимний, из них 4 эндемичны для нее; в австралийской флоре - 16 видов, из них 11 - ее эндемики. В Европе встречается 11 видов этого рода, из них лишь один вид эндемичен для европейской флоры. В Африке, главным образом тропической, известны 7 видов гипогимний, 6 из них - эндемики, в Южной Америке - 4 вида, все они эндемичны для этого континента. К. А. Рассадина также указывала, что 15 видов рода Hypogymnia, характеризующиеся более обширными ареалами, проявляют связь с флорой Азии. Обширный ареал рода, охватывающий все континенты земного шара, свидетельствует о его древней дифференциации. На основании морфологического строения слоевища К- А. Рассадиной (1967) была предпринята попытка сгруппировать виды рода Hypogymnia в филогенетические ряды. В зависимости от родственных связей видов они были объединены ею в несколько секций, относимых к двум под- родам Tubulosae (Bitt.) Krog и Solidae (Bitt.) Krog. Анализируя состав и географическое распространение представителей этих секций, можно заметить, что с районами Восточной Азии связано распространение преимущественно подрода Tubulosae (Bitt.) Krog - секций Delavayi Rassad. и Metaphysodes Rassad. К представителям этих секций относятся главным образом эпифитные формы, характеризующиеся наиболее примитивным строением слоевища из несомкнутых, вытянутых лопастей и рыхлой сердцевиной, имеющей внутреннюю полость. Третья секция этого подрода - Subobscura Rassad. - объединяет филогенетически тесно связанные друг с другом гипоарктомонтанные - Н. bitteri (Lynge) Ahti, Н. austerodes (Nyl.) Ras и арктовысокогорный - H. subobscura (Vain.) Poelt - виды, имеющие циркумполярное распространение в Голарктике. Представители второго подрода - Solidae (Bitt.) Krog, характеризующиеся сердцевиной плотного строения, преимущественно эпилитные арктовысокогорные формы циркумполярного, евразиатского и амфиберингийского распространения. Как отмечала К. А. Рассадина (1965а), переход некоторых видов рода Hypogymnia к обитанию с древесного субстрата на каменистый способствовал расширению ареала ряда видов, и в целом рода, и их продвижению на север. Представители двух последних групп составляют основу видового состава гипогимний во флорах Европы и Северной Америки. Таким образом, Восточная Азия является не только областью наибольшего видового разнообразия рода Hypogymnia, но к этому району земного шара тяготеют и наиболее примитивные морфотипы этой группы лишайников. Все это дает основание искать центр формирования рода в районах Восточной Азии.
Тот факт, что центр современного видового многообразия рода находится в пределах современного ареала дальневосточных третичных лесов, позволяет высказать предположение, что исходные формы этого рода, возможно, были связаны с третичными лесами тургайского типа. Об этом также свидетельствуют экологические особенности многих видов гипогимний, о которых писала К. А. Рассадина (1967), а именно о их способности обитать на деревьях как хвойных, так и лиственных пород. Как отмечал Р. В. Камелин (1979), в составе тургайской флоры, по-видимому в горных условиях, где эта флора могла вобрать в себя и ряд глубокоархаичных типов, формировались не только неморальные, но и первичнотаежные группировки.
К числу дериватов тургайской флоры относятся также бореальные виды рода Evernia - Е. mesomorpha Nyl., Е. divaricata (L.) Ach. Как уже указывалось, этот род А. Н. Окснер (1946а) считал автохтонным тургайским родом, на который впоследствии оказал сильное воздействие процесс бореализации. Возникновение исходных форм рода датируется верхним мелом (Флоровская, 1952). К числу наиболее древних видов Е. Ф. Флоровская (1952) причисляет и Е. divaricata (L.) Ach. Исходным центром формирования этой группы лишайников, как и в случае рода Hypogymnia, видимо, была Восточная Азия. Из 13 видов еверний 8 встречаются в Азии, из них 6 - в Восточной Азии. С этим районом земного шара связано распространение таких древних форм этой систематической группы, как Е. esorediosa (Mull. Arg.) DR. (Флоровская, 1952). E. mesomorpha Nyl., по-видимому, является более поздним дериватом этой исходной тургайской формы. О близости этих двух видов свидетельствует значительное сходство в морфологическом строении их слоевищ. Однако в отличие от фертильного Е. esorediosa (Mull. Arg.) DR., размножающегося спорами и всегда образующего плодовые тела, Е. mesomorpha Nyl. размножается вегетативно, с помощью соредиев и изидиев. Эти два вида, вероятно, можно рассматривать как "парные виды", один (стерильный) из которых, как писал Пельт (Poelt, 1972), примыкает к другому (генеративному) как апомиктический дериват. Переход Е. mesomorpha Nyl. к вегетативному, более эффективному и быстрому, способу размножения, возможно, позволил этому виду расширить свой ареал, выйти за пределы Восточной Азии и широко расселиться в таежных лесах Сибири, а также в Европе и Северной Америке (Bird, 1974). Анализируя реликтовые элементы в современной флоре лишайников восточной Прибалтики, Н. А. Миняев (1940) указывал, что сопоставление ареалов видов лишайников сосновых боров убеждает в том, что они пришли с востока. В числе комплекса боровой флоры им названы Е. mesomorpha Nyl., а также такие бореальные виды, как Hypogymnia physodes (L.) Nyl., Cetraria pinastri (Scop.) S. Gray, Parmelia olivacea (L.) Ach. и др. Причем он отмечает, что этот комплекс включает виды очень древние и, наоборот, молодые, сформировавшиеся лишь в начале четвертичного периода, чрезвычайно активные, например Evernia mesomorpha Nyl.
Роль тургайского генетического элемента в современной бореальной флоре подчеркивалась М. Ф. Макаревич (1963, 1964). На основании анализа лихенофлоры Украинских Карпат 104 она заключила, что среди бореальных лишайников этого района довольно богато представлен "тургайский генетический элемент - виды, возникшие из обитателей широколиственных лесов, которые с конца олигоцена и в миоцене, распространяясь из Восточной Азии, постепенно завоевывали европейские пространства, занятые субтропической полтавской флорой (Криштофович, 1946, 1947), например эпибриофильные и литофилные виды рода Bacidia, род Bryopogon, Alectoria sarmentosa" (Макаревич, 1964, с. 27). К числу дериватов тургайской флоры, видимо, относятся многие виды рода Bryoria, который, вероятно, горно-тургайского происхождения, но впоследствии был охвачен процессом бореализации (Окснер, 1946а; Макаревич, 1963).
Характер распространения на земном шаре и генетические связи как отдельных видов бореального элемента, так и ряда систематических групп, в настоящее время широко представленных в подзоне хвойных лесов Голарктики, указывают на их тропогенезис. К числу таких групп относится род Cladonia, представители которого, как известно, играют заметную фито- ценотическую роль в составе хвойных, особенно сосновых лесов. Древность рода Cladonia не может вызывать сомнения (Макаревич, 1963). Этот полиморфный род насчитывает 280 видов, распространенных по всему земному шару, от Арктики до тропических районов и Антарктики (Окснер, 1968). Особенно богаты видами кладоний флоры тропических и субтропических областей, где концентрируется большинство эндемичных видов. При этом наибольшее число эндемичных видов этого рода приходится на тропическую и субтропическую Южную Америку. Из 280 видов этого рода только около 40 видов имеет очень широкое распространение на земном шаре, охватывающее северное и южное полушария, и достигает массовости в холодно- умеренных районах обоих полушарий. В числе этих видов и многие виды бореального элемента лихенофлоры Монголии, например Cladonia coccifera (L.) Willd., С. pleurota (Flk.) Schaer., C. bacillaris (Ach.) Nyl., C. phyllophora Hoffm., C. cenotea (Ach.) Schaer., C. uncialis (L.) Wigg.
В числе систематических групп лишайников, играющих заметную роль в составе ценозов хвойных лесов Голарктики, но в своем генезисе связанных с тропиками, может быть назван также род Peltigera. Этот род насчитывает 70 видов, широко распространенных на земном шаре, причем большинство видов встречается в тропических областях южного полушария (Окснер, 1956). Род Usnea, виды которого принимают заметное участие в составе эпифитных лишайниковых синузий хвойных лесов Голарктики, также тропического происхождения. Бореализированными тропогенными лишайниками являются некоторые представители рода Lecanora (Макаревич, 1963, 1964).
Таким образом, виды бореального элемента различны по своему происхождению. Некоторые из них, например виды рода Cornicularia, по-видимому, автохтонного бореального генезиса и относятся к числу древних бореальных типов. Однако более богато в бореальном элементе представлены бореализированные дериваты тургайской флоры, а также тропогенные формы. Распространение ряда систематических групп лишайников бореального элемента тяготеет к районам восточной и юго-восточной нетропической Азии. Это представляет большой интерес с историко-ботанической точки зрения, так как позволяет проследить, каким путем тропические типы, переходя в теплоумеренные условия, постепенно трансформировались в современные бореальные (Васильев, 1958).
IV. Неморальный элемент. Понятие неморального элемента было впервые предложено А. С. Лазаренко (1944), который относил к этой географической группе виды мхов, распространенные преимущественно в широколиственных лесах Горлактики. В статье, посвященной анализу неморального элемента в лихенофлоре советской Арктики, А. Н. Окснер (1944) указывал, что неморальный элемент охватывает виды, ареалы которых ограничены областью широколиственных лесов. М. Ф. Макаревич (1963, 1964), а вслед за ней Ш. О. Бархалов (1975) включали в состав неморального элемента виды, встречающиеся в зоне широколиственных лесов Голарктики. Однако X. X. Трасс (1970) указывал, что для анализа флор ограниченной территории важно объединить в неморальный элемент и те члены неморальной флоры, которые, кроме Голарктики, встречаются также и во флорах других флористических царств. Таким образом, X. X. Трасс, а за ним и А. В. Питеранс (1982) включили в неморальный элемент виды лишайников, распространенные в зоне широколиственных лесов Голарктики и вне Голарктики в аналогичных местообитаниях.
Высокое географическое положение Монголии над уровнем океана и крупные горные поднятия, расположенные на ее территории, являются причиной того, что во флоре Монголии из неморальных лишайников встречаются виды горно-неморального распространения. Таким образом, неморальный элемент в лихенофлоре Монголии объединяет 45 видов лишайников (5.9%), распространенных в равнинных широколиственных лесах и в лесах предгорий и нижнего пояса горных районов Голарктики и других флористических царств.
В состав неморального элемента входят представители 11 семейств. Наибольшее число неморальных видов насчитывают семейства Physciaceae (14) и Parmeliaceae (13). В семействе Candelariaceae неморальный элемент представлен 3 видами, в семействах Pertusariaceae Usneaceae, Lobariaceae, Lecideaceae и др. - 2 видами.
А. Голарктический неморальный субэлемент охватывает 25 видов лишайников, ареалы которых связаны с подзоной широколиственных лесов и нижним лесным поясом горных районов Голарктики. Им свойственны следующие типы ареалов.
1. Восточноазиатский тип ареала характерен для 4 видов - Phaeophyscia hirtuosa (Krempelh.) Golubk., Parmelia cochleata Zahlbr., P. bulbochaeta Hale, Cetraria rugosa (Asah.) Sato. Ареалы этих видов связаны с лесами Дальнего Востока, Японии, Восточного и Юго-Восточного Китая, реже с горными районами субтропических и тропических областей Юго-Восточной Азии. Вне этих районов, главным образом в Сибири и Северной Монголии, имеются лишь единичные разобщенные местонахождения этих видов, при этом западная граница их ареалов обычно проходит не западнее района Алтая (см. главу "Реликтовые элементы").
2. Евразиатский тип ареала свойствен 5 видам - Cetraria laureri Krempelh. (рис. 18), Pertusaria henrici (Harm.) Erichs., Arthopyrenia grisea (Schleich.) Koerb., Candelariella reflexa (Nyl.) Lett., Evernia arenaria (Retz.) Fr., Leptogium hildenbrandii (Garov.) Nyl.
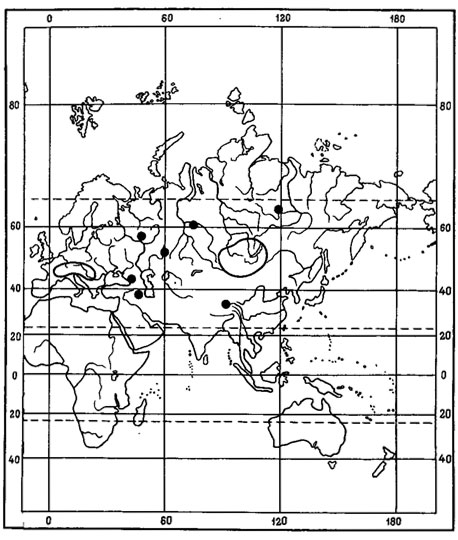
Рис. 18. Распространение на земном шаре Cetraria laureri Krempelh. (По: Макаревич, 1968)
Основной ареал Cetraria laureri Krempelh. (рис. 19,3) связан с Азией, охватывает о-в Сахалин, Даурию, горы Якутии, западное и восточное побережья оз. Байкал, Саяны, Монголию (Прихубсугулье, Хангай), Алтай. Этот вид известен также на Кавказе. В Европе имеются лишь разрозненные местонахождения этого лишайника на Урале, в Татарской АССР, в восточных Карпатах, Альпах. А. Н. Окснер (1946а) писал, что распространение этого вида связано с рефугиумами неморальной флоры Евразии. В Монголии этот лишайник встречается в районе восточного Прихубсугулья, где обитает в лиственничных лесах, иногда с примесью березы, на коре деревьев (Голубкова, 1981). В Хангае был собран лишь однажды, в верховьях реки Хух-Сум, на высоте 2100 м над ур. м., на основании ствола кедра в лиственнично-кедровом лесу (Бязров и др., 1983).
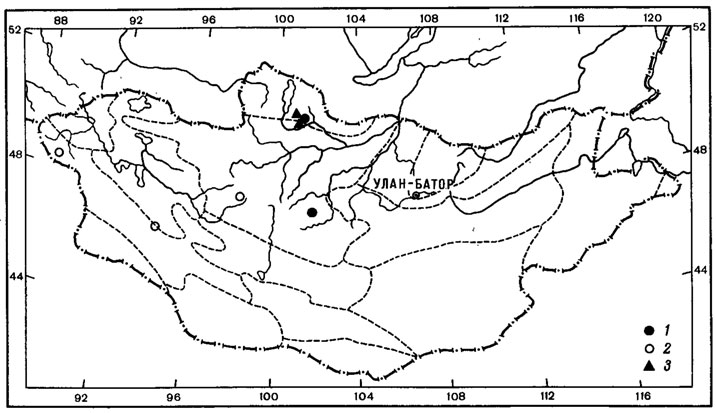
Рис. 19. Распространение на территории Монголии Cetraria komarovii Elenk. (1), C. potaninii Oxn. (2) и C. laureri Krempelh. (3). (Ориг.)
Ареалы других видов этой группы приурочены главным образом к листопадным лесам Европы, а в Азии имеются лишь отдельные разрозненные местонахождения этих лишайников. Pertusaria henrici (Harm.) Erichs. - эпифитный лишайник, обитающий главным образом на коре деревьев лиственных и нередко плодовых пород, рассеянно встречается по всей Европе, от южной Скандинавии до Италии, Испании, в Крыму, на Кавказе (Erichsen, 1936; Макаревич, 19716). В Азии этот вид найден только на Алтае (Седельникова, 1977а); он обнаружен также в Северо-Западной Монголии, в Сагил сомоне, на перевале Улан-Даба, в лиственничном лесу (Schubert, Klement, 1971), а кроме того, в центральном Хангае. Как отмечали Л. Г. Бязров и др. (1983), в Хангае этот лишайник является редким, обитающим на коре лиственницы в травяных лиственничниках нижнего лесного пояса.
Распространение Candelariella reflexa (Nyl.) Lett, сконцентрировано в Северной, Средней и Западной Европе, а также в Средиземноморской области, где этот лишайник обитает на старой коре на основаниях стволов деревьев, нередко плодовых, на гниющей древесине (Hakulinen, 1954; Poelt, 1969; Макаревич, 19716). На территории Азии этот вид найден в Монголии, в южном Хэнтэе, на коре тополя (Schubert е. а., 1969), а также в Монгольском Алтае, в лиственничном лесу на коре лиственницы (Голубкова, 1981).
Распространение Evernia arenaria (Retz.) Fr. недостаточно хорошо изучено, так как долгое время этот вид рассматривали как одну из форм Е. prunastri (L.) Ach.
Leptogium hildenbrandii (Qarov.) Nyl. (рис. 8, 2) ранее относили к мультирегиональным лишайникам, однако этот лишайник является евразиатским видом (J∅rgensen, 1973, 1975). В Европе основное распространение L. hildenbrandii (Garov.) Nyl. связано с южными районами; северная граница его ареала, видимо, проходит на Британских островах, но даже на севере Франции он очень редок (J∅rgensen, 1975). В этих районах этот лишайник обитает главным образом на коре деревьев широколиственных пород, на липах, вязах, ясенях и др. В Азии этот вид встречается в Китае, Японии (J∅rgensen, 1975), в Приморском крае, в Восточном Саяне, в Монголии, а также на Каказе, в Армении (Гербарий БИН АН СССР). В Монголии L. hildenbrandii (Garov.) Nyl. был найден только в Монгольско-Даурском районе, в долине р. Улдза. Здесь он произрастает во влажных местообитаниях на замшелых скалах в березовых лесах, среди нагромождений скал, поросших мхами (Голубкова, 1981).
Это вид восточноазиатского генезиса. В секции Mallotium он принадлежит к систематической группе видов, характеризующихся наличием на нижней стороне слоевища довольно длинных белых густых ризин. Центром развития этой группы "длинноволосатых" видов, насчитывающей 8 видов, является Юго-Восточная Азия (J∅rgensen, 1975), причем из них 4 вида относятся к числу эндемиков этого района земного шара. Изолированные местообитания L. hildenbrandii (Garov.) Nyl. на территории Монголии, так же как и в Восточном Саяне, видимо, реликтовые и свидетельствуют об обширном непрерывном ареале этого вида, вероятно, в третичное время, охватывающем Юго-Восточную Азию, Дальний Восток, Южную Сибирь, Кавказ и Южную и Западную Европу.
3. Голарктический тип ареала характерен для 14 видов лишайников - Parmelia exasperata DNot., P. glabratula (Lamy) Nyl., Arthopyrenia rhyponta (Ach.) Massal., Candelariella xanthostigma (Pers.) Lett., Phaeophyscia endococcina (Koerb.) Moberg, Physcia tenella (Scop.) DC., Ph. dimidiata (Arnold) Nyl., Physconia grisea (Lam.) Poelt, Rinodina pyrina (Ach.) Arnold, Catinaria laureri (Hepp) Degel., Biatora sylvana Koerb., Ochrolechia parella (L.) Massal., Cetrelia olivetorum (Nyl.) Culb. et Culb., C. cetrarioides (Del. ex Duby) Culb. et Culb.
Ареалы большинства видов неморального элемента голарктического типа не имеют непрерывного широтного простирания в Голарктике, они в основном фрагментарны. При этом для одних видов характерна приуроченность основного ареала к районам Северной Америки, Европы, в то время как в Азии имеются лишь отдельные изолированные местонахождения, причем наиболее восточные из них находятся на территории Монголии. Основная часть ареала других видов, наоборот, приурочена к Азии, и на территории МНР проходит наиболее западная граница их распространения в Азии.
В качестве примера рассмотрим распространение представителей родов Parmelia - P. exasperata DNot. и P. glabratuia (Lamy) Nyl., и Cetrelia - С. olivetorum (Nyl.) Culb. et Culb., C. cetrarioides (Del. ex Duby) Culb. et Culb.
Основной ареал Parmelia exasperata DNot. связан с восточными районами Северной Америки и Европой, где этот вид обитает почти исключительно на стволах и ветвях деревьев листопадных пород, изредка переходя на хвойные породы и на скалы (Esslinger, 1977). На территории Северной Америки его ареал простирается от Новой Англии и Великих озер и далее на юг до северных районов штата Джорджия в Аппалачах и до северной части штата Арканзас в долине р. Миссисипи. Этот вид известен в Исландии. В Европе этот эпифитный лишайник широко распространен от Северной Скандинавии до средиземноморских районов Европы и далее к югу до южных районов Северной Африки, от Великобритании и Португалии на западе до территории европейской части СССР на востоке (Esslinger, 1977). В европейской части Советского Союза ареал P. exasperata DNot. охватывает Прибалтику, Белоруссию, Новгородскую, Архангельскую, Московскую, Ярославскую, Владимировскую области, Татарию, Саратовскую область, на юге достигая южных районов Украины, Крыма, Кавказа (Рассадина, 1971); отдельные местообитания этого вида известны и в более северных районах - в Архангельской и Мурманской областях, где, однако, этот вид редок (Домбровская, 1970а). Особенно характерен этот лишайник для южной части лесной зоны, а также для лесостепной (Рассадина, 1971). Далее на восток известны лишь отдельные изолированные местонахождения этого вида на Урале - Среднем и Южном (Рябкова, 1982), в Западной Сибири, Казахстане, Монголии. Так, в Западной Сибири он приводится для Горной Шории (Седельникова, 1977б), в Казахстане - для Заалийского Алатау (Окснер, 19396), где обитает в лесном поясе, на высоте 1300 - 1700 м, на коре елей. На территории МНР, в горных районах - в западном и центральном Хангае и южном Хэнтэе (Kjement, 1965; Schubert е. а., 1969; Schubert, Klement, 1971; Бязров и др., 1983), видимо, находятся восточные местообитания этого неморального лишайника на Евразиатском континенте. В Монголии этот вид является редким. Так, Л. Г. Бязров и др. (1983) отмечали, что в районе центрального Хангая этот лишайник изредка встречается на стволах деревьев в нижнем лесном поясе в травяных лиственничниках.
Ареал Parmelia glabratuia (Lamy) Nyl. также связан с областью распространения широколиственных лесов Европы и Северной Америки: в Азии известны лишь отдельные местонахождения его (Esslinger, 1977). В отличие от P. exasperata DNot. P. glabratula (Lamy) Nyl. на территории Северной Америки встречается не только в восточных, но и в западных районах. На западе этого континента он распространен от южного берега Аляски до Южной Калифорнии, далее на восток до района Великих озер, Новой Англии, достигая на юге Северной Каролины. Этот лишайник известен также в Исландии. В Европе ареал P. glabratula (Lamy) Nyl. охватывает территорию от Южной Скандинавии до Южной Европы, достигая на юге Северной Африки (Марокко), и от Британских островов на западе до территории Советского Союза на востоке (Esslinger, 1977). В европейской части СССР P. glabratula (Lamy) Nyl. встречается в Карелии, Прибалтике, Белоруссии, Ленинградской области, на Украине (Карпаты, Крым), в Молдавии, на Кавказе, проникая на восток до Татарии, Кировской области, Южного Урала; известны также его находки на юге Мурманской области (Домбров ская, 1970а; Рассадина, 1971; Esslinger, 1977; Рябкова, 1982; Гербарий БИН АН СССР). В Азии имеются изолированные местообитания этого вида в Западной Сибири, в долине р. Конды (Vainio, 1928), в Восточной Сибири - в Байкальском хребте (Гербарий БИН АН СССР), а также в Монголии - в южном Хэнтэе (гора Булган-Ула) и в долине р. Тэрэлджийн-Гол (Schubert е. а., 1969).
Представители рода Cetrelia в своем распространении, наоборот, проявляют явное тяготение к районам Восточной и Юго-Восточной Азии. Юго-восточная часть Азии является центром видовой насыщенности этой систематической группы лишайников; из 14 видов этого рода 13 встречаются здесь, причем 10 видов эндемичны для флоры этих районов (Culberson, Culberson, 1968). Особенно богата видами рода Cetrelia флора Юго-Восточного Китая, где известно 10 видов этих лишайников. В Европе встречаются 2 вида, в Северной Америке - 4, из них один вид - эндемик североамериканской флоры. Во флоре Монголии род Cetrelia представлен двумя видами - С. olivetorum (Nyl.) Culb. et Culb. и С. cetrarioides(Del. ex Duby) Culb. et Culb.
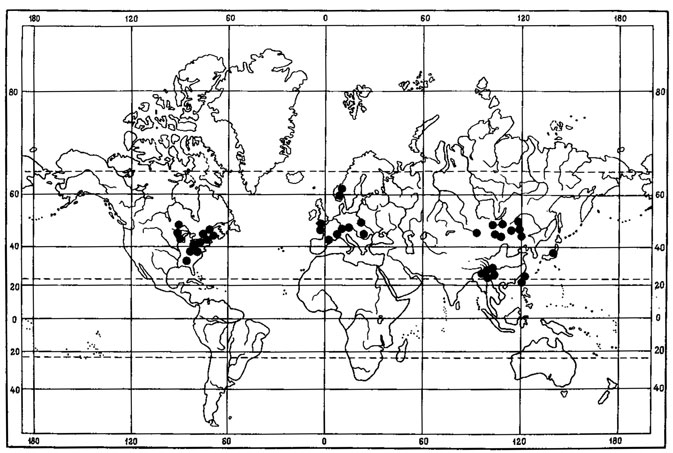
Рис. 20. Распространение на земном шаре Cetrelia olivetorum (Nyl.) W. Culb. et C. Culb. (По: Culberson, Culberson, 1968, с дополнениями автора)
Ареал Cetrelia olivetorum (Nyl.) Culb. et Culb. (рис. 20) состоит фактически из нескольких элементарных ареалов, связанных с рефугиальными центрами неморальной флоры и разобщенными крупными дизъюнкциями, что характерно, как отмечал А. Н. Окснер (1946а), для форм тургайского геоэлемента. Основная часть ареала этого вида связана с юго-восточными районами Азии - Японией, Китаем (провинции Юньнань, Сычуань, о-в Тайвань), Непалом. После некоторой дизъюнкции этот вид вдстречается на территории Монголии - в Восточной Монголии, в долинах р. Керулен и Улдза, в Хангае, Прихубсугулье, где обитает на замшелых силикатных и базальтовых скалах, иногда на растительных остатках. Другая часть ареала С. olivetorum (Nyl.) Culb. et Culb. связана с северо-восточными районами Северной Америки - штатами Мэн, Нью-Гэмпшир, Вермонт, Нью-Йорк, Пенсильвания, Виргиния, Северная Каролина, Огайо, Мичиган, Висконсин, т. е. в основном приурочена к горам Аппалачи (Culberson, Culberson, 1968). Кроме того, этот лишайник приводится также для Аляски (Krog, 1968). Еще одна часть ареала этого вида приурочена к Средней и приатлантической Европе: он встречается в горных районах Румынии, Польши, Венгрии, Швейцарии, ФРГ, Франции (Culberson, Culberson, 1968). На севере ареал этого вида захватывает южные районы Скандинавии, Южной Норвегии (Lynge, 1921; J∅rgensen, Ryvarden, 1970; J∅rgensen, ∅vstedal, 1975), Швеции и Финляндии (Du Rietz, 1924; Rasanen, 1939; Hakulinen, 1963, и др.) Однако, некоторые эти сообщения, возможно, относятся к С. cetrarioides (Duby) Culb. et Culb.
Б. Омнинеморальный субэлемент объединяет виды лишайников, ареалы которых связаны с листопадными лесами Голарктики и других флористических царств.
1. Mультирегиональный тип ареала свойствен 21 виду неморальных лишайников - Parmelia soredica Nyl., P. rudecta Ach., P. subrudecta Nyl., P. caperata (L.) Ach., Candelaria concolor (Dicks.) Stein, Physcia aipolia (Ehrh.) Fürnr., Ph. stellaris (L.) Nyl., Physconia pulverulacea Moberg, Anaptychia ciliaris (L.) Koerb., Hyperphyscia adglutinata (Flk.) Mayrh. et Poelt, Leptogium cyanescens (Hoffm.) Koerb., L. burnetiae Dodge, Lobaria retigera (Bory) Trevis., L. isidiosa (Müll. Arg.) Vain., Pseudevernia cirrhata (Fr.) Schub. et Klem., Ramalina farinacea (L.) Ach., Phaeophyscia hispidula (Ach.) Moberg, Heterodermia speciosa (Wulf.) Trevis. (рис. 21), H. leucomela (L.) Poelt, Xanthoma parietina (L.) Th. Fr., X. fallax (Hepp) Arnold.
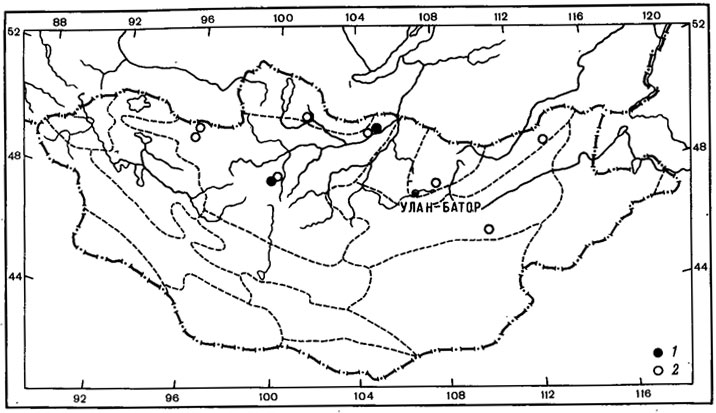
Рис. 21. Распространение на территории Монголии Heterodermia speciosa (Wulf.) Trevis. (1) и Phaeophyscia hispidula (Ach.) Moberg (2). (Ориг.)
Для этих видов характерны обширные ареалы, охватывающие северное и южное полушария, прерываемые, как правило, огромными дизъюнкциями. В Голарктике их распространение связано в основном с широколиственными и смешанными лесами, но некоторые из них встречаются и в таежной зоне и заходят даже в Арктику. Как отмечал А. Н. Окснер (1946а), обладая в самых широких границах способностью к адаптации, лишайники гораздо свободнее, чем цветковые растения, выживают в неблагоприятных условиях далеко от основной части своего ареала и иррадиируют за пределы области развития географического элемента, к которому они относятся. Среди видов неморального элемента мультирегионального типа выделяется довольно большая тропогенная группа видов (более 10), основной ареал которых связан с тропическими и субтропическими районами земного шара.
В качестве примера приведем ареал Leptogium burnetiae Dodge (рис. 22). Основная часть ареала L. burnetiae находится в Азии, где он распространен в тропической Индии, а также в Гималаях, на о-ве Шри Ланка, в Пакистане, Китае - в провинциях Юньнань, Хунань и Шэньси, в Японии и далее севернее на Дальнем Востоке в Хабаровском крае, в Монголии (южный Хэнтэй, Хангай) и на Алтае. В Европе L. burnetiae Dodge обитает в горных лесах Средней и Южной Европы (Франция, Швейцария, Австрия, Италия). Одна часть ареала этого вида (J∅rgensen, 1973) связана с тропическими районами Восточной Африки (Эфиопией, Кенией, Танзанией), где он встречается на коре деревьев главным образом в горных лесах; на западе Африканского континента известен в Анголе, на юге достигает Капской провинции (ЮАР). Еще одна часть ареала вида приурочена к юго-восточным районам Северной Америки, от Атлантического побережья Северной Америки до штатов Колорадо и Южная Дакота, на севере он известен в Аляске. Кроме того, этот вид встречается на Гавайских островах и в Южной Америке - в Венесуэле.
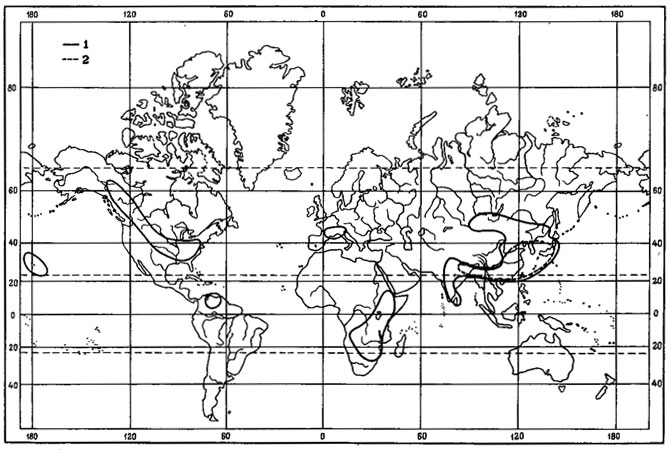
Рис. 22. Распространение на земном шаре Leptogium burnetiae Dodge (1) и L. рedicellatum J∅ rg. (2). (По: J∅ frgensen, 1975)
На территории Монголии этот вид является редким, он был найден только в трех местообитаниях - в южном Хэнтэе, в окрестностях Улан-Батора, и в центральном Хангае, западнее г. Цэцэрлэг, на горе Булган-Ула и в долине р. Худжирт-Гол, где он обитал на замшелых скалах в лиственничных лесах (Schubert, Klement, 1971; J∅rgensen, 1973; Голубкова, 1981).
Leptogium burnetiae Dodge генетически связан с восточноазиатским флористическим центром. Этот вид, как и евразиатский неморальный лишайник L. hildenbrandii (Garov.) Nyl., относится к секции Mallotium, к группе видов, для которых характерно образование на нижней поверхности слоевища густых беловатых ризин. Как отмечал Иоргенсен (J∅rgensen, 1975), на земном шаре имеются два центра, концентрирующих виды этой группы лишайников. Один из них находится в Южной Америке, где встречается 13 видов из этой группы лишайников. Причем здесь распространены главным образом "коротковолосатые" формы, длина ризин которых не превышает 20 мкм. Второй центр видовой насыщенности этой группы лишайников находится в Юго-Восточной Азии - 8 видов из этой группы лептогиумов. Эти, виды характеризуются образованием более длинных ризин, до 100 мкм длиной ("длинноволосатые" формы). Центральное место среди них занимает группа L. tichophorum Miill. Agr., объединяющая азиатские виды лептогиумов, при этом фертильные формы. В составе группы 4 азиатских эндемика - L. tichophorum Miill. Arg. (Индия, Япония, Филиппины), L. delavayi Hue (Индия, Япония), L. arisanense Asah. (о-в Тайвань), L. pedicellatum J∅rg. (Индия, Япония). Остальные представители восточноазиатской группы лишайников являются стерильными "парами" этих фертильных форм, для них характерно образование изидий. Эти стерильные формы часто имеют более широкое распространение по сравнению с их фертильными "парами" (J∅rgensen, 1975), тем самым подтверждая ту же закономерность в распространении "видовых пар", установленную еще Пельтом (Poelt, 1970). Все изидиозные формы лептогиумов азиатской группы выходят за пределы Азии и встречаются в других районах земного шара. Например, L. asiaticum Jtfrg. - изидиозная пара L. tichophorum Miill. Arg., помимо Юго-Восточной Азии, встречается в Восточной Африке. L. burnetiae Dodge филогенетически наиболее близок к индо-японскому виду L. pedicellatum J∅rg.
Эта изидиозная форма, характеризующаяся, как было показано выше, обширным мультирегиональным ареалом, является стерильной парой L. pedicellatum J∅rg. (рис. 22, 2).
Большинство видов неморального элемента является редкими на территории МНР и было собрано лишь в отдельных местонахождениях. В числе наиболее широко распространенных в Монголии неморальных лишайников можно назвать Xanthoria fallax (Нерр) Arnold, Parmelia soredica Nyl. При этом следует подчеркнуть, что местонахождения неморальных лишайников, как правило, концентрируются в определенных районах МНР. Особенно богат ими район Прихубсугулья (восточное Прихубсу- гулье), где были найдены такие неморальные виды лишайников, как Lobaria retigera (Bory) Trevis., L. isidiosa (Miill. Arg.) Vain., Cetraria laureri Kremplh., Parmelia soredica Nyl., Hyperphyscia adglutinata (Flk.) Mayrh. et Poelt, Phaeophyscia hispidula (Ach.) Moberg и др. Они были собраны главным образом в окрестностях оз. Хубсугул, в бассейне р. Эгин-Гол, в долине р. Аригийн-Гол. Довольно богата видами неморального элемента флора южного Хэнтэя (долина р. Тэрэлджийн-Гол, окрестности г. Улан-Батора, гора Богдо-Ула). В этом районе встречаются Lobaria retigera (Bory) Trevis., L. isidiosa (Mull. Arg.) Vain., Parmelia caperata (L.) Ach., Phaeophyscia hirtuosa (Krempelh.) Golubk., Ph. endococcina (Koerb.) Moberg и др. Хангай беднее неморальными видами лишайников по сравнению с первыми двумя районами, при этом они были найдены здесь в различных частях района. В северозападном Хангае, на правобережье р. Эгин-Гол, была собрана Phaeophyscia hirtuosa (Krempelh.) Golubk.; в долинах рек Селенги и Орхон - Cetrelia olivetorum (Nyl.) Culb. et Culb.; Heterodermia speciosa (Wulf.) Trevis. был отмечен также в окрестностях оз. Цаган-Нур. Наиболее богатым видами лишайников неморального элемента оказался район в окрестностях г. Цэцэрлэг - гора Булган-Ула, где были найдены Parmelia bulbochaeta Hale, P. cochleata Zahlbr., Cetraria rugosa (Asah.) Sato, Phaeophyscia endococcina (Koerb.) Moberg, Leptogium burnetiae Dodge и др. Некоторые неморальные лишайники - Leptogium hildenbrandii (Garov.) Nyl., L. cyanescens (Hoffm.) Koerb., Phaeophyscia hispidula (Ach.) Moberg, Parmelia soredica Nyl. и др. были собраны в долинах рек Улдза, Керулен и Онон, особенно богата ими долина р. Улдза. В районе Прихинганья были встречены такие неморальные виды, как Hyperphyscia adglutinata (Flk.) Mayrh. et Poelt, Xanthoria parietina (L.) Th. Fr., Phaeophyscia hirtuosa (Krempelh.) Golubk., Parmelia soredica Nyl. Слабо представлен неморальный элемент во флоре лишайников Монгольского Алтая. Изредка неморальные лишайники - Cetrelia olivetorum (Nyl.) Culb. et Culb., Parmelia caperata (L.) Ach., Hyperphyscia adglutinata (Flk.) Mayrh. et Poelt - встречаются в Средней Халхе и Восточной Монголии.
Следует отметить также, что на территории МНР очень немногие виды этого элемента обитают на коре деревьев лиственных пород. Среди них можно назвать Xanthoria parietina (L.) Th. Fr. и Phaeophyscia hirtuosa (Krempelh.) Golubk., найденные в району Прихинганья, в пойме р. Нумэргин-Гол, на коре вяза. Некоторые неморальные лишайники, например Parmelia caperata (L.) Ach., встречаются на коре лиственниц. Большинство же представителей неморальных лишайников - Lobaria retigera (Bory) Trevis, Leptogium cyanescens (Hoffm.) Koerb., Heterodermia speciosa (Wulf.) Trevis. и др. - в условиях Монголии обитает на замшелых скалах, изредка переходя на замшелые гниющие стволы деревьев, замшелую почву, растительные остатки. Как отмечала Н. В. Седельникова (1977а), в лихенологической литературе имеется немало данных относительно изменения субстрата некоторыми лишайниками в периферийных частях ареала с депрессивными для них условиями существования. А. Н. Окснер (1946а) по этому поводу писал, что переход с древесного субстрата на затененные скалы, покрытые лишайниками и мхами, ставит теплолюбивые мезофильные лишайники в условия, лучшие не только по влажности (Suza, 1933), но и по температурному режиму. Характерно также, что неморальные виды лишайников в условиях Монголии предпочитают для своего обитания микроэкотопы с повышенной влажностью воздуха. Большинство из них поселяется по берегам рек, озер, под покровом леса среди нагромождений замшелых скал и т. д.
А. Н. Окснер (1946а) указывал, что по происхождению неморальный элемент в основном представляет собой дериват тургайской флоры. Но, рассматривая вопрос о генетических корнях неморальной флоры Голарктики, А. Н. Окснер подчеркивал, что в неморальной флоре Голарктики можно обнаружить лишь незначительное число автохтонных тургайских родов, причем большинство из них не сохранило свой первоначальный облик, а были позднее охвачены процессом бореализации. Главное место в неморальной флоре Голарктики занимают тропогенные формы. В комплексе тропогенных форм А. Н. Окснер различает ряд генетических элементов: реликты тургайской флоры, реликты полтавской флоры, неморальные, сравнительно молодые, виды, возникшие в условиях умеренной области в конце третичного периода или в квартере в результате переработки форм из тропогенных родов и тропические формы, пришельцы из тропических стран.
Виды неморального элемента, обнаруженные в Монголии, по своему происхождению не представляют однородного комплекса. К числу автохтонных тургайских родов, видимо, можно отнести род Cetrelia. Основное распространение рода связано с Восточной и Юго-Восточной Азией: из 14 видов этого рода 13 встречаются в этих районах земного шара, причем 10 видов эндемичны для них (Culberson, Culberson, 1968). Особенно богата видами этого рода флора Юго-Восточного Китая, где известно 10 видов этих лишайников.
К группе автохтонных тургайских родов А. Н. Окснер (1946б) относил род Evernia, который, однако, как он отмечает, в дальнейшем подвергся сильной бореализации. Е. Ф. Флоровская (1952), монограф этой группы лишайников, считала, что возникновение исходных форм рода Evernia можно отнести к верхнему мелу. Причем ею высказано предположение, что в третичный период этот род был представлен несколькими видами, из которых наиболее древними являются Е. divaricata (L.) Ach., Е. esorediosa (Mull. Arg.) DR., E. prunastri (L.) Ach.
В настоящее время род Evernia насчитывает 13 видов, причем почти все они встречаются в Голарктике, не выходя за пределы умеренной зоны, лишь один вид проник в южное полушарие, в Южную Америку (Чили.). Из 12 видов еверний 8 встречается в Азии, из них 6 - в Восточной Азии.
К автохтонным тургайским родам, по всей вероятности, можно причислить и род Cetraria, который, так же как и род Evernia, позднее подвергся сильному влиянию процесса бореализации. Как писала К. А. Рассадина (1950, с. 184), монограф этой группы лишайников, "исходный центр происхождения Cetraria надо искать в Азии, в горах Южной Сибири, Гималаях, Китае, Индии". Особенно богата флора данных районов эпифитными формами этого рода, представителями секции Platysma, с эволюционно наиболее примитивно устроенными листоватыми слоевищами, к числу которых относятся, например, С. laureri Krempelh.
А. Н. Окснер (1946а) полагает, что в неморальной флоре Голарктики очень скромная роль отведена древнесредиземно- морским формам. К числу таких форм он причисляет Anaptychia ciliaris (L.) Koerb., представленную и в лихенофлоре Монголии. Особым богатством анаптихий отличается гирканский флористический район (и, видимо, Северный Иран). A. ciliaris (L.) Koerb. - древний средиземноморский вид, как полагает А. Н. Окснер, развился не позднее середины третичного периода в Средиземноморской области, а в дальнейшем широко расселился в Северной Америке и мигрировал по горным цепям Передней и Средней Азии к хребтам Юго-Западного Китая, а также в Монголию.
Среди видов неморального элемента во флоре лишайников Монголии, как и в неморальной флоре Голарктики, одно из основных мест занимают тропогенные формы, которые представлены видами родов Heterodermia - Н. leucomela (L.) Poelt, Н. speciosa (Wulf.) Trevis., Lobaria - L. retigera (Bory) Trevis., L. isidiosa (Mull. Arg.) Vain., Pseudevernia cirrhata (Fr.) Schub. et Klem., и др. Большинство из них является неизменными тропическими формами.
Слабее в неморальном элементе лихенофлоры Монголии представлены формы, возникшие в составе тургайской флоры в результате трансформации тропических форм. Примером могут служить виды рода Arthopyrenia, у которых прослеживаются тропогенные связи.
В качестве примера рассмотрим род Heterodermia. Тропический генезис рода вполне очевиден. Род Heterodermia насчитывает более 70 видов, причем наибольшее число этих видов (около 75%) распространено в тропических и субтропических зонах земного шара (Kurokawa, 1962; Poelt, 1965; Culberson, 1966; Swinscow, Krog, 1976). Многим видам рода свойственны обширные пантропические ареалы, охватывающие тропические районы Юго-Восточной Азии, Африки, Южной и Центральной Америки. Особенно богаты видами этого рода флоры Юго-Восточной Азии и Центральной Америки. Всего 10 видов этой группы лишайников проникает в умеренные районы Голарктики, преимущественно в восточные районы Северной Америки и в Восточную Азию (Япония, Дальний Восток, Курилы, Камчатка). В Европе известно 4 вида этого рода, причем основное их распространение связано с Южной и приатлантической Европой. Во флоре Монголии род Heterodermia представлен 2 видами - Н. speciosa (Wulf.) Trevis., Н. leucomela (L.) Poelt. Ареал H. speciosa (Wulf.) Trevis. (Kurokawa, 1962; Culberson, 1966) очень обширен, с огромными разъединениями, свидетельствующими о древнем происхождении этого вида (рис. 21). Основная часть его азиатского ареала связана с Восточной и
Юго-Восточной Азией, где он встречается в Индии, Китае, Японии, на о-ве Сахалин, в Приморском и Хабаровском краях. Далее на запад имеются изолированные местонахождения этого вида в Монголии (Хангай) (Голубкова, 1981), в Забайкалье (р. Ингода) (Окснер, 1946б), в Кузнецком нагорье, в долинах рек Конды и Иртыша, на Урале (Vainio, 1928; Седельникова, 1977а, 1977б). Изолированное местообитание этого вида известно также на побережье Берингова моря, в устье Анадыря (Рассадина 1934). В Европе он рассеянно встречается в лесах горных районов Средней Европы, в Альпах и Средиземноморских районах, реже в океанических районах Западной Европы, обитая на старых замшелых стволах деревьев, реже на скалах (Poelt, 1969), на севере достигая Финляндии (Hakulinen, 1962), Центральной Норвегии (Flatberg е. а., 1975), Этот лишайник известен также в Крыму и на Кавказе (Пахунова 1956; Бархалов, 1957, 1975). На территории Северной Америки его ареал приурочен к горным районам восточной части, но этот лишайник отсутствует на побережье (Culberson, 1966); имеются также изолированные местонахождения его на тихоокеанском побережье Аляски (Krog, 1968). Еще одна часть ареала Н. speciosa (Wulf.) Trevis. приурочена к тропическим и субтропическим районам Центральной и Южной Америки, Гавайским островам (Kurokawa, 1962; Culberson, 1966). Он известен также на островах Тристан-да-Кунья (J∅rgensen, 1977). Кроме того, этот лишайник обычен в горных районах Восточной Африки (Swinscow, Krog, 1976), на стволах деревьев, реже на скалах, в пределах высот 1100 - 3600 м над ур. м. В Монголии этот вид был собран в Хангае, в долине р. Селенги и окрестностях оз. Цаган-Нур, где обитал в горных степях на замшелых скалах (Голубкова, 1981).
Второй вид этого рода - Н. leucomela (L.) Poelt - также широко распространен на земном шаре. Его ареал охватывает пантропики и неотропики (Индия, острова Тайвань, Ява, Филиппинские, Гавайские, Африка, Южная и Центральная Америка). За пределами тропических и субтропических зон этот вид встречается в горных районах восточной части Северной Америки, в Южной и Средней Европе (Швейцария, Франция, Испания), в умеренной Азии (Япония, Китай, МНР) (Kurokawa, 1962; Poelt, 1965; Culberson, 1966; Бязров, 1976). Местонахождение этого вида на территории Монголии рассматривается нами как реликтовое.
Находки видов неморальной флоры в Монголии, где отсутствуют формации широколиственных лесов, несомненно представляют интерес. Ареалогический и флорогенетический анализ видов неморального элемента, обнаруженных во флоре лишайников Монголии, приводит к выводу, что ряд видов этого элемента является, по-видимому, во флоре Монголии реликтами. Причем одни из них, возможно, третичные реликты тургайской флоры, другие, вероятно, мигрировали на территорию Монголии в одну из влажных фаз голоцена.
Иммиграция мезофильных неморальных элементов на территорию Монголии, видимо, происходила разными путями. Причем особенно велика роль в формировании мезофильной неморальной флоры лишайников этой страны восточноазиатского флористического центра, о чем свидетельствуют как генетические связи неморальных видов лишайников, так и их современные ареалы. Как было показано выше, к числу видов, иммигрировавших на территорию Монголии с востока, относятся виды рода Cetrelia, Cetraria laureri Krempelh., Phaeophyscia hirtuosa (Krempelh.) Golubk., Parmelia subrudecta Nyl., P. rudecta Ach. и др., а также целый ряд тропических форм - Lobaria isidiosa (Miill. Arg.) Vain., L. retigera (Bory) Trevis., L. burnetiae Dodge и др. Значительно меньшая роль принадлежит западноевропейскому центру (Parmelia glabratuia (Lamy) Nyl., Pertusaria henrici (Harm) Erichs.). Иммиграция некоторых неморальных видов, возможно, происходила из сибирских рефугиумов неморальной флоры. Не исключена также их миграция из горных районов Средней Азии, где термофильная неморальная флора могла пережить эпоху оледенения в нижнем поясе среднеазиатских гор. Об этом, например, свидетельствуют находки в Заалийском Алатау неморального лишайника Parmelia exasperata DNot. (Окснер, 1939б). Как отмечал Е. П. Коровин (1961), конец ледникового и начало послеледникового времени было в Средней Азии самым благоприятным для развития лесной мезофильной флоры.
Л. В. Бардунов (1973а), характеризуя флорогенетические особенности моховых флор, писал, что для бриофлоры любого региона свойственно наличие довольно больших групп видов мхов, способных выдержать даже достаточно серьезные физико-географические перестройки и войти в состав новой флоры без изменения своей видовой специфики и без перехода в реликтовое состояние в качестве полноправных ее представителей. Это справедливо и в отношении лишайников. Так, среди неморального элемента лихенофлоры Монголии, помимо реликтовой группы видов, можно выделить селективную группу видов лишайников, вошедших в состав флоры Монголии и таежных районов Сибири как полноправные ее компоненты и широко в ней расселившихся. В числе лишайников этой группы - Physcia stellaris (L.) Nyl., Ph. aipolia (Ehrh.) Fiirnr., Ph. tenella (Scop.) DC., Xanthoria fallax (Hepp) Arnold, Parmelia soredica Nyl. и др.
V. Аридный элемент. Объединяет 166 видов (22% от общего числа видов), ареалы которых связаны с аридными областями Голарктики (голарктический аридный субэлемент), а также с другими флористическими царствами (омниаридный субэлемент).
В состав аридного элемента входят представители 14 семейств. Наиболее богато аридный элемент представлен в следующих семействах: Acarosporaceae (32 вида), Verrucariaceae и Aspiciliaceae (по 20 каждое), Lecanoraceae (16), Lichinaceae (15), Lecideaceae (12), Teloschistaceae (10), Heppiaceae и Physciaceae (no 7 каждое), Parmeliaceae и Thelotremataceae (no 6 видов каждое).
А. Голарктический аридный субэлемент включает 133 вида. Им свойственны следующие типы ареалов.
1. Монгольский тип ареала характерен для 21 вида, распространенного в пределах Монгольской провинции Центральной Азии, в понимании В. И. Грубова (1963). В составе этой группы эндемики и субэндемики флоры лишайников МНР. - Toninia gobica Golubk., Rhizoplaca baranowii (Poelt) Golubk., Peltula zabolotnoji (Elenk.) Golubk., Endopyrenium mongolicum (H. Magn.) Golubk., E. minutum (H. Magn.) Golubk., E. perminu- tum (H. Magn) Golubk., Endocarpon mongolicum H. Magn., Psorotichia dispersa H. Magn., Peccania polyspora H. Magn., Peltula minuta (H. Magn.) Golubk., Collema minutum H. Magn., Acarospora sarcogynoides H. Magn., A. suprasedens H. Magn., A. fulva Golubk. (табл. V, 2), A. umbrina H. Magn., A. verruculosa H. Magn. (табл. III, 2), Buellia mongolica H. Magn., Toninia olivaceoatra H. Magn., Sarcogyne picea H. Magn., Lecanora subminuta H. Magn., Squamarina kansuensis (H. Magn.) Poelt.
На территории МНР распространение этих видов связано с пустынно-степными районами гобийской части страны и Восточной Монголии; за пределами МНР эти лишайники встречаются во Внутренней Монголии и в Хэси (Китай), некоторые из них иррадиируют в горные степи северной части МНР.
Toninia gobica Golubk. и Rhizoplaca baranowii (Tomin) Poelt (табл. VI, 1, 2) характеризуются узколокальными ареалами. Ареал Toninia gobica Golubk. приурочен к восточной части Гобийского Алтая (Голубкова, 1973а; Голубкова, Цогт, 1974а; Голубкова, 1981), к хребтам Гурван-Сайхан, Хурху, Ноён, где этот вид обитает по склонам гор на гумусированных участках и в щелях скал (рис. 39, 2).
Ареал Rhizoplaca baranowii (Tomin) Poelt - напочвенного кочующего лишайника - связан с северо-западными районами Монголии. Он обитает в холоднополынно-типчаковых горных степях в хребте Сайлюгем и в северо-западной части Монгольского Алтая, проникая в Туву в нагорье Сангилен (Poelt, 1958; Голубкова, 1981; Цогт, 1981; Седельникова, 1983).
У многих других видов лишайников монгольского распространения также проявляется определенная приуроченность к тем или иным районам центральноазиатской части Монголии. С гобийскими районами МНР связано, например, распространение слизистого лишайника Peccania polyspora Н. Magn., растущего на гранитных и базальтовых скалах в гобийских пустынях, низкогорных хребтах, изредка в пустынных степях. Ареал этого лишайника простирается от южных районов Долины Озер, к восточным хребтам Гобийского Алтая (хр. Хурху), пустыням Галбын-Гоби и Алашаньская Гоби (Голубкова, 1981). За пределами МНР этот вид встречается в Хэси (Magnusson, 1940).
Acarospora verruculosa H. Magn., обитающая на скалах в горных и пустынных степях, характеризуется ареалом, охватывающим юго-западные и южные районы Монголии - Джунгарскую Гоби, хр. Адж-Богдо, юго-восточные хребты Гобийского Алтая (хребты Хурху, Тост-Ула, Ноен), на севере достигающим юго-восточной части Долины Озер, за пределами МНР - южных районов Внутренней Монголии (Китай) (Голубкова, 1981; Magnusson, 1944) (табл. III, 2).
Ареал ряда видов, например Acarospora sarcogynoides Н. Magn., A. suprasedens Н. Magn., Toninia olivaceoatra H. Magn., приурочен к юго-восточным районам МНР. Эти эпилитные лишайники являются характерными представителями флоры лишайников горных степей Восточной Монголии, Прихинганья; западная граница их ареалов проходит приблизительно в восточной части Средней Халхи; изредка они проникают в ее центральную часть и встречаются в юго-восточных хребтах Гобийского Алтая (Голубкова, 1981). За пределами МНР эти виды распространены на территории Внутренней Монголии (Magnusson, 1944), в Иньшане.
Ареал Acarospora fulva Н. Magn. (рис. 23, 2) связан главным образом с северными окраинами Центральной Азии (Голубкова, 1979, 1980б, 1981). Этот лишайник, обитающий на гранитных валунах и скалах, нередко на слоевищах других лишайников, обычен в сухих и горных степях Средней Халхи, в долине р. Керулен, проникает также в северо-западные районы Монголии и юго-восточные районы Хангая, проявляет инвазии в Прибайкалье.
Рис. 23. Распространение на территории Монголии Acarospora gobiensis H. Magn. (1), A. fulva Golubk. (2) и A. stapfiana (Mull. Arg.) Hue (3). (Ориг.)
Ряд видов этой группы лишайников широко распространен на территории Монголии. К числу их относится Peltula zabolotnoji (Elenk.) Golubk. Этот лишайник, растущий на почве, растительных остатках, гумусированных участках и в расщелинах скал, часто встречается в равнинных и горных степях, а также в пустынях Монголии. Особенно широко распространен он в юго-западной части Монголии - в Монгольском Алтае, Котловине Больших Озер, в Долине Озер, в Джунгарской, Заалтайской и Восточной Гоби, проникает и в Восточную Монголию (Голубкова, 1981; Цогт, 1981). Этот вид иррадиирует в северные районы Хангая, в долину р. Керулен.
В Гобийском Алтае, в Средней Халхе и Котловине Больших Озер часто встречается эпилитный лишайник Buellia mongolica Н. Magn., южная часть ареала которого проходит на территории Иньшаня (Китай) (Magnusson, 1944).
2. Тибетско-монгольский тип ареала свойствен видам, распространенным в пределах Монгольской и Тибетской провинций Центральной Азии (в понимании В. И. Грубова (1963)). В группу входят 12 видов. - Endopyrenium atrocinereum (Н. Magn.) Golubk., Е. kansuense (Н. Magn.) Golubk., Psorotichia mongolica H. Magn., P. kansuensis H. Magn., P. nigra H. Magn., Lecidea hedinii H. Magn., Buellia sarcogynoides H. Magn., B. hedinii H. Magn., Lecanora invadens H. Magn., L. semipallida H. Magn., L. isabellina H. Magn., Aspicilia determinata (H. Magn.) Golubk. Это эпилитные лишайники, иногда обитающие на гумусированных участках скал и в их расщелинах в пустынно-степных ценозах Монголии.
Довольно широко распространены в пустынно-степных районах Монголии слизистые лишайники рода Psorotichia, образующие на скалах, почвенных наносах и растительных остатках в щелях скал тонкие, черные налеты. Из видов этого рода наиболее часто встречаются P. kansuensis Н. Magn., P. nigra Н. Magn. Ареал P. kansuensis Н. Magn. охватывает Монгольский Алтай, Долину Озер, пустыни Заалтайскую и Восточную Гоби (Голубкова, 1981; Цогт, 1981), а за пределами МНР на территории Ганьсу, в северных предгорьях Нань-Шаня, видимо, проходит южная граница ареала этого вида (Magnusson, 1940). Более обширен ареал P. nigra Н. Magn. - от Монгольского Алтая, Котловины Больших Озер, Гобийского Алтая к пустыням Восточная Гоби и Заалтайская Гоби до южной части Алашаньской пустыни, гор Нань-Шаня и Цинхая (Magnusson, 1940; Голубкова, 1981).
Ареал P. mongolica Н. Magn. расположен в основном за пределами МНР - Цинхай, Ганьсу, Иньшань (Magnusson, 1940); северная часть ареала заходит в юго-восточную часть Гобийского Алтая (хр. Хурху) (Magnusson, 1940; Hanelt, Davazamc, 1965).
Более обширный ареал имеет, например, Endopyrenium atrocinereum (Н. Magn.) Golubk., развивающийся на гумусированных участках базальтовых и силикатных скал в пустынных и горных степях. На территории Монголии ареал этого вида охватывает Гобийский Алтай и Долину Озер, за пределами этой страны он известен в провинции Ганьсу (Китай) (Magnusson, 1940) и на Восточном Памире (Голубкова, 19716, 1973а).
Рис. 24. Распространение на территории Монголии Endocarpon pusillum Hedw. (1) и Е. sinense Н. Magn. (Ориг.)
3. Центральноазиатский тип ареала характерен для видов, встречающихся в пределах Монгольской, Тибетской и Джунгаро-Туранской провинций Центральной Азии (в понимании В. И. Грубова (1963)). Группа включает 20 видов - Endopyrenium inaequale (Н. Magn.) Golubk., E. bohlinii (H. Magn.) Golubk., Endocarpon sinense H. Magn. (рис. 24, 2), Peltula impressula (H. Magn.) Golubk. (табл. III, 1; рис. 25, 2), Collema kansuense H. Magn., Lecidea subconcava H. Magn., Squamarina pamirica Golubk., Aspicilia asiatica (H. Magn.) Oxn., A. verruculifera (Oxn.) Golubk., Polysporin gyrocarpa gobiensis H. Magn. (рис. 23, 1), A. invadens H. Magn., A. superans H. Magn., A. glypholecioides H. Magn. (табл. III, 4), A. bohlinii H. Magn., A. brevilobata H. Magn., Candelariella kansuensis H. Magn., Caloplaca bicolor H. Magn. и др.
В составе этой группы эпилитные, эпигейные и эпибриофитные лишайники, обитающие в горных, сухих и пустынных степях, в гобийских пустынях главным образом по склонам низкогорных хребтов. В числе их ряд видов играет заметную фитоценотическую роль в пустынно-степных растительных ценозах Монголии. К таким видам можно отнести Acarospora gobiensis Н. Magn. (табл. V, 1), A. bohlinii Н. Magn., Endopyrenium inaequale (Н. Magn.) Golubk., E. bohlinii (H. Magn.) Golubk., Endocarpon sinense H. Magn., Peltula impressula (H. Magn.) Golubk. и др.
Рис. 25. Распространение на территории Монголии Heppia lutosa (Ach.) Nyl. (1) и Peltula impressula (H. Magn.) Golubk. (2). (Ориг.)
Рассмотрим ареалы некоторых из этих видов. Ареал Peltula impressula (Н. Magn.) Golubk. (рис. 25, 2), растущего на почве, скалах, растительных остатках в пустынях, пустынных, сухих, а также горных степях, охватывает Монгольский Алтай, Среднюю Халху, Восточную Монголию, Котловину Больших Озер, Долину Озер, Джунгарскую, Заалтайскую, Алашаньскую и Восточную Гоби (Голубкова, 1981; Цогт, 1981). За пределами МНР этот вид известен в Северо-Западном Китае (Бей-Шань) (Magnusson, 1940), на территории СССР - в Центральном Тянь-Шане (Бредкина, 1981а). Инвазии P. impressula (Н. Magn.) Golubk. отмечены также в горных степях Хангая (Голубкова, 1981; Бязров и др., 1983).
Ареал Endopyrenium inaequale (Н. Magn!) Golubk. (рис. 26, 1), обитающего в пустынно-степных ценозах Монголии на скалах, в их расщелинах и на гумусированных участках, простирается от Монгольского Алтая, Средней Халхи, Долины Озер до Гобийского, Алтая и далее к югу до пустынь Джунгарская, Заалтайская Гоби, Галбын-Гоби, достигая за пределами МНР горных районов Нань-Шаня (Ганьсу) (Magnusson, 1940). На территории СССР этот лишайник встречается в горных и высокогорных степях Центрального Тянь-Шаня (Бредкина, 1981а) и на Восточном Памире (Голубкова, 1971б, 1973а).
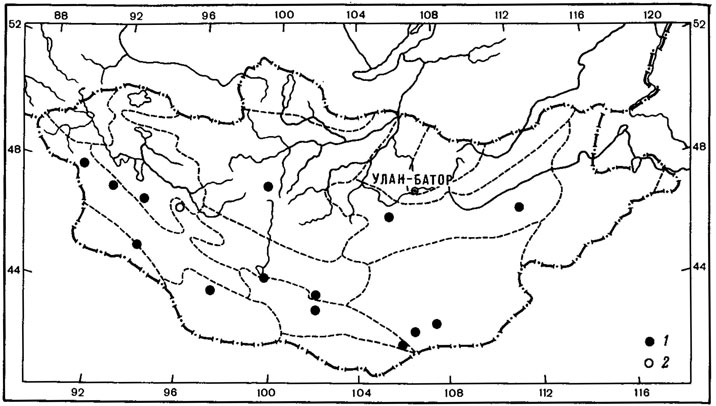
Рис. 26. Распространение на территории Монголии Endopyrenium inaequale (Н. Magn.) Golubk. (1) и Placidiopsis tominii Bredk. (2). (Ориг.)
Одним из широко распространенных лишайников юго-западных районов Монголии является Acarospora gobiensis Н. Magn. (табл. V, 1). Этот вид обильно развивается на поверхности скал по горным склонам в пределах высот 1000 - 2700 м над ур. м., образуя своими слоевищами крупные ярко-желтые пятна, хорошо заметные издали. На территории МНР ареал этого вида охватывает Джунгарскую, Заалтайскую и Восточную Гоби, Гобийский и Монгольский Алтай, западную часть Средней Халхи, на севере достигая долины р. Кобдо, Котловины Больших Озер и южных склонов Хангайского хребта. За пределами МНР этот лишайник часто встречается в восточной части Восточной Гоби (Magnusson, 1944), в Ганьсу (Magnusson, 1940), в восточной Кашгарии (Magnusson, 1929), Цинхае (Magnusson, 1940), в Джунгарии - в Джунгарском Алатау, Тянь-Шане, Джаркенской котловине (Голубкова, Шапиро, 1978; Бредкина, 1979; Голубкова, 1980б), проявляет инвазии в горы Южной Сибири, в Прибайкалье (Голубкова, Шапиро, 1978).
4. Ирано-туранский тип ареала характерен для 30 видов, произрастающих в пределах Ирано-Туранской области Древнесредиземноморского подцарства (в понимании A. Л. Тахтаджяна (1978). К этой группе видов относятся некоторые виды родов Endopyrenium - Е. desertorum (Tomin) Dzhur. (рис. 27, 1), Е. cinereofuscescens (Vain.) Oxn.; Endocarpon - E. alaicum Tomin, E. subfoliaceum Tomin; Acarospora - A. compacta H. Magn., A. ruf a (Vain.) H. Magn., A. fuscohepatica (Nyl.) Hue, A. tominiana H. Magn., A. bornmuelleri Steiner; Aspicilia - A. lacunosa (Ориг.). Mereschk., A. vagans Oxn.; Rinodina mucronatula H. Magn., Lecanora kukunorensis H. Magn., L. usbekica Poelt, Placidiopsis tominii Bredk. (рис. 26, 2), и др.
К сожалению, флора лишайников Ирано-Туранской области изучена далеко недостаточно: для большинства видов известны лишь отдельные местонахождения в пределах этого региона, по которым можно наметить приблизительные контуры ареалов того или иного вида. Наши слабые знания распространения лишайников в пределах Ирано-Туранской области не позволяют пока дать более дробную классификацию их ареалов; по-видимому, это дело будущего.
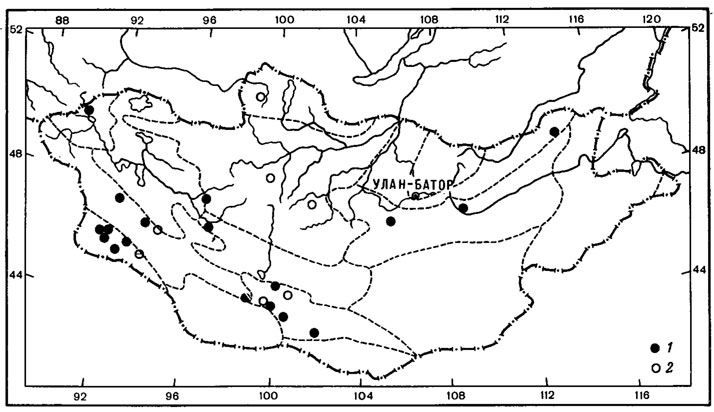
Рис. 27. Распространение на территории Монголии Endopyrenium desertorum (Tomin) Dzhur. (1) и Endocarpon alaicum Tomin (2). (Ориг.)
В качестве примера ирано-туранского ареала может служить распространение Endopyrenium desertorum (Tomin) Dzhur. (рис. 27, 1). Ареал этого вида охватывает территорию Волгоградской области и Восточного Кавказа, Казахстана, Туркменистана, Памиро-Алая, Тянь-Шаня, Монголии (Монгольский Алтай, Восточная Монголия, Котловина Больших Озер, Долина Озер, Гобийский Алтай, Заалтайская Гоби); этот лишайник встречается также в пустыне Негев в Израиле (Galun, 1970). Е. desertorum (Tomin) Oxn. дает инвазии в северные районы Монголии (Хангай) и в Южную Сибирь (Алтай), в среднее течение р. Индигирки (Афонина и др., 1979).
5. Древнесредиземноморский тип ареала присущ 18 видам, распространенным в пределах Древнесредиземно- морского подцарства Голарктики (в понимании А. Л. Тахтаджяна (1978)). В составе группы Lichinella stipatula Nyl., Gonohymenia myriospora Zahlbr., G. algerica Steiner, Endopyrenium trachyticum (Vain.) Hazsl., Peccania coralloides Massal., Peltula radicata Nyl., Acarospora hilaris (Duf.) Hue, Teloschistes contortuplicatus (Ach.) Clauz. et Rond., T. lacunosus (Rupr.) Savicz, Squamarina gypsacea (Sm.) Poelt, Aspicilia reticulata Krempelh., Physconia servitii (Nadv.) Poelt, Caloplaca conglomerata (Bagl.) Jatta, Toninia tabacina Flag., Chrysopsora testacea (Hoffm.) Choisy и др.
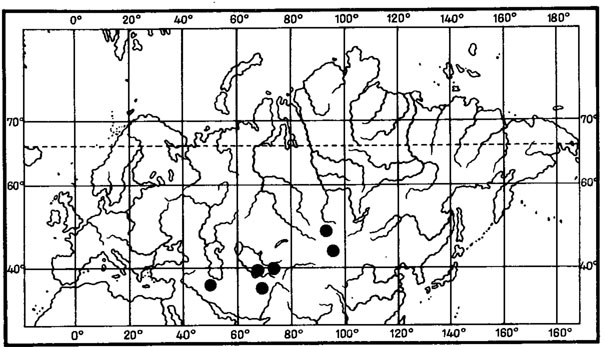
Рис. 28. Распространение на земном шаре Acarospora stapfiana (Mull. Agr.) Hue. (Ориг.)
Примером древнесредиземноморского ареала может служить распространение Peccania coralloides Massal. Ареал этого вида охватывает средиземноморские районы Европы, Северной Африки, Среднюю Азию и Монголию (Котловина Больших Озер, Долина Озер, Гобийский Алтай, Джунгарская, Заалтайская и Восточная Гоби). Инвазии этого вида отмечены в северных районах Монголии (Прихубсугулье), а также в Средней Европе. Некоторые виды древнесредиземноморских лишайников, например Caloplaca conglomerata (Bagl.,) Jatta (рис. 35), имеют ареалы с широкими дизъюнкциями. Этот вид распространен в средиземноморских районах Европы, проникает к северу в теплые альпийские долины (Poelt, 1969), известен в Израиле (Верхняя Иорданская долина) (Galun, 1970), а после обширной дизъюнкции часто встречается в степях восточных районов Монголии, в предгорьях Хэнтэя, в долинах рек Онон, Керулен и Улдза, в Прихинганье, в восточных районах Средней Халхи и в Восточной Монголии. Такие ареалы с огромными дизъюнкциями, характерные для многих видов древнесредиземноморских лишайников, свидетельствуют о древнем происхождении этих видов.
6. Сонорско-древнесредиземноморский тип ареала присущ видам, обитающим в пределах Древнесредиземноморского и Сонорского (Мадреанского) подцарств Голарктики (в понимании A. Л Тахтаджяна (1978)). Группа включает 10 видов- Acarospora bullata Anzi (рис. 29), Colle,ma ryssoleum (Tuck.) A. Schneid., Psora globifera (Ach.) Massal., Lecanora bolcana (Pollin.) Poelt, Acarospora heutleriana Koerb., Glypholecia scabra (Pers.) Müll. Arg и др.
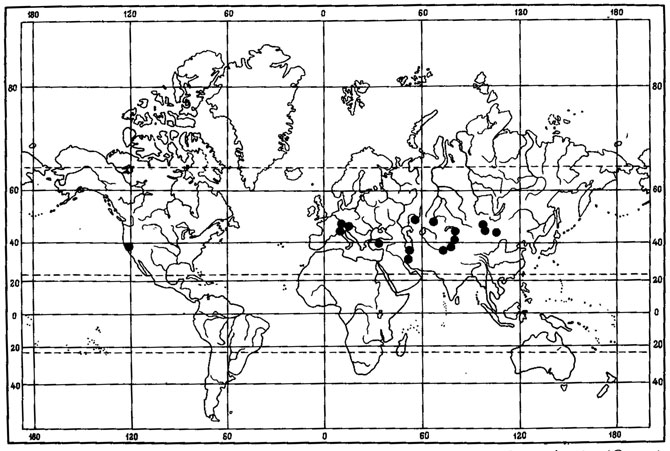
Рис. 29. Распространение на земном шаре Acarospora bullata Anzi. (Ориг.)
В качестве примера может служить ареал Acarospora heufleriana Koerb. (рис. 41). Этот вид в Европе встречается в Южной Франции и Северной Италии (Magnusson, 1929, 1936а; Poelt, 1958; Leuckert, 1968; Poelt, Steiner, 1971), на Южном берегу Крыма (Окснер, 1968), известен в Северной Африке - Марокко (Werner, 1967), на Кавказе (Бархалов, 1944; Челидзе, 1971), в Азии - в Северном Иране (Окснер, 1946б), в горах Копетдага (Джураева, 1978); после обширной дизъюнкции встречается в Гобийском Алтае, в хр. Хурху (Голубкова, 1980б), а также в Восточной Гоби (Magnusson, 1944). В Северной Америке найден в штате Нью-Мексико (Magnusson, 1937).
7. Сонорско-центральноазиатский тип ареала характеризуется дизъюнктивным ареалом, одна часть которого находится в пределах Сонорского подцарства Голарктики (в понимании Тахтаджяна 1978)), другая - на территории Центральной Азии. Представлен ареалами 3 видов - Acarospora peltastica Zahlbr., A. cinereoalba (Fink) H. Magn., A. nevadensis H. Magn. В пределах МНР A. peltastica Zahlbr. найдена лишь однажды в горных полынно-злаковых степях в хр. Адж-Богдо на высоте 3000 м над ур. м.; в Северной Америке этот вид встречается в Калифорнии (Magnusson, 1929). A. cinereoalba (Fink) Н. Magn. на территории Монголия отмечена в степях Средней Халхи и Восточной Монголии, приводится для Восточной Гоби в пределах Внутренней Монголии (Magnusson, 1944), в Северной Америке известна из штата Нью-Мексико, откуда проникает в более северные районы, в штат Иллинойс (Magnusson, 1929).
A. nevadensis Н. Magn. также редкий на территории Монголии вид, собранный в Гобийском Алтае (Голубкова, 1981). В Азии ранее он был отмечен для Восточного Памира (Голубкова, 1972, 1973а), а также для Зеравшанского хребта. В Северной Америке встречается в горах штата Невада (Magnusson, 1929).
8. Хэнтэе-хангайский тип ареала свойствен одному виду: Aspicilia changaica (Klem.) Golubk. Этот вид является эндемиком горно-степных районов Монголии. В Хангае и Хэнтэе он обитает на горизонтальных поверхностях скал в горных степях, иногда проникая в высокогорный пояс, изредка в лиственничных лесах.
9. Забайкальско-хангайский тип ареала характерен для одного вида - Aspicilia mikhnoi (Zahlbr.) Oxn., распространение которого ограничено горно-степными районами Забайкалья (Zahlbruckner, 1911), а на территории Монголии - Хангаем, хотя, возможно, будет найден и в других районах этой страны. Как отмечал Л. Г. Бязров (Бязров и др., 1983), в Хангае этот лишайник является редким, растущим на выходах горных пород в степных и высокогорно-луговых сообществах на склонах и вершинах сопок, гребнях хребтов.
10. Сибирско-центральноазиатский тип ареала присущ 4 видам - Lecanora baicalensis Zahlbr., Acarospora jenisejensis H. Magn., A. sibirica H. Magn., Lecidea asiaecentralis H. Magn.
Основная часть ареала Lecanora baicalensis Zahlbr (рис. 30, 2; табл. 1, 4) связана с территорией МНР, где этот лишайник часто встречается на скалах и валунах в горах, горных и сухих степях, поднимаясь в горах до высокогорного пояса, в Хэнтэе, Хангае, Монгольско-Даурском районе, Монгольском Алтае, Средней Халхе, в Гобийском Алтае (Голубкова, 1981), проникая на севере за пределы Монголии в горные степи Забайкалья (Zahlbruckner, 1911), а на юге - на территорию Внутренней Монголии (Китай) (Magnusson, 1944). Кроме того, L. baicalensis Zahlbr. приводится также для Каракорума (Poelt, 1961) и холодных пустынь Памира (на территории СССР и Афганистана) (Poelt, Wirth, 1968; Голубкова, 1973а).
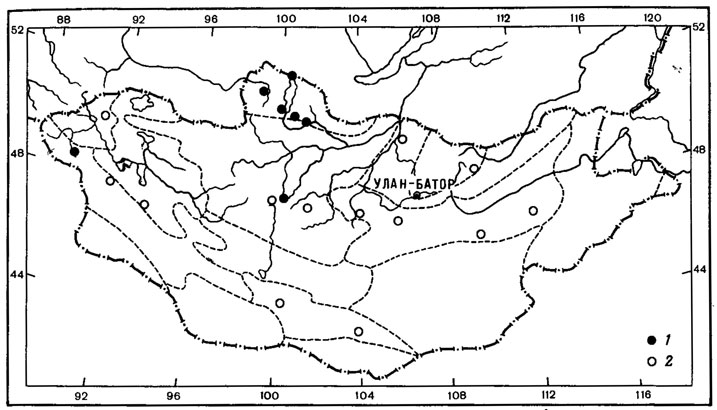
Рис. 30. Распространение на территории Монголии Lecanora epibryon (Ach.) Ach. и L. baicalensis Zahlbr. (2). (Ориг.)
Lecidea asiaecentralis Н. Magn. - чрезвычайно широко распространенный в Монголии лишайник, обитающих на почве, растительных остатках, у основания и в расщелинах скал в горных и равнинных степях и пустынях (рис. 31). Ареал этого лишайника охватывает всю территорию Монголии, от районов Прихубсугулья на севере до пустынь Джунгарская и Заалтайская Гоби на юге и от Монгольского Алтая на западе до района Прихинганья на востоке. За пределами Монголии он простирается на территории Внутренней Монголии и провинции Ганьсу, (Magnusson, 1940, 1944), а также Центрального Тянь-Шаня (Бредкина, 1979). Изолированное местонахождение этого вида известно в среднем течении р. Индигирки (Афонина и др., 1979).
Рис. 31. Распространение на территории Монголии Lecidea asiaecentralis H. Magn. (Ориг.)
Acarospora jenisejensis Н. Magn. рассеянно встречается на территории Северо-Западной Монголии, в Хангае, Гобийском Алтае, Прихинганье и Восточной Монголии, где обитает на скалах в горах, горных и сухих степях (Голубкова, 1981), проникая в Китае в район Внутренней Монголии и Ганьсу (Magnusson, 1940, 1944). В Южной Сибири этот лишайник известен из окрестностей Красноярска (Magnusson, 1936b). Ареал Acarospora sibirica Н. Magn. на территории Монголии связан с горными степями Средней Халхи и Долины Озер, за пределами Монголии этот лишайник известен в степях Хакасии (Magnusson, 1929).
11. Азиатский тип ареала свойствен 2 видам лишайников - Diploschistes steppicus Reichert и Parmelia tominii Oxn.
Ареал Diploschistes steppicus Reichert, растущего на почве и растительных остатках в степях и лугах, охватывает Монголию (Хангай, Прихинганье), Кавказ, Малую Азию и п-ов Аравия, в Восточной Сибири известен из среднего течения р. Индигирки (Афонина и др., 1979).
Ареал Parmelia tominii Oxn. простирается в Западной и Восточной Сибири, Средней Азии, охватывает всю территорию Монголии.
12. Восточноевропейско-азиатский тип ареала характерен для Aspicilia transbaicalica Oxn. и A. maculata (Н. Magn.) Oxn.
A. transbaicalica Oxn. встречается в степных районах Поволжья (Куйбышевская, Ульяновская области, Башкирская АССР), на Кавказе, в Казахстане (Андреева, 1961; Бредкина, Макарова, 1971), в Горной Шории (Седельникова, 19776), на западном побережье оз. Байкал (Рассадина, 1936), в Забайкалье (Окснер, 1939а), в Монголии (Голубкова, 1981), в Центральном Тянь-Шане (Бредкина, 1979, 1981а, 19816), на Восточном Памире (Голубкова, 1973а) и в Китае (Шаньси) (Wej, 1981). На территории Монголии этот вид обитает на валунах, выходах горных пород в горных степях Хэнтэя, Монгольско-Даурского района, Средней Халхи (рис. 42).
Основной ареал A. maculata (Н. Magn.) Oxn. связан с Центральной Азией, где этот вид часто встречается в центрально- азиатской части Монголии (Монгольский Алтай, Средняя Халха, Восточная Монголия, Котловина Больших Озер, Восточная Гоби, Долина Озер, Гобийский Алтай, Джунгарская и Заалтайская Гоби) (Голубкова, 1981), а также в Китае - во Внутренней Монголии, Цинхае (Magnusson, 1940), в СССР - на Восточном Памире (Голубкова, 1973а), а северо-восточной части Афганистана (Poelt, Wirth, 1968), северо-западных Гималаях (Awasthi, 1963, 1965). Этот вид приводится также для юго-восточных районов Европы - Донецкой области и. Приазовья (Коваленко, 1976, 1977).
13. Палеарктичекский тип ареала присущ 3 видам - Aspicilia farinosa Arnold, A. radiosa (Hoffm) Poelt et Leuck., Diploschistes clausus (Koerb.) Zahlbr.
Эпилитный лишайник Aspicilia farinosa Arnold распространен в средиземноморских районах Европы (Греция, Италия, Франция) и Северной Африки (Марокко, Алжир); он проникает на территорию Европы в более северные районы - в Румынию, Венгрию, Польшу, Прибалтику, а также на Украину, в Уьяновскую область. В Азии он приводится для Сирии, Израиля, Ирана, Кавказа, Туркмении. После значительной дизъюнкции этот лишайник встречается в Монголии, в центральном Хангае (Schubert, Klement, 1971), а также в Центральном Тянь-Шане.
14. Голарктический тип ареала характерен для видов, встречающихся в аридных районах Голарктики. Группа включает 15 видов - Squamarina lentigera (Web.) Poelt, Parmelia taractica Krempelh., P. tinctina Mah. et Gill., Synalissa ramulosa Fr., Thyrea pulvinata (Schaer.) Massal., Aspicilia alphoplaca (Wahlenb.) Poelt et Leuck., A. praeradiosa (Nyl.) Poelt et Leuck. (табл. 11, 2), A. desertorum (Krempelh.) Mereschk., Ramalina capitata (Ach.) Nyl., Anaptychia ulothricoides (Vain.) Vain., Caloplaca tominii Savicz и др.
Лишайники этой группы, как и виды с ареалами палеарктиче- ского типа, иррадиируют в северные районы Голарктики, достигая в Европе Скандинавии, а вместе со степными комплексами растительности проникают далеко на север, в горные районы Восточной Сибири, где обитают в подходящих ксерических экотопах. Например, Squamarina lentigera (Web.) Poelt, обитающая на карбонатных почвах, известняковых скалах, реже на мхах, известна в средиземноморских районах Европы и Северной Африки, рассеянно встречается в степных и полупустынных районах Средней Европы, Украины, Заволжья, Кавказа, Средней Азии, достигая на востоке Саян, Монголии и Якутской АССР. На севере S. lentigera (Web.) Poelt проникает в Эстонию и Южную Норвегию (острова Эланд, Готланд). В Северной Америке известна только в Скалистых горах и на р. Юкон. На территории МНР - редкий лишайник, собранный в Прихубсугулье и Долине Озер.
Б. Омниаридный субэлемент объединяет 20 видов. Для них характерен один тип ареала.
1. Мультирегиональный тип ареала характеризует виды, обитающие в теплоаридных областях Голарктики и других флористических царств. Группа включает 20 видов - Le- canora frustulosa (Dicks.) Ach., Parmelia pulla Ach., P. glabrans Nyl. (рис. 32), P. vagans Nyl., Psora decipiens (Hedw.) Hoffm., Fulgensia fulgens (Sw.) Elenk., F. bracteata (Hoffm.) Ras., Acarospora schleicheri (Ach.) Massal., A. sulphurata Arnold (рис. 33), A. strigata (Nyl.) Jatta, Peltula obscurans Gyeln., P. euploca (Ach.) Wetm., Endopyrenium hepaticum (Ach.) Koerb. и др.
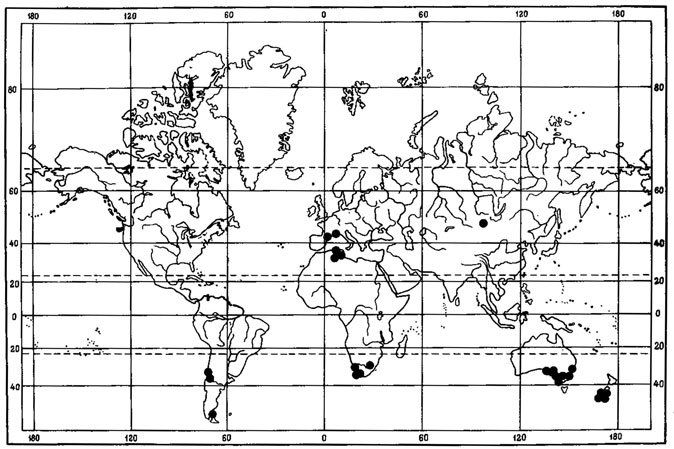
Рис. 32. Распространение на земном шаре Parmelia glabrans Nyl. (По: Eslinger, 1977, с дополнениями автора)
Примером может служить обширный дизъюнктивный ареал Acarospora schleicheri (Ach.) Massal., распространение которого на земном шаре проиурочено к степным и полупустынным растительным сообществам аридных районов (Голубкова, Шапиро, 1978) (рис. 34) . Этот вид часто встречается в средиземноморских районах Европы и Северной Африки (Magnusson, 1929, 1936а; Crespo, Barreno, 1970). После обширной дизъюнкции на Африканском континенте широко представлен в степных и пустынных районах Южной Африки, в районе бассейна р. Оранжевой, в областях Высокий Велд и Бушвелд, в пустыне Намиб, в Капской провинции (ЮАР) (Almborn, 1966). Восточнее средиземноморских районов A. schleicheri (Ach.) Massal. встречается в низовьях Волги, в Прикаспийской низменности, в Северном и Центральном Казахстане, Южной Сибири, Монголии (Голубкова, Шапиро, 1978; Голубкова, 1980б), южнее - на равнинах юго-западной Туркмении (Джураева, 1974) и в северо-западном Каракоруме (Poelt, 1961). В Северной Америке основное распространение вида связано с североамериканскими прериями. В Канаде он известен в провинции Саскачеван, на территории США - в горах Блэк-Хилс, восточных отрогах Скалистых гор, в Техасе, на плато Колорадо, а также в южной Калифорнии (Magnusson, 1929; Fink, 1935; Howard, 1950; Egan, 1970; Wetmore, 1970, и др.). Co степными комплексами A. schjejcheri (Ach.) Massal. довольно далеко заходит на север, на плато Лено-Амгинского междуречья и в долину р. Индигирки (Караваев, 1976; Афонина и др., 1979), отмечен также на юго-восточном побережье Гренландии (Magnusson, 1929; Lynge, 1940). В южном полушарии, помимо Южной Африки, встречается в Южной Америке, в Чили, в пустыне Атакама (Rendon, 1973). На территории Монголии A. schleicheri (Ach.) Massal. обитает на почве и растительных остатках в горных злаково-разнотравных, ковыльно-карагановых, холодно- полынно-типчаковых и осоково-кобрезиевых степях западных и центральных горно-степных и лесостепных районов. Южнее, в пустынно-степных и пустынных районах Монголии, этот вид не найден; неизвестен он и в Восточной Монголии. Обширный ареал этого лишайника, охватывающий северное и южное полушария и прерываемый широкими дизъюнкциями, свидетельствует о древнем происхождении вида. Имеются основания предполагать, что A. schleicheri (Ach.) Massal. является древним третичным видом, дошедшим до нас в неизменном состоянии. Видимо, это один из представителей древней ксерофильной флоры лишайников, центром происхождения которой были районы Южной и Юго-Западной Африки.
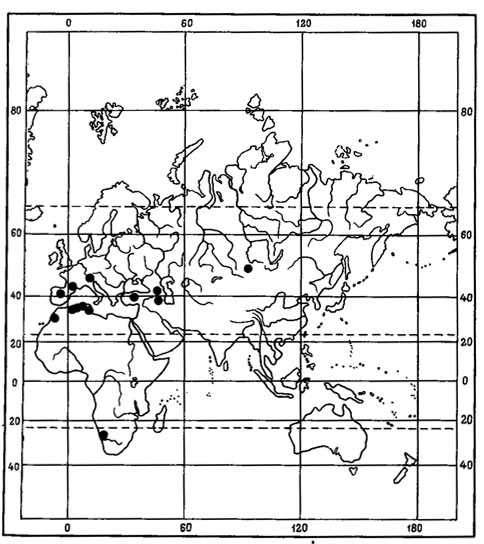
Рис. 33. Распространение на земном шаре Acarospora sulphurata Arnold. (Ориг.)
Охарактеризуем ареал другого представителя желтоокрашенных акароспор - Acarospora sulphurata Arnold. Этот лишайник довольно широко распространен в Северной Африке (Алжир, Тунис, Марокко), где он растет как в прибрежных, так и во внутренних районах. Так, Вернер (Werner, 1967) приводит A. sulphurata Arnold для степных и пустынных районов Марокко. Севернее этот вид встречается в Южной Европе, в восточных Пиренеях (Франция), в Иберийских горах (Испания), в южном Тироле, в хр. Вингачу (Северная Италия) (Magnusson, 1929, 1936; Clauzade е. а., 1981). На территории Азии он известен в Малой Азии, в Турции (Magnusson, 1929) и на Кавказе, в Азербайджане (Бархалов, 1955). После значительного разрыва в ареале этот лишайник появляется в Монголии, в Котловине Больших Озер, где был найден в нанофитовой пустыне в окрестностях г. Улангом и в горной степи в окрестностях Тургэн сомона (Klement, 1965; Schubert, Klement, 1971). Одна из разновидностей этого вида - A. sulphurata var. austroafricana Zahlbr. - встречается в Юго-Западной Африке, тем самым фиксируя древние флористические связи Монголии с пустынными районами Южной Африки, а также свидетельствуя о древнем происхождении этого вида. Разрыв в ареале A. sulphurata Arnold на территории Азии мог произойти на рубеже третичного и четвертичного периодов, но дефект в ареале на Африканском континенте относится, видимо, к гораздо более раннему времени.
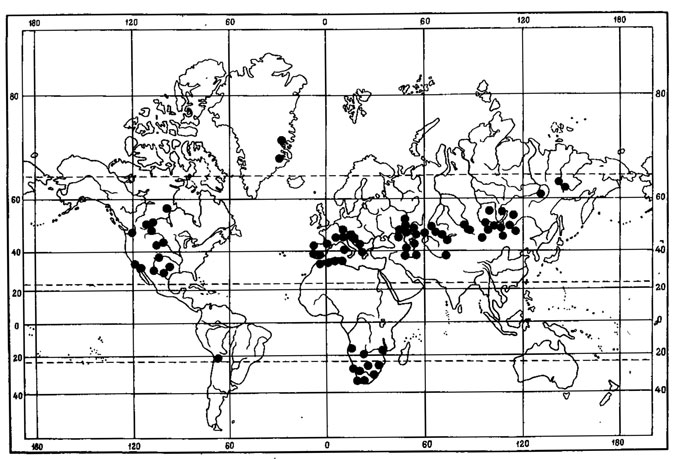
Рис. 34. Распространение на земном шаре Acarospora schleicheri (Ach.) Massal (Ориг.)
Таким образом, подводя итоги изучения ареалов лишайников аридного элемента, можно заключить, что в его составе преобладают виды с ареалами, ограниченными территориями Древнего Средиземья (100 видов, 67.1% от общего числа видов аридного элемента). Это позволяет предположить древнесредиземномор- скую основу в его развитии. Более половины этих видов (53 вида, или 31.8%) имеет ареалы, ограниченные территориями различных провинций Центральной Азии, при этом преобладают монгольские эндемичные виды (21 вид, или 19.6% от общего числа видов аридного элемента). Это дает основание предполагать значительную роль автохтонного элемента в формировании аридной лихенофлоры изучаемого района. 30 видов (18.1%) проявляет ирано-туранские флористические связи, 18 видов (10.8%) - широкие древнесредиземноморские флористические связи, 10 видов (8.3%) фиксируют связи с флорой Северной Америки. Беднее представлены в аридном элементе связи с флорами Южной Сибири (17 видов, или 10.7%) и понтическими районами Европы (3 вида, или 2%). Немногочисленны среди представителей аридного элемента виды с голарктическими ареалами (15 видов, или 9%). Древние связи аридной флоры Монголии с флорами аридных районов Южной Африки, Южной Америки и Австралии фиксируют 20 видов (или 12%).
VI. Высокогорный элемент. Объединяет 19 видов (или 2.8%), основное распространение которых связано с высокими горами Голарктики (голарктический высокогорный субэлемент), а также с другими флористическими царствами (омнивы-сокогорный субэлемент), где эти виды растут в высокогорных безлесных поясах.
В состав высокогорного элемента входят представители 8 семейств лишайников: Lecideaceae (6 видов) и Acarosporaceae (3), Lecanoraceae (2), Microglaenaceae, Aspiciliaceae, Parmeliaceae, Usneaceae, Teloschistaceae, Umbilicariaceae (по 1 виду каждое).
А. Голарктический высокогорный субэлемент охватывает 19 видов. Для ареалов лишайников этого субэлемента можно наметить следующие типы.
1. Ирано-туранский тип ареала свойствен одному виду - Acarospora stapfiana (Mull. Arg.) Hue, распространенному в высокогорьях Ирано-Туранской области Древнесредиземноморского подцарства Голарктики (в понимании А. Л. Тахтаджяна (1978)).
Ареал этого вида на территории Монголии охватывает Монгольский Алтай, где он был собран в хребтах Адж-Богдо и Сутай-Ула, в криофитных степях, на высоте 2730 - 3000 м над ур. м. За пределами Монголии этот лишайник встречается в Центральном Тянь-Шане, в высокогорьях Памира (на территории СССР и Афганистана) (Poelt, Steiner, 1971; Голубкова, 1972, 1973а; Голубкова, Шапиро, 1978), Зеравшанского хребта (Кудратов, 1977б; 1981; Голубкова, Шапиро, 1978), в горах Акташ Узбекистана (Pisut, 1978), а также в горах Ирана (Miiller, 1892; Magnusson, 1929; Szatala, 1957). Этот вид из группы желтоокрашенных акороспор подрода Xanthothallia обычно обитает на слоевищах других лишайников, чаще всего видах секции Gasparrinia рода Caloplaca, реже на поверхности горных каменистых пород. В Монголии, как и на Памире, он обитает на талломе Caloplaca hedinii Н. Magn. (рис. 23,3; 28) (Голубкова, Шапиро, 1978; Голубкова, 1980б, 1981).
2. Хангайско-центральноазиатский тип ареала присущ одному виду - Caloplaca hedinii Н. Magn. (рис. 35,2). Основное распространение этого лишайника связано с высокогорными районами Центральной Азии, где он обитает в горах Нань-Шаня, в пределах высот 2700 - 3700 м над ур. м. (Magnusson, 1940), Памира (на территории СССР и Афганистана) (Poelt, Wirth, 1968; Голубкова, 1973а), Центрального Тянь-Шаня, а также на территории МНР в Монгольском Алтае (Голубкова, 1981). Этот вид был найден в хребте Сутай-Ула, на высоте 2730 м над ур. м., и в хр. Адж-Богдо, у подножия горы Их-Обо-Ула, на высоте 3000 м над ур. м. Кроме того, этот лишайник был собран в районе магистрального хребта Хангая, в окрестностях оз. Хух-Нур, на высоте 2700 м над ур. м., в злаково-типчаковой степи (Голубкова, 1981). Недавно был указан для нагорья Сангилен (Тува) (Седельникова, 1983).
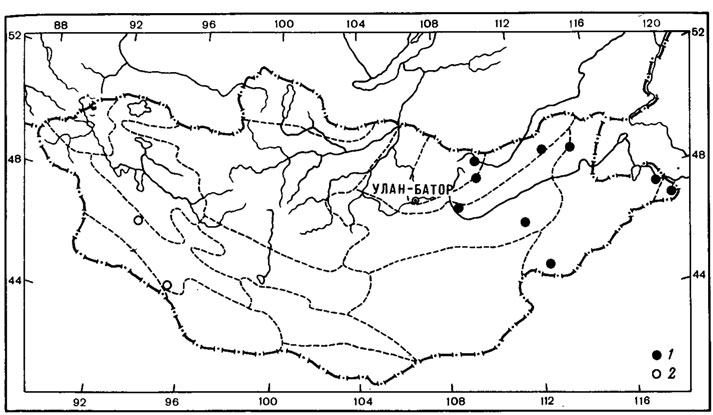
Рис. 35. Распространение на территории Монголии Caloplaca conglomerata (Bagl.) Jatta (1) и С. hedinii Н. Magn. (2). (Ориг.)
3. Хангайско-гималайский тип ареала свойствен одному виду - Lecanora tschomolongmae Poelt. До находок этого вида в юго-западной части Хангая (Schubert, Klement, 1971) он был известен из Гималаев (Poelt, 1966), где был собран на гнейсовых скалах на высоте 4950 - 5000 м над ур. м.
4. Хангайско-алтайский тип ареала включает ареал одного вида - Evernia terrestris (Tomin) Golubk. Этот эпигейный кочующий лишайник на территории Монголии обитает в высокогорных степях Хангая и Монгольского Алтая. В Хангае он был собран в злаково-типчаковых степях в окрестностях оз. Хух-Нур, на высоте 2700 м над ур. м., в Монгольском Алтае - в злаковых горных степях в хр. Хасагт-Хайрхан-Ула, на высоте 2970 м над ур. м. (Голубкова, 1981), а также в окрестностях оз. Холбо-Нур (Томин, 1933). За пределами Монголии этот вид отмечен на Алтае (Рассадина, 1938), где он обитает на ледниковых моренах и в альпийской тундре, в истоках р. Канаса, а также в Казахстане в долине р. Каджар (Томин, 1933).
Выше были показаны филогенетические связи Evernia terrestris (Tomin) Golubk. с лесной эпифитной формой Е. esorediosa (Miill. Arg.) DR. Как мы полагаем, дифференциация этого крио-ксерофитного вида от исходной мезофильной формы, возможно, могла произойти в начале плейстоцена, во время деградации древних лесных третичных типов, и процесс формирования его в самостоятельный вид проходил в условиях криоксерофитных степей плейстоцена в горных районах Монголии и Южной Сибири.
5. Тибетско-хангайско-алтайский тип ареала характерен для одного вида - Cetraria potaninii Охn. (рис. 19, 2). Этот свободноживущий кочующий эпигейный лишайник обитает в злаково-типчаковых и полынно-злаковых горных степях Хангая и Монгольского Алтая (Голубкова, 1981), ранее был известен с Тибета. С. potaninii Oxn. можно причислить к группе видов, выявленной среди цветковых растений (Грубов, 1976), дизъюнктивные ареалы которых указывают на существовавшие, вероятно в плейстоцене, связи высокогорных флор Хангая и Саян, с одной стороны, и Тибета и Гималаев, с другой.
6. Кавказско-алтайский тип ареала также характеризуется дизъюнктивным ареалом, одна часть которого расположена на Кавказе, другая - в горах Монгольского Алтая. Он присущ одному виду - Aspicilia szechenyi (Vain.) Hue.
7. Евразиатский тип ареала свойствен видам, произрастающим в высокогорьях Европы и Азии. Группа включает 9 видов - Lecidea leptoboloides Nyl., L. paratropoides Mull. Arg., L. steineri Hertel, Microglaena corrosa Arnold, и др.
Примером может служить ареал Lecidea paratropoides Mull. Arg. (рис. 36), охватывающий горы Пиренейского полуострова, Альпы, горы Македонии (Балканский полуостров) и Центральной Азии (Памиро-Алай, Западный Тянь-Шань, горы Афганистана, Монгольский Алтай, Хангай, северо-западные Гималаи) (Hertel, 1977). Изредка этот лишайник встречается в горно-степных районах и в сухих степях Средней Халхи (МНР) и Внутренней Монголии (Китай).
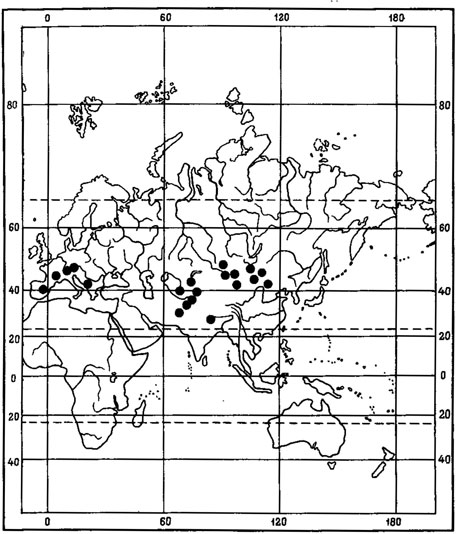
Рис. 36. Распространение на земном шаре Lecidea paratropoides Mull. Arg. (По: Hertel, 1977)
8. Голарктический тип ареала характерен для 2 видов, распространенных в высокогорьях Голарктики - Lecidea athroocarpa (Ach.) Ach. и L. marginata Schaer.
Например, ареал Lecidea athroocarpa (Ach.) Ach. охватывает Северо-Западную Европу (Фенноскандию, Великобританию, СССР - КАССР), горы Франции, Альпы, Тироль, горы южной части ФРГ и ГДР, Польши (Силезия), Чехословакии, Карпаты, горы Северной Африки, Кавказ (Окснер, 1968), в Азии - Алтай, горы Монголии, Гималаи и северо-западную часть Юньнаньского нагорья (Hertel, 1977; Седельникова, 1977б). Этот лишайник заходит и в западное полушарие, в горные районы Мексики. В Гималаях известен с очень больших высот - 5200 м над ур. м. В МНР найден в горах Прихубсугулья, в Хэнтэе и Хангае.
Б. Омнивысокогорный субэлемент.
1. Азиатско-восточноафриканский тип ареала свойствен одному виду - Lasallia pertusa (Rassad.) Llano. Этот лишайник часто встречается в горных районах Монголии (Прихубсугулье, Хэнтэй, Хангай, северная часть Монгольского Алтая, Котловина Больших Озер, Средняя Халха), где обитает на скалах по склонам гор в лиственничных лесах, реже в горных степях, в пределах абсолютных высот 1300 - 2300 м. L. pertusa (Rassad.) Llano известен в Прибайкалье, на западном побережье оз. Байкал и на о-ве Ольхон (Рассадина, 1936). Вторая часть ареала связана с Гималаями - северо-западные Гималаи, Кашмирские и в пределах Непала (Llano, 1950; Frey, Poelt, 1977). Третья часть ареала этого вида находится в палеотропиках, в горах Эфиопии (Krog, 1973). В Гималаях и горах Эфиопии этот лишайник обитает на высоте 3600 - 4300 м над ур. м. Таким образом, L. pertusa (Rassad.) Llano, с одной стороны, фиксирует географические связи флоры горных районов Монголии с Гималаями, существовавшие, как полагает В. И. Грубов (1976), вероятно, в плейстоцене, с другой, он указывает, по-видимому, на более древние связи с флорой Африканского континента.
2. Евразиатско-южноамерика некий тип ареала характерен для одного вида - Lecidea secernens Н. Magn. Дизъюнктивный ареал этого вида охватывает горные районы Швеции, Центральной Азии (Гималаи, горы Монголии) и Анды в Южной Америке (Hertel, 1977).
Таким образом, более чем третья часть видов высокогорного элемента (6 видов, или 35.2%) характеризуются ареалами, ограниченными высокогорьями Азии. При этом они фиксируют флористические связи высокогорных районов Монголии с высокогорными флорами Тибета, Гималаев, Юньнаньского нагорья, Тянь-Шаня, Памиро-Алая, Ирана, Кавказа, а также Алтая. 9 видов высокогорного элемента (или 47.3%) имеет евразиатские ареалы, фиксируя географические связи флоры Монголии за пределами Азии, с высокогорными флорами Европы. 2 вида проявляют широкоголарктические флористические связи, и 2 вида фиксируют, видимо, древние флористические связи горных районов Монголии с флорами Африки и Южной Америки.
VII. Монтанный элемент. Советские лихенологи относят к монтанному элементу виды, произрастающие в лесных поясах гор Голарктики (Трасс, 1970). М. Ф. Макаревич (1963, см. выше) так характеризовала монтанный элемент: "К монтанному элементу мы относим растения, которые растут в условиях горных лесов (лесных поясов) гор Голарктики, нередко снижаясь в предгорья, поднимаясь выше в безлесные пояса и снижаясь на равнины".
Однако, как отмечал А. А. Юнатов (1950), резкая континентальность климата налагает особый отпечаток на характер смен вертикальных поясов в горах МНР. Только в редких случаях можно наблюдать "нормальные", полные, ряды поясов. Гораздо чаще те или иные пояса (особенно лесной) выпадают, и тогда обнаруживается непосредственный контакт, например, высокогорного пояса со степью или даже с пустынной степью. Пестрота поясного распределения растительности объясняется сложностью горного рельефа. Ведущую роль здесь приобретает экспозиция горных склонов. Наиболее наглядно ее влияние проявляется в Северной Монголии, где леса приурочены исключительно к северным склонам, а южные совершенно безлесны. Аналогичные соотношения наблюдаются и в смежных районах Забайкалья. Вместе с тем среди лишайников МНР выделяется группа видов, которые встречаются лишь в горах, в среднем горном поясе, на низкогорных хребтах, в основном на скальных обнажениях, щебнистых и мелко-земистых субстратах петрофильных растительных сообществ. Исходя из специфики природных условий горных районов МНР, нам кажется более приемлемым понимание объема монтанного элемента А. С. Лазаренко (1956), который писал, что, кроме горных лесных видов, в сложении монтанного элемента принимают участие скальные и водные виды, которые выразительно оттеняют специфику элемента. Эти виды, как правило, ограничены средним горным поясом и почти не заходят в высокогорья.
Монтанный элемент объединяет во флоре лишайников МНР 88 видов (11.6%).
А. Голарктико-монтанный субэлемент. Лишайники этого субэлемента в своем распространении связаны с горами Голарктики. Им свойственны следующие типы ареалов.
1. Прибайкальско-хангайский тип ареала присущ одному виду - Usnea capitata Ras. О распространении этого вида данных пока недостаточно; он известен лишь из двух местонахождений - западного побережья оз. Байкал, где встречается по скалистым берегам близ улуса Сахюрта (Ольхонские ворота) (Рассадина, 1936), и из Хангая, где обитает на каменистом субстрате в лиственничных лесах среднего пояса гор (Бязров, 1974а).
2. Саянский тип ареала присущ одному виду - Acarospora angusta Н. Magn. Как и для Usnea capitata Ras., данных о распространении этого лишайника очень мало, и контуры ареала вида намечены предварительно. До находок его на территории Монголии, в районе Прихубсугулья, в бассейне р. Аригийн-Гол (Голубкова, 1981), он был известен из верховья р. Енисея (Magnusson, 1939b).
3. Алтайско-хангайско-юньнаньский тип ареала свойствен 2 видам - Squamarina callichroa (Zahlbr.) Poelt и Buellia cervinoplaca Zahlbr. Одна часть ареала этих видов охватывает Монгольский Алтай и Хангай, другая - Юньнацьское нагорье (Zahlbruckner, 1930; Poelt, 1958; Schubert, Klement, 1971).
4. Азиатский тип ареала характерен для 3 видов - Psora pulcherrima (Vain.) Elenk., Lecanora crustacea (Savicz) Zahlbr., Parmelia borisorum Oxn.
Ареал Psora pulcherrima (Vain.) Elenk. охватывает Закавказье - Талыш (Бархалов, 1975), Копетдаг (Vainio, 1896), Саяны (Еленкин, 1902), Монголию - горные районы Прихубсугулья, Забайкалье - хр. Черского, Приморский край - хр. Сихоте-Алинь (Окснер, 1940).
5. Азиатско-американский тип ареала присущ 3 видам - Peltigera mauritzii Gyeln., Anema jenisejensis H. Magn., Lecidella alaiensis (Vain.) Hertel.
Peltigera mauritzii Gyeln. - один из широко распространенных в МНР лишайников. Он встречается в Прихубсугулье, Хэнтэе, Прихинганье, Монгольском Алтае и Средней Халхе, где обитает в горах на почве среди мхов, замшелых скалах. Помимо Монголии, ареал этого вида охватывает Кавказ, горы Западной и Восточной Сибири, Средней Азии, Камчатки, Дальнего Востока и Китая, а также горные районы Северной Америки.
Anema jenisejensis Н. Magn. - редкий лишайник, на территории МНР найден лишь в районе Прихубсугулья, в Дархатской котловине, где обитает на известняковых скалах; известен также из окрестностей Красноярска и горных районов Северной Америки.
Основной ареал Lecidella alaiensis (Vain.) Hertel (Hertel, 1977) связан с горными районами Турции, Ирана, Средней Азии, Афганистана, Монголии и Северо-Западного Китая (Ганьсу, Цинхай); кроме того, этот вид также приводится для центральных и западных районов Северной Америки. На территории МНР встречается в юго-западной части Монгольского Алтая (Голубкова, 1981).
6. Евразиатский тип ареала присущ 40 видам, распространенным в горных районах Европы и Азии. Группа содержит виды рода Verrucaria - V. applanata Hepp, V. silicea Serv., V. floerkeana DT. et Sarnth., Dermatocarpon moulinsii (Mont.) Zahlbr., D. vellereum Zsch., Polyblastia plicata (Massal.) Lonnr., Amphoridium caesiopsilum (Anzi) Serv., Thelidium absconditum Rabenh., Porocyphus rehmicus (Massal.) Zahlbr., Moelleropsis nebulosa (Hoffm.) Gyeln., Aspicilia verruculosa Krempelh. и др.
Примером может служить ареал Verrucaria applanata Нерр, обитающего на камнях в горных реках и на скалах по берегам водоемов. Ареал этого лишайника охватывает горы Средней Европы - Шварцвальд, Гарц, Судеты, Татры, Карпаты (Zschacke, 1934; Servit, 1954; Окснер, 1956; Nowack, Tobolewski, 1975), Кавказ (Бархалов, 1975), Памир (Голубкова, 1973а, 1973б), Алтай (Седельникова, 1977б), Хангай и Монгольский Алтай.
7. Палеарктический тип ареала характерен для 3 видов - Polycoccum marmoratum (Krempelh.) D. Hawksw., Lecidea sulphurea (Hoffm.) Wahlenb., Caloplaca arenaria (Pers.) Mull. Arg.
Первый вид, обитающий на известняковых камнях и скалах, встречается рассеянно по всей Европе, в горах Фенноскандии, Великобритании, Франции, ФРГ, Австрии, Италии, Венгрии, в Крыму, Северной Африке (?) (Keissler, 1938; Макаревич, 1977). В Азии он известен в горах Таджикистана и в настоящее время найден на территории МНР, в Прихубсугулье.
Ареал Lecidea sulphurea (Hoffm.) Wahlenb. охватывает горы Европы, Крыма, Северной Африки (Алжир) (Макаревич, 1971а), Памир, Гималаи, Ганьсу (Hertel, 1977); в Монголии найден в горах Прихубсугулья.
8. Евразиатско-гренландский тип ареала свойствен одному виду Acarospora nitrophila Н. Magn., распространенному в горах Европы и Азии, а также в Гренландии.
9. Голарктический тип ареала присущ видам, встречающимся в горах Голарктики. Группа состоит из 23 видов - Stenocybe byssacea (Fr.) Koerb., Psorotichia schaereri (Massal.) Arnold, Thelidium olivaceum (Fr.) Koerb., Verrucaria laevata Ach., Acarospora impressula Th. Fr., A. oxytona (Ach.) Massal., Usnea perplexans Stirt., U. fulvoreagens (Ras.) Ras., Bryoria vrangiana (Gyeln.) Brodo et D. Hawksw., Dimelaena oreina (Ach.) Norm, и др.
Ареал Acarospora oxytona (Ach.) Massal. охватывает средиземноморские и горные районы Южной и Средней Европы, на севере включая Швецию, Канарские острова, горы Северной Африки, Кавказ, горы Ирана, Копетдаг, Прибайкалье, Якутскую АССР (среднее течение р. Индигирки), Хабаровский край (горы Шаман-Тиуль), Северную Монголию (Хангай, Хэнтэй, Монгольско-Даурский район, Средняя Халха), а также юго-западные районы Северной Америки и Центральную Америку (Голубкова, Шапиро, 1978; Голубкова, 1980б).
Б. Омнимонтанный субэлемент объединяет виды, встречающиеся, помимо Голарктики, в горах других флористических царств. Для видов этого субэлемента можно наметить следующие типы ареалов.
1. Голарктико-полинезийский тип ареала присущ одному виду - Polychidium muscicola (Sw.) S. Gray. Основная часть ареала этого вида связана с умеренной зоной Нового и Старого Света (Henssen, 1963). Этот слизистый лишайник встречается от Гренландии, о-ва Ян-Майен и арктической Европы до гор Западной и Средней Европы и далее, южнее - до Италии, Балканского полуострова, восточнее он известен в Эстонии, Карелии и Курской области, в Азии - на Алтае и в МНР, приводится также для Северной Африки и островов Океании (Окснер, 1956).
2. Мультирегиональный тип ареала свойствен 10 видам - Conotrema urceolatum Tuck., Parmelia delisei (Duby) Nyl., Peltigera degenii Gyeln., P. horizontalis (Huds.) Baumg., Pseudocyphellaria scrobiculata (Scop.) Blum, Collema subnigres- cens Degel. и др.
Примером может служить ареал Parmelia delisei (Duby) Nyl. (Esslinger, 1977), охватывающий Европу, от южной Скандинавии до южных районов и от Великобритании до европейской части СССР (Воронежская область, Крым), Северную Африку, Канарские острова, Кавказ, Азию (Сирия, Турция, Казахстан - СССР, Монголия), Австралию и Южную Америку. На территории МНР этот лишайник известен лишь из Хангая (Бязров и др., 1983), где встречается на камнях в петрофитной степи в восточной части этого горного района.
Ареал Collema subnigrescens Degel. приурочен к океаническим и горным районам Европы, включает Кавказ, тропические и умеренные районы Азии (Восточная Сибирь, Монголия, Китай, Япония, Непал, Индия, Шри Ланка, Малайзия, Индонезия, Филиппины), Тропическую и Северную Африку, Северную Америку (Канада, Флорида), Центральную Америку (Гондурас, Бермудские острова), где этот слизистый лишайник обитает главным образом в горах до высоты 3600 м над ур. м. (Degelius, 1954, 1974). В Монголии он был найден в горах Хэнтэя, в долине р. Онон и Тэрэлджийн-Гол на замшелых скалах (Голубкова, 1981).
Таким образом, среди видов монтанного элемента преобладают виды с евразиатскими ареалами (40 видов, или 45.5% от общего числа видов монтанных лишайников), что свидетельствует о существовании энергичных связей между лихенофлорами горных районов Европы и Азии. 23 вида (26.1%) фиксируют широкоголарктические флористические связи монтанных лишайников МНР, а 10 видов (11.3%), по-видимому, древние связи монтанной флоры МНР с флорами горных районов южного полушария, в частности палеотропиков. Только 7 видов (7.9%) имеют ареалы, ограниченные Азией; из них один вид отражает географические связи флоры Хангая с флорами горных районов Прибайкалья, один вид - с флорой Гималаев, 2 вида - географические связи флор Алтая и Хангая, с одной стороны, и Юньнаня, с другой. 3 вида (3.1%) имеют ареалы, охватывающие горные районы Азии и Северной Америки.
VIII. Субокеанический элемент. Охватывает виды, имеющие центры массовости в океанических и субокеанических регионах материков, а кроме того, нередко и во внутренних частях материков в подходящих местообитаниях (например, в горах по влажным долинам) (Трасс, 1970). Во флоре МНР субокеанический элемент объединяет 8 видов лишайников (1.06%). Для этих видов можно наметить следующие типы ареалов.
А. Голарктический субокеанический субэлемент.
1. Тихоокеанский тип ареала присущ одному виду - Cladonia kanewskii Oxn. (рис. 37). Основное распространение С. kanewskii Oxn. связано с тихоокеанскими побережьями Азии и Северной Америки (Ahti, 1973), но, кроме того, С. kanewskii Oxn. был найден в глубине Азиатского и Американского материков, на побережье оз. Байкал и на территории МНР, в южном Хэнтэе (Ahti, 1976; Голубкова, 1981; Ahti, Brodo, 1981). Обитание этого океанического вида в Монголии связано с особыми микроклиматическими условиями. Он был собран лишь однажды в верховье р. Тола, где произрастал в нижней части пойменной террасы, образуя здесь вместе с другими растениями довольно плотное и густое покрытие.
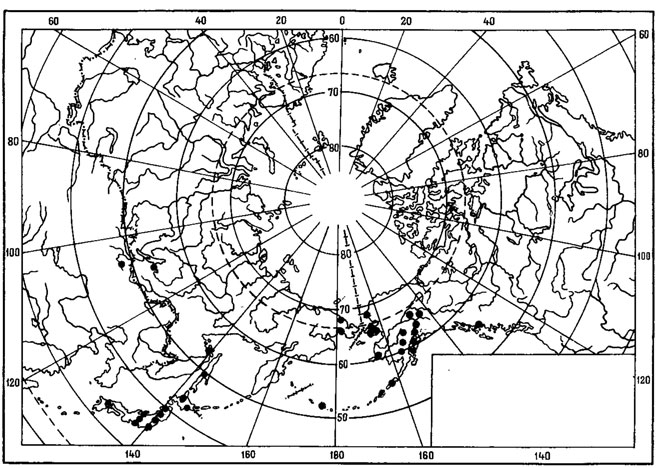
Рис. 37. Распространение на земном шаре Cladonia kanewskii Oxn. (По: Ahti, 1973; Ahti, Brodo, 1981, с дополнениями автора)
2. Евразиатский тип ареала характерен для одного вида - Xanthoria aureola (Ach.) Erichs., встречающегося в приатлантических, средиземноморских районах Европы; изолированные участки ареалов этого вида имеются во внутренних горных районах Азии.
3. Голарктический тип ареала свойствен 4 видам - Bryoria subcana (Nyl. ex Stiz.) Brodo et D. Hawksw., Sticta wrightii Tuck., Leptogium palmatpm (Huds.) Mont., Umbilicaria grisea Hoffm.
Одна часть ареала Bryoria subcana (Nyl. ex Stiz.) Brodo et D. Hawksw. связана с атлантическими и горными районами Европы, другая - с западным побережьем Северной Америки, от Аляски до Калифорнии (Brodo, Hawksworth, 1977); местонахождение на территории МНР в Монгольском Алтае (Schubert, Klement, 1971) - первое на материке Азии.
Основной ареал Sticta wrightii Tuck, совпадает с тихоокеанскими районами Азии, где этот вид встречается в Японии, на о-ве Тайвань, в Приморском крае, на Курильских островах. Далее на запад имеются изолированные местонахождения этого лишайника на Евразиатском континенте - в Прибайкалье, Красноярском крае, Монголии (Хангай), в Омской области, на Урале, в европейской части СССР (Вологодская область, Карелия), Средней Е вропе (горы ФРГ). Этот вид приводится также для Аляски (Krog, 1968), где был найден в районе восточной части Тихоокеанского побережья.
Основной ареал Leptogium palmatum (Huds.) Mont. (Degelius, 1935) связан с атлантическими и средиземноморскими районами Европы, где он встречается в юго-западных районах Норвегии, Швеции, в Дании, на Британских островах, во Франции, в Испании, Португалии, Италии, на Балканском полуострове; кроме того, этот лишайник распространен в некоторых горных районах Европы, в Швейцарии, Тироле, ФРГ, Польше (Силезия), а также в СССР - в Крыму. Он известен также на Канарских островах и в Северной Африке (Алжир). На территории Северной Америки его ареал приходится на западные районы этого материка, Аляску (Тихоокеанское побережье), Канаду, Калифорнию (Degelius, 1935; Krog, 1968). В Азии известно несколько изолированных местонахождений этого вида: в Западной Сибири, Монголии (Хангай), Китае (Шэньси), Японии, Северной Индии (Degelius, 1935; Окснер, 1956; Бязров и др., 1983). В Хангае этот вид был собран лишь однажды в верховьях реки Хух-Сум, на высоте 1820 м, в лиственничном лесу на мшистых скалах (Бязров и др., 1983).
Б. Омнисубокеанический субэлемент включает 2 вида, встречающихся в океанических областях за пределами Голарктики, - Hypogymnia enteromorpha (Ach.) Nyl. и Pannaria conoplea (Ach.) Bory.
Ареал Hypogymnia enteromorpha (Ach.) Nyl. связан с бассейном Тихого океана, где этот вид растет на Дальнем Востоке (Амурская и Магаданская области, Хабаровский край, о-в Сахалин), в Китае, на Тихоокеанском побережье Аляски и в Калифорнии, а также в Австралии, Новой Зеландии и на о-ве Тасмания. На территории Монголии встречается в Хангае (Бязров и др., 1983).
Как отмечал Шауер (Schauer, 1965), Pannaria conoplea (Ach.) Bory (рис. 38) относится к числу лишайников - индикаторов возрастания океаничности климата. Йоргенсен (J∅rgensen, 1978) указывал, что этот вид требует для своего обитания довольно высокой влажности, и в континентальных районах он обычно растет вблизи водопадов, в узких горных ущельях. P. conoplea (Ach.) Bory широко распространен в атлантической, Средней и Южной Европе (Degelius, 1935; Schauer, 1965; J∅rgensen, 1978), от Северной Норвегии до Южной Португалии, и от Ирландии до европейской части СССР. Далее по направлению к востоку его встречаемость постепенно падает: он растет в Крыму, на Кавказе, Урале и далее после значительной дизъюнкции - на территории Монголии (J∅rgensen, 1978; Голубкова, 1981) и затем в Японии. В Северной Америке обычен на равнинах северо-восточных районов США и прилегающих районах Канады, на западе - на Аляске и в Скалистых горах, спускается к югу до Мексики и Южной Америки (Венесуэла) (J∅rgensen, 1978), известен также в горных районах Восточной Африки и некоторых других районах палеотропиков. На территории МНР обитает на замшелых скалах в лиственничных и березовых лесах, горных степях Северо-Западной Монголии (хр. Тургэн-Ула), в центральном Хангае (окрестности оз. Цаган-Нур), в Хэнтэе и долине р. Улдза.
Рис. 38. Распространение на территории Монголии Pannaria conoplea (Ach.) Bory. (Ориг.)
IX. Мультизональный элемент. Охватывает виды лишайников, широко распространенные во многих растительно-климатических зонах Голарктики, а также в других флористических царствах. Он насчитывает в лихенофлоре МНР 86 видов (11.4%).
А. Голарктический мультизональный субэлемент. Видам этого субэлемента свойственны следующие типы ареалов.
1. Евразиатский тип ареала присущ 2 видам - Rhizocarpon reductum Th. Fr. и Aspicilia cinerea (L.) Koerb,. обитающим в ряде растительно-климатических зон Европы и Азии.
2. Голарктический тип ареала характерен для 40 видов, распространенных во многих растительно-климатических зонах Голарктики. К этой группе относятся виды родов Lecanora - L.crenulata (Dicks.) Hook., L. hagenii Ach., L. campestris (Schaer.) Hue, Aspicilia - A. contorta (Hoffm.) Krempelh.; Acarospora - A. veronensis Massal., A. fuscata (Rohl.) Arnold, A. glaucocarpa (Wahlenb.) Koerb.; Caloplaca - C. decjpens (Arnold) Blomb. et Forss., C. variabilis (Pers.) Mull. Arg.; Verrucaria muralis Ach., Endopyrenium rufescens (Ach.) Koerb., Endocarpon pusillum Hedw., Dermatocarpon miniatum (L.) Marin, Sarcogyne clavus (Ram.) Krempelh., Rinodina sophodes (Ach.) Massal., R. bischoffii (Hepp) Massal., R. badiella (Nyl.) Th. Fr., Bacidia muscorum (Ach.) Mudd и др.
Многие из этих лишайников в своем распространении тяготеют к южным районам Голарктики, например Acarospora glaucocarpa (Wahlenb.) Koerb., Caloplaca variabilis (Pers.) Mull Arg., Lecanora garovaglii (Koerb.) Zahlbr., другие же, наоборот, к северным, например Bacidia muscorum (Ach.) Mudd, который особенно широко распространен в таежной зоне Голарктики.
Б. Омнимультизональный субэлемент объединяет виды, которым свойствен один тип ареала - мультирегиональный.
1. Мультирегиональный тип ареала характерен для 44 видов, обитающих во многих растительно-климатических зонах земного шара. Эта группа включает виды родов Peltigera - P. canina (L.) Willd., P. rufescens (Weis.) Humb.; Rhizocarpon - R. viridiatrum (Wulf.) Koerb., R. disporum (Naeg.) Mull. Arg., R. montagnei (Flot.) Koerb.; Lecanora - L. atra (Huds.) Ach., L. rupicola (L.) Zahlbr., L. bicincta Ram. (табл. 1, 3); Candelariella - C. aurella (Hoffm.) Zahlbr., C. vitellina (Ehrh.) Miill.; Caloplaca - C. cerina (Ehrh.) Th. Fr., C. flavovirescens (Wulf.) DT. et Sarnth., C. saxicola (Hoffm.) Nordin; Physcia - Ph. caesia (Hoffm.) Fiirnr., Ph. phaea (Tuck.) Thorns., Ph. dubia (Hoffm.) Lett.; Parmelia sulcata Tayl., Polysporina simplex (Dav.) Vezda, Sarcogyne regularis Koerb. em. Oxn. и др.
Большинство лишайников мультизонального элемента - это растения с широкой экологической амплитудой, что позволяет им широко расселяться от арктических районов до областей тепло- умеренной зоны, в аридных и даже в тропических районах земного шара. Среди них имеется немало эвритопных видов, способных поселяться на самых разных субстратах и в различных местообитаниях. Например, Candelariella aurella (Hoffm.) Zahlbr. - лишайник, растущий на известняках, карбонатной почве, обработанной и гнилой древесине, коре деревьев, цементе, черепице и других субстратах. Он широко распространен по всей Голарктике: вся Европа, от Арктики и Гренландии до Кавказа и Южного Урала, Средняя Азия, Восточная Сибирь, Монголия, Северная Африка, Северная и Центральная Америка.
Lecanora hagenii Ach. - лишайник, чрезвычайно широко распространенный в Голарктике, обычно встречается в нитротических условиях, где поселяется на обработанной древесине, коре деревьев, костях, листовом железе и других субстратах.
Иногда мультизональные виды являются синтопными лишайниками, строго приуроченными к какому-либо субстрату или к определенным местообитаниям. Среди видов этого элемента выделяется довольно обширная группа лишайников, обитание которых всегда связано с карбонатными породами. В различных растительно-климатических зонах на этих породах можно встретить очень сходную по видовому составу флору кальцефильных лишайников. К числу таких видов относятся Acarospora glaucocarpa (Wahlenb.) Koerb., Sarcogyne regularis Koerb. em. Oxn., Polysporina simplex (Dav.) Vezda, Aspicilia calcarea (L.) Mudd.
Строгая приуроченность к определенному субстрату является причиной того, что некоторые мультизональные лишайники редки на территории МНР. Примером может служить Acarospora glaucocarpa (Wahlenb.) Koerb., которая встречается лишь на известняковых скалах в районе Прихубсугулья. Большинство же мультизональных лишайников широко распространено в Монголии.
На основании проведенного анализа можно сделать вывод о том, что флора лишайников МНР представляет собой сложный комплекс различных географических групп лишайников. Для лихенофлоры МНР мы выделяем 9 географических элементов: арктовысокогорный, гипоарктомонтанный, неморальный, бореальный, аридный, высокогорный, монтанный, субокеанический, и мультизональный. Распределение видов флоры лишайников МНР по географическим элементам приводится ниже.
| Число видов | % от общего числа видов флоры | |
| Аридный | 166 | 22.0 |
| Бореальный | 145 | 19.2 |
| Арктовысокогорный | 108 | 14.3 |
| Монтанный | 88 | 11.6 |
| Мультизональный | 86 | 11.4 |
| Гипоарктомонтанный | 59 | 7.8 |
| Неморальный | 45 | 5.9 |
| Высокогорный | 19 | 2.8 |
| Субокеанический | 8 | 1.06 |
| Невыясненные ареалы | 30 | 3.9 |
На ход исторического развития флоры лишайников Монголии как в прошлое историческое время, так и в настоящее большое влияние оказала прогрессирующая аридизация резко континентального климата в сочетании со значительной физико-географической изоляцией ее территории. Это способствовало широкому расселению в пределах Монголии ксерофитных видов лишайников. В лихенофлоре Монголии ведущее положение среди географических групп лишайников занимает аридный элемент, охватывающий 166 видов (22%). Именно аридные лишайники в значительной степени определяют специфические черты лихенофлоры МНР. Ареалогический анализ аридных лишайников показывает, что среди них во флоре Монголии преобладают виды с ареалами, ограниченными территорией Азии (100 видов, или 60.1% от общего числа видов аридного элемента). Более половины этих видов (53 вида, или 31.8%) характеризуется ареалами, ограниченными территориями различных провинций Центральной Азии, при этом среди них преобладают монгольские эндемичные виды (21 вид, или 12.6% от числа видов аридного элемента). Все это дает основание предполагать значительную роль автохтонного элемента в формировании аридной лихенофлоры Монголии. 30 видов (18.1% от числа видов аридного элемента) имеют ареалы, охватывающие территорию Ирано-Туранской области Древнесредиземноморского подцарства Голарктики, тем самым подтверждая флористические связи Монголии с флорой Средней Азии, Казахстана и Передней Азии. 18 видов (10.8% от числа видов аридного элемента) характеризуются ареалами, связанными со всей территорией Древнесредиземноморского подцарства Голарктики, фиксируя географические связи флоры Монголии с флорой не только Ирано-Туранской, но и Средиземноморской областей. Общее число видов аридных лишайников Монголии, ареалы которых ограничены территорией Древнего Средиземья, достигает 100 (60.1% от общего числа видов аридного элемента). Уже только это позволяет предположить древнесредиземноморскую основу в развитии аридного элемента лихенофлоры Монголии. Географические связи изучаемой лихенофлоры с флорой пустынных районов юго-запада Северной Америки, Сонорского подцарства Голарктики, фиксирует 8.3% видов. Беднее представлены в аридном элементе связи с флорами понтических районов Европы (3 вида, или 2%). Флористические связи Монголии и Сибири подтверждают 17 видов (10.2%). Древние связи аридной флоры Монголии с флорами аридных районов Южной Африки, Австралии, Южной Америки проявляют 20 видов (12%).
Второе место среди географических групп лишайников Монголии принадлежит бореальному элементу, охватывающему 19.2% состава лихенофлоры Монголии (145 видов). Бореальные виды лишайников широко представлены в лесных ценозах Монголии, хотя некоторые из них часто встречаются в горных степях, поднимаясь до высокогорного пояса, а также в равнинных степях Монголии, особенно ее восточных районов. Как можно видеть, в составе бореального элемента преобладают виды с циркумполярным распространением в Голарктике (46.8%). Широкими мультирегиональными ареалами характеризуются 40.8% видов бореального элемента. Только 8.2% видов имеет более узкие евразиатские ареалы, а 4.1% - азиатские. Это указывает на слабую специфичность состава бореального элемента лихенофлоры Монголии по сравнению с бореальными флорами других районов Голарктики. Как подчеркивал Ахти (Ahti, 1977), циркумполярному расселению лишайников в зоне хвойных лесов способствуют относительное однообразие экологических условий, слабое влияние флорогенетических изоляций и замедленное протекание эволюционных процессов компонентов флоры. Как было показано, бореальной элемент монгольской лихенофлоры представляет группу видов, неоднородную в историческом и генетическом отношении. Некоторые из них, видимо, автохтонно бореального генезиса и относятся к числу древних бореальных типов (Parmeliopsis aleurites (Ach.) Nyl., Buellia schaereri DNot., B. zahlbruckneri Steiner, Cornicularia' aculeata (Schreb.) Ach., Parmelia olivacea (Scop.) S. Gray и др.). Другие, возможно, молодые виды, сформировавшиеся лишь в начале четвертичного периода (Rinodina exiguella Н. Magn., R. sibirica H. Magn., Evernia mesomorpha Nyl. и др.). В бореальном элементе лишайников Монголии, по-видимому, богато представлены бореализированные дериваты тургайской флоры - Evernia mesomorpha Nyl., Е. divaricata (L.) Ach. виды рода Bryoria. В формировании бореальной флоры лишайников Монголии, очевидно, значительна роль восточноазиатского флористического центра, с которым связан генезис ряда систематических групп лишайников, играющих заметную фитоценотическую роль в лесах Монголии и других хвойнолесных районов Голарктики, например видов родов Hypogymnia, Cetraria, Evernia и др. Среди бореального комплекса видов прослеживается также тропогенный элемент. К числу бореализированных тропогенных лишайников, видимо, можно отнести ряд представителей родов Cladonia, Peltigera, Usnea, Lecanora.
Как уже указывалось, общей и характерной физико-географической особенностью МНР является ее высокое положение над уровнем океана. Крупные горные поднятия в сочетании с обширными впадинами и возвышенными равнинами определяют в основных чертах современный рельеф Монголии. Поэтому неудивительно, что в географическом спектре флоры лишайников МНР широко представлены такие географические группы видов, как арктовысокогорный, монтанный и гипоарктомонтанный элементы, в значительной степени определяющие специфику флоры лишайников этого региона. Горные виды лишайников занимают значительное место в лихенофлоре Монголии (более 36% ее состава), что дает основание предполагать видную роль горных флористических центров в формировании флоры лишайников изучаемого района.
Третье место в числе географических групп видов в лихенофлоре Монголии принадлежит арктовысокогорному элементу, насчитывающему 108 видов (14.3%). Заметная роль арктовысоко- горного элемента в составе флоры лишайников Монголии, охватывающего почти 1/7 часть ее состава, привлекает внимание, особенно если учесть, что высокогорные пустоши и горные тундры представлены на территории этой страны весьма скудно - ими заняты 3% от общей площади Монголии (Юнатов, 1954). Большинство арктовысокогорных лишайников ограничено в своем распространении, на территории МНР высокогорным поясом Монгольского Алтая, Хэнтэя, Хангая и горных районов Прихубсугулья, где они играют заметную фитоценотическую роль в составе кобрезиевников, высокогорных степей, горных тундр. Характерно также широкое участие арктовысокогорных видов лишайников в составе лесных и степных сообществ Монголии, что может найти объяснение как в современных физико-географических условиях Монголии, так и в истории становления ее растительного покрова. Широкими голарктическими ареалами характеризуется 51.4% видов арктовысокогорного элемента; ареалы, простирающиеся за пределы Голарктики, имеет 30.5% видов, евразиатские - 12.1, азиатско-американские - 2.8% видов. Это свидетельствует о широких флористических связях между горными районами Монголии и горными системами и полярными районами Сибири, а также между другими районами Голарктики и за ее пределами. На основании этого можно говорить о достаточно крупных миграциях холодостойких форм, видимо, в плейстоцене и высказать предположение, что формирование арктовысокогорного элемента лихенофлоры Монголии происходило в основном аллохтонно, за счет иммиграций видов с других территорий. В генетическом отношении арктовысокогорный элемент представляет собой гетерогенную группу видов. Многие из них, по-видимому, голарктического горного генезиса (например, род Solorina), другие- арктического происхождения, генезис третьих связан с высокогорными и полярными районами южного полушария (например, род Neuropogon). Значительное место в составе флоры Монголии занимают арктовы- сокогорные виды лишайников азиатского горного генезиса (например, виды родов Asahinea, Cetraria и др.). Прослеживаются генетические связи некоторых арктовысокогорных форм с лесными флорами тургайского типа. Дифференциация и формирование некоторых из холодостойких ореофитных форм (например, Evernia perfragilis Llano), возможно, происходили в результате деградации мезофильных лесных элементов в криоксеротических условиях перигляциальных пространств Северной Монголии в ледниковое время. Сухой и холодный климат плейстоцена, видимо, способствовал не только развитию в верхних поясах гор высокогорной растительности, в составе которой значительное участие приняли арктовысокогорные виды лишайников, но и широкому расселению арктовысокогорных видов лишайников в средне- и нижнегорных поясах, а также их проникновению в равнинные степи Монголии. Современные природные условия Монголии способствуют консервации гляциальных и перигляциальных реликтовых видов лишайников в лесных и степных ценозах Монголии.
В составе монтанного элемента лихенофлоры Монголии преобладают виды с евразиатскими ареалами (45.5% от общего числа видов монтанных лишайников), что является показателем энергичных взаимосвязей между флорами лишайников горных районов Европы и Азии. Голарктическими ареалами характеризуется 26.1% видов, а мультирегиональными - 11.3% видов, фиксируя, видимо, древние географические связи монтанной флоры Монголии с флорами горных районов не только Голарктики, но и других флористических областей земного шара, в частности палеотропиков. Ареалы, ограниченные Азией, имеет 7.9% видов монтанного элемента, в чем проявляются флористические связи горных районов Монголии с флорами Южной Сибири, Гималаев, Юньнаня. Ареалами с тихоокеанским разъединением харктеризуется 3.1% видов монтанных лишайников, подтверждая географические связи горных флор Азии и Северной Америки.
Среди гипоарктомонтанного элемента, охватывающего 7.8% состава флоры лишайников Монголии, 5% видов также имеет американо-азиатские ареалы. Однако большинство видов этой географической группы лишайников (71.1%) с широко голарктическими ареалами, а остальная часть видов имеет ареалы, простирающиеся за пределы Голарктики. Это, видимо, является показателем значительной роли миграционных процессов в формировании этой географической группы лишайников на территории Монголии. Характерная особенность растительного покрова Монголии, как и в случае арктовысокогорных лишайников, - широкое участие гипоарктомонтанных видов лишайников в составе лесных, а также степных растительных сообществ. При этом некоторые из них, например Hypogymnia bitteri (Lynge) Ahti, Cladonia amaurocraea (Flk.) Schaer., Physconia mustigena (Ach.) Poelt, Aspicilia cupreoatra (Ach.) Arnold, играют определенную фитоценотическую роль в составе лесных и горностепных ценозов Монголии.
Роль высокогорного элемента в составе флоры лишайников Монголии, как можно было видеть, мала. Эта географическая группа лишайников объединяет всего 2.2% видов от общего состава лихенофлоры. Однако 1/3 часть видов высокогорного элемента характеризуется ареалами, ограниченными высокогорьями Азии. Один из них - Evernia terrestris (Tomin) Golubk. - имеет ареал, связанный с Хангайско-Алтайской горной страной, другие виды фиксируют флористические связи высокогорных районов Монголии с Тибетом, Гималаями, Юньнаньским нагорьем, Тянь-Шанем, Памиро-Алаем, Кавказом и горами Ирана.
В составе флоры лишайников МНР две географические группы занимают несколько обособленное положение, выпадая из общей специфики географического спектра лихенофлоры. Речь идет о неморальном и субокеаническом элементах. Неморальный элемент охватывает 5.9% состава лихенофлоры этого района. Среди представителей этой географической группы преобладают виды, характеризующиеся широкими дизъюнктивными ареалами как в Голарктике (31.1% от числа видов неморального элемента), так и за ее пределами (46.6%). Виды с более узкими евразиатскими ареалами составляют 11.1%, и такое же относительное число видов - с восточноазиатскими. В генетическом отношении неморальный элемент лихенофлоры Монголии представляет собой гетерогенную группу. В нем можно различить автохтонный тургайский геоэлемент, представленный, например, видами родов Cetrelia, Cetraria, Evernia и др., и древнесредиземноморский (Anaptychia ciliaris (L.) Koerb).
Заметную роль в составе неморального элемента в лихенофлоре Монголии играют тропогенные формы. В их составе, с одной стороны, тропические формы, основная часть ареалов которых связана с тропическими и субтропическими областями земного шара. К их числу относятся, например, Pseudevernia cirrhata (Fr.) Schub. et Klem., Lobaria retigera (Bory) Trevis., L. isidiosa (Miill. Arg.) Vain, и др. С другой стороны, среди них встречаются тропогенные формы, ассимилированные и переработанные в тургайской флоре, например виды рода Arthopyrenia.
В формировании мезофильной неморальной флоры лишайников Монголии, как было показано, особенно велика роль восточно-азиатского флористического центра. Значительно меньшая роль принадлежит западноевропейскому центру. Иммиграция некото рых форм, возможно, могла происходить из сибирских рефугиумов неморальной флоры. Не исключена их иммиграция из горных районов Средней Азии. Современное географическое распространение некоторых видов неморальных лишайников и их генетические связи позволяют высказать предположение, что их местообитания на территории МНР, видимо, реликтовые.
Субокеанический элемент представлен 8 видами (1.06%). Тем не менее находки видов этого элемента в составе флоры страны, характеризующейся крайне континентальным климатом, безусловно представляют интерес. Среди субокеанических лишайников можно различить группу видов, например Cladonia kanewskii Oxn., Sicta wrightii Tuck., Hypogymnia enteromorpha (Ach.) Nyl., основное распространение которых в основном связано с бассейном Тихого океана. Ареалы других видов, например Leptogium palmatum (Huds.) Mont., Pannaria conoplea (Ach.) Bory, приурочены главным образом к атлантическим и средиземноморским районам Европы. Некоторые из этих видов на территории МНР имеют реликтовые ареалы.
Ареалогический анализ лишайников Монголии выявляет, что среди них наибольшее число видов, 263 вида, или 34.8% от общего числа видов лихенофлоры, характеризуется широкими голарктическими ареалами. Среди видов голарктического типа распространения выделяются группы лесных бореальных лишайников (69 видов) и арктовысокогорных лишайников (55 видов). Довольно многочисленны также группы гипоарктомонтанных (41 вид) и мультизональных (40), а также монтанных лишайников (23 вида).
Обширные ареалы, охватывающие не только Голарктику, но и другие флористические царства, имеют 207 видов (27.4% от общего числа видов лихенофлоры). В их числе преимущественно бореальные (59 видов), мультизональные (44) и арктовысокогорные (33 вида) лишайники: Среди группы видов мультирегионального типа распространения заметное место также занимают неморальные (21 вид) и, аридные (20 видов) лишайники.
Ареалы, простирающиеся в пределах Европы и Азии, имеет 80 видов. Причем значительную часть группы евразиатских лишайников составляют монтанные виды (40 видов). Более слабо в ней представлены арктовысокогорные (13) и бореальные (12 видов) лишайники. В группе лишайников с азиатско-американскими ареалами только 11 видов. Среди них арктовысокогорные (4 вида), гипоарктомонтанные (3), аридные (3) и субокеанические (1 вид) лишайники.
Видов с ареалами, расположенными в пределах Азии, 127 (16.8% от общего числа видов лихенофлоры). Большую часть группы азиатских видов составляют аридные лишайники (100 видов). Остальные виды азиатского распространения распределяются между монтанными (7 видов), бореальными (6 видов), высокогорными (6 видов), неморальными (5) и арктовысокогорными (3 вида) лишайниками. Из азиатских видов - 53 (7.02% от общего числа видов лихенофлоры) являются центральноазиатскими эндемиками. Из них 21 вид (2.7% от общего числа видов) - монгольские эндемичные виды.
Территория МНР является своеобразным биогеографическим рубежом. В лесах Монголии, с одной стороны, проходят южные границы ареалов многих бореальных и бореально-монтанных видов лишайников. С другой стороны, на территории Монголии находятся северные границы многих пустынно-степных центрально-азиатских и ирано-туранских видов лишайников. Многие ирано-туранские виды и виды с обширными древнесредиземноморскими ареалами имеют на территории МНР восточные границы своего распространения. Но в свою очередь с востока на территорию Монголии проникают некоторые восточноазиатские виды лишайников, которые имеют в Монголии свои наиболее западные местонахождения.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'