Бентосные водоросли (Ю. Е. Петров)
К числу бентосных (донных) водорослей относятся водоросли, приспособленные к существованию в прикрепленном состоянии на дне водоемов и на разнообразных предметах, живых и мертвых организмах, находящихся в воде.
В зависимости от места произрастания среди бентосных водорослей различаются: 1) эпилиты, которые растут на поверхности твердого грунта (скалы, камни и т. д.); 2) эпипелиты, населяющие поверхность рыхлых грунтов (песок, ил); 3) эпифиты, живущие на поверхности других водорослей; 4) эндолиты, или сверлящие водоросли, внедряющиеся в известковый субстрат (скалы, раковины моллюсков, панцири ракообразных); 5) эндофиты, которые поселяются в слоевищах других водорослей, но, в отличие от паразитических видов, содержат нормальные хлоропласты в клетках; 6) паразиты, живущие в слоевищах других водорослей, хлоропласты в клетках не выражены.
Иногда водоросли, растущие на предметах, введенных в воду человеком (суда, плоты, буи), относят к перифитону. Выделение этой группы обосновывают тем, что входящие в ее состав организмы (водоросли и животные) живут на предметах, большей частью находящихся в движении или обтекаемых водой. Кроме того, эти организмы удалены от дна и, следовательно, находятся в условиях иного светового и температурного режимов, в других условиях поступления биогенных веществ, источником которых служат донные отложения. Иногда выделение перифитона обосновывают еще и практическими соображениями: это обрастания, которые могут причинять практический ущерб - уменьшать скорость судов, засорять водозаборные отверстия и трубопроводы.
Между эпилитами, эпипелитами и эпифитами часто нет резкой грани, особенно это относится к микроскопическим бентосным водорослям. Правда, существуют виды, которые живут только на других водорослях, и притом только на определенном виде. Например, полисифония шерстистая (Polysiphonia lanosa) растет исключительно на аскофиллуме узловатом (Ascophyllum nodosum).
Обычно зачатки водорослей (споры, гаметы, зиготы) с водой заносятся на самые разнообразные субстраты. Многие эпилиты произрастают в качестве эпифитов. Возможность роста их до зрелого состояния определяется размерами их слоевищ и слоевища хозяина. Когда в качестве эпифита растет крупная водоросль, слоевище хозяина рвется под действием волн и течения.
Существует определенная связь между размерами водорослей, размером частиц грунта, к которым они прикрепляются, и интенсивностью движения воды. На песке и иле способно расти относительно небольшое число макроскопических водорослей. В условиях движения воды в морях к этому хорошо приспособлены, например, некоторые виды рода халимеда: их тонкие ризоиды цементируют песок при основании слоевища в плотную массу. Как правило, чем крупнее взрослое слоевище водоросли и чем сильнее движение воды, тем больших размеров (большей массы) должны быть камни, на которых они растут. Иначе волны или течения сносят их на большие глубины или выбрасывают на берег.
Зачатки водорослей способны закрепляться на частицах грунта любого размера. По мере их роста происходит отсев и гибель тех слоевищ, которые растут на недостаточно крупных камнях. При росте водорослей на рыхлых грунтах, например на песке, существует опасность перемещения грунта придонными течениями, в результате чего растения или заносятся грунтом, или перетираются им. Но даже в таких местах песок не остается совершенно лишенным водорослей. В неровностях песчинок живут микроскопические одноклеточные и колониальные виды.
Обычно считается, что для роста водорослей особенно важен свет. Однако, выйдя на берег реки, озера или моря, можно заметить, что водоросли растут далеко не всюду, где имеется достаточно прозрачная вода. Часто бывает наоборот, бентосные водоросли неплохо растут в местах с мутной водой, но зато находящейся в движении.
Как правило, места с интенсивным движением воды отличаются пышным развитием бентосных водорослей. В морях это проливы с течениями, районы мысов и другие прибрежные участки, где наблюдается постоянный прибой. В реках - это камни на перекатах, где вода течет с большой скоростью.
Движение воды оказывает многообразное действие на различные стороны жизни бентосных водорослей. Во-первых, усиливается поглощение водорослями биогенных (питательных) веществ за счет того, что в местах с постоянным течением или волнами вода непосредственно около водорослей непрерывно обновляется с большой скоростью (от нескольких сантиметров до нескольких метров в секунду).
Одновременно фотосинтез усиливается в два раза, а дыхание - на 30-50%. Иными словами, бентосные водоросли, растущие в условиях движения воды, получают преимущества по сравнению с водорослями в тихих местах: один и тот же уровень фотосинтеза может быть достигнут у них при меньшей освещенности, что в итоге способствует росту более крупных слоевищ с большим содержанием белка и углеводов. Движение воды предотвращает оседание на скалы и камни илистых частиц, которые мешают закреплению зачатков водорослей. Кроме того, движение воды благоприятствует росту бентосных водорослей тем, что с поверхности грунта удаляются животные, питающиеся водорослями. Наконец, хотя при сильном течении или сильном прибое происходит повреждение слоевищ водорослей или отрыв их от грунта, движение воды все же не препятствует поселению микроскопических водорослей и микроскопических стадий крупных водорослей. Дело в том, что из-за трения о поверхность грунта скорость движения воды по мере приближения к нему уменьшается и на самой поверхности грунта существует так называемый пограничный слой воды, где течение отсутствует. Толщина этого слоя прямо пропорциональна протяженности обтекаемого предмета (например, камня или выступа скалы) и обратно пропорциональна скорости течения. Обычно толщина пограничного слоя бывает не меньше 10-100 мкм, что соответствует размерам спор, гамет и зигот большинства бентосных водорослей.
В пресных водах движение воды имеет ощутимое значение в реках, ручьях и горных потоках. В этих водоемах выделяется группа бентосных реофильных организмов, предпочитающих места с постоянным течением. В озерах не бывает сильных течений, и здесь для произрастания бентосных водорослей приобретают значение волны.
В морях волны также оказывают значительное влияние на жизнь бентосных водорослей. По мере приближения к берегу глубина уменьшается, это приводит к возрастанию высоты волн, но до определенного предела. Когда глубина становится равной примерно 1,3 высоты волн, происходит их разрушение. Существуют три типа разрушения волн: 1) медленное разрушение или рассыпание (скользящий бурун) на малых уклонах дна, где на расстоянии 100 м от берега глубина составляет 1-5 м; 2) опрокидывание гребня (ныряющий бурун) на больших уклонах (глубина на расстоянии 100 м колеблется от 5 до 10 м); 3) опрокидывание всего переднего склона волны при таких уклонах, где глубина на расстоянии 100 м от берега превышает 10 м. Больше всего благоприятствует росту водорослей разрушение волн по первому типу, которое сопровождается появлением длинных прибойных потоков. В таких условиях, так же как в местах с постоянными течениями, водоросли достигают наиболее крупных размеров. Разрушение волн по второму и особенно по третьему типу сопровождается сильными ударами больших масс воды о дно, что приводит к повреждению слоевищ водорослей, и в таких местах они становятся короче.
Действие волн может сказываться на вертикальном распределении водорослей. У открытых побережий морей ... океанов, к которым подходят крупные волны с больших водных пространств, на глубине от 0 до 15 м, в зависимости от рельефа дна бывают зоны, лишенные макроскопических (крупных) водорослей. Исключение составляют корковые известковые красные водоросли, способные противостоять сильным ударам волн. Заросли прочих водорослей располагаются или на малых глубинах, куда доходят ослабленные волны, образующиеся после разрушения более крупных, или на большой глубине, если там достаточно света, где не сказывается разрушительное действие крупных волн.
Интенсивному развитию бентосных водорослей способствует также умеренное (не до загрязнения) содержание в воде биогенных веществ. В пресных водах это характерно для неглубоких прудов и берегов озер, заводей по берегам рек; в морях - для мелких заливов. Источниками биогенных веществ в воде служат береговые стоки и донные отложения, особенно велика роль последних как аккумуляторов органических остатков. В донных отложениях в результате жизнедеятельности бактерий и грибов происходит минерализация органических остатков; сложные органические вещества переходят в простые неорганические соединения, доступные для использования фотосинтезирующими растениями. Наиболее благоприятны для развития бентосных водорослей места с отложением органических остатков на небольших глубинах с достаточным освещением, твердыми грунтами для поселения водорослей и слабым движением воды. Здесь продукты минерализации быстро вовлекаются в процессы фотосинтеза, а слабое движение воды содействует их переносу от донных отложений в заросли водорослей и способствует обогащению придонной воды кислородом, что препятствует появлению в ней сероводорода, губительного для живых организмов.
Влияние изменений в содержании биогенных веществ в воде на рост макроскопических бентосных водорослей особенно заметно в тропиче-ских морях. В экваториальном поясе воды океанов бедны биогенными элементами, соответственно по открытым берегам островов такие водоросли встречаются нечасто и имеют мелкие слоевища (высотой до 10 см). В лагунах и заливах при наличии илистого песка на малых глубинах количество водорослей резко возрастает, а те виды, что встречались в открытых местах, увеличиваются в размерах в 2-3 раза. В тропическом поясе встречаются большие пространства прибрежных вод, например западное побережье Индии, отличающиеся повышенным содержанием биогенных веществ из-за апвеллинга - подъема глубинных вод. В таких районах тропических морей заросли крупных водорослей наблюдаются и на открытых берегах.
В местах, где отсутствует движение воды и она не обогащена биогенными веществами, бентосные водоросли растут плохо. Это бывает на скалистых берегах бухт с большим уклоном дна и значительными глубинами в центре, из-за чего вещества от дойных отложений не выносятся в верхние горизонты. Макроскопические морские водоросли могут отсутствовать в открытых местах на отвесных стенах скал, когда глубина около них превышает полдлины волны. При таких условиях волны отражаются без разрушения, а значит, отсутствует забурунивание и интенсивное движение воды у поверхности скал. Зато на тех же скалах макроскопические водоросли образуют сплошной покров на наклонной поверхности волноприбойной ниши, выбитой волнами у уреза воды.
Кроме света, движения воды и содержания биогенных веществ, произрастание бентосных водорослей зависит от присутствия растительноядных водных животных (морских ежей, брюхоногих моллюсков, ракообразных, рыб). Это особенно заметно по зарослям ламинариевых водорослей, отличающихся среди морских водорослей крупными размерами. Нижняя граница их произрастания нередко определяется присутствием морских ежей. Если уничтожать ежей и следить, чтобы они не наползали снова, то ламинариевые начинают расти глубже. У атлантических берегов Канады установлена зависимость между выловом омаров и величиной площади под зарослями ламинариевых водорослей. Когда вылавливали много омаров, заросли водорослей сокращались. Достаточно было уменьшить промысел омаров - площадь под зарослями увеличивалась. Оказывается, омары питаются морскими ежами и вылов омаров способствовал увеличению их численности.
Замечено, что в тропических морях в некоторых местах рыбы полностью выедают виды зеленых, бурых и красных водорослей с мягкими слоевищами, в результате остаются несъедобные для рыб и других животных сине-зеленые водоросли и представители других групп водорослей, имеющих жесткие слоевища. Брюхоногие моллюски, ползая по дну, поедают микроскопические водоросли и мелкие проростки макроскопических видов.
На произрастание определенных видов водорослей влияют не только свет, грунт, движение воды, ее химический состав, растительноядные животные, но и присутствие других видов водорослей. Лучше всего это заметно на морских видах фукусов. Обычно они растут в зоне, обнажаемой во время отливов, глубже грунт занят другими водорослями. В Арктике, где число видов водорослей уменьшается, Fucus distichus растет и глубже. То же самое происходит с Fucus vesiculosus и в сильно опресненном Балтийском море.
В настоящее время в связи с усиливающимся загрязнением прибрежных вод наблюдается исчезновение крупных морских водорослей в некоторых бухтах. Но если из чистых мест туда перенести и закрепить крупные слоевища водорослей, то они будут расти. Как установлено, в загрязненных местах усиленно развиваются микроскопические бентосные водоросли, которые быстро обрастают проростки более крупных водорослей и губят их. Немаловажную роль при этом играет то обстоятельство, что зачатки крупных водорослей по размерам часто не превосходят даже одноклеточных водорослей.
Влияние бентосных водорослей друг на друга начинается с момента оседания и прорастания спор. Например, во время эксперимента зооспоры ламинариевых не прорастали, когда в сосуде находились кусочки слоевища аскофиллума.
В холодных морях на бентосные водоросли оказывает влияние лед. В зависимости от его толщины, движения и торошения заросли водорослей могут быть уничтожены (стерты) до глубины в несколько метров. Поэтому, например, в Арктике многолетние водоросли (фукус, ламинария) легче всего найти у берега среди валунов и выступов скал.
Многообразное влияние на жизнь бентосных водорослей оказывает температура. Наряду с другими факторами она определяет скорость роста, темп и направление развития бентосных водорослей, момент закладки у них органов размножения. Так, во многом именно температура определяет, в какой момент слоевища некоторых бурых водорослей (например, пунктарии) из микроскопических и нитевидных развиваются в макроскопические и паренхиматические. При этом имеет значение температура воды в строго определенные моменты жизни водорослей. Так, органы размножения на гаметофитах ламинарии японской (Laminaria japonica) закладываются только при температуре ниже +10 °С, причем достаточно, чтобы она продержалась в течение всего лишь одной ночи. До этого момента и после температура может быть выше (+15... +20 °С).
Температура воды влияет и на глубину произрастания бентосных водорослей. При уменьшении температуры интенсивность дыхания ослабевает быстрее, чем интенсивность фотосинтеза. Это приводит к тому, что компенсационная точка (момент, когда процессы фотосинтеза и дыхания уравновешивают друг друга) устанавливается при меньшей освещенности, т. е. на большей глубине. Поэтому бывают случаи, когда в северном полушарии вид на севере ареала растет на большей глубине, чем на юге.
Температура оказывает влияние на по широтное географическое распространение водорослей. В первую очередь ее действие проявляется косвенным путем - в ускорении или замедлении темпов роста и развития отдельных видов, что ведет к вытеснению одних водорослей другими.
Лишь иногда на границе распространения некоторых видов наблюдаются признаки их морфологического недоразвития, связанного с непосредственным влиянием температуры. Обычно это происходит в Арктике, где общее число видов сокращается и соответственно ослабевает конкуренция между оставшимися видами бентосных водорослей.
Из всего сказанного можно сделать вывод: свет - основной источник энергии для бентосных водорослей как фотосинтезирующих растений, но степень его использования зависит от обеспеченности всех сторон обмена веществ, т. е. от температуры, содержания биогенных и биологически активных веществ, кислорода и неорганических источников углерода, а главное - от темпов поступления этих веществ в слоевище, что находится в зависимости от концентрации веществ и от скорости движения воды. В значительной степени возможность произрастания бентосных водорослей в конкретном месте определяется биотическими факторами - конкуренцией со стороны других водорослей и численностью растительноядных животных. Все это приводит к тому, что отдельные виды бентосных водорослей растут далеко не на всех глубинах, где позволяет им свет, и не во всех водоемах с подходящими световым и гидрохимическим режимами.
В континентальных водоемах в зависимости от приспособленности видов бентосных водорослей к определенным гидрохимическим, световым и гидрологическим условиям, от их способности переносить высыхание сложились сообщества мелких стоячих водоемов (луж, прудов), крупных водоемов (озер, водохранилищ) и текучих вод (ручьев, рек). В глубоких водоемах, где свет не проникает до дна, водоросли населяют прибрежную зону, называемую иногда литоралью (в морской гидробиологии этот термин имеет иной смысл). Нижняя граница прибрежной зоны в континентальных водоемах соответствует границе проникновения света.
Сообщества бентосных водорослей в море выделяются по биотопам (местообитаниям), характеризующимся иными факторами, чем в континентальных водоемах.
В большинстве морей наблюдаются явления прилива и отлива, выражающиеся в периодическом повышении и понижении уровня воды вследствие влияния сил притяжения Луны и Солнца. Приливы случаются одновременно на противоположных сторонах Земли, лежащих вблизи линии, направленной на Луну и Солнце. "Горб", обращенный к небесным телам, появляется в результате прямого притяжения, а "горб" на противоположной стороне Земли вызван наименьшим притяжением. Из-за вращения Земли в каждом пункте на берегу моря в течение суток должно наблюдаться два прилива и два отлива. Приливы не бывают всюду одинаковыми, так как перемещение водной массы (приливной волны) и уровень подъема воды зависят от очертаний суши, широты места, атмосферного давления, ветра и других факторов. Поэтому в разных местах Земли бывают приливы: полусуточные (2 прилива в сутки), суточные (1 прилив в сутки), смешанные с преобладанием по высоте полусуточных приливов, смешанные с преобладанием по высоте суточного прилива. У побережья Баренцева моря, на Кольском полуострове, высота приливов достигает 4 м, в Гижигинском заливе - до 11 м. В то же время во внутренних морях, отделенных от океана узкими проливами, прилив не заметен. Например, в Таллине он меньше 3 см.
Прибрежную часть приливных морей, населенную бентосными водорослями, делят на три зоны: супралиторальная, литоральная, сублиторальная. Литоральная зона (литораль) лежит между крайними уровнями прилива и отлива. Выше расположена супралиторальная зона (супралитораль). Ниже литоральной зоны находится сублиторальная (сублитораль). В бесприливных морях (Черное, Балтийское, Каспийское и др.) тоже выделяют три зоны, но литоралью здесь именуют прибойную зону, которую иногда более правильно называют псевдолиторалью.
Супралитораль бедна водорослями, так как поверхность грунта увлажняется нерегулярно, только брызгами прибоя при сильном волнении. Значительное влияние на развитие водорослей здесь оказывают атмосферные осадки и сток с суши. Поэтому в супралиторали живут в основном микроскопические водоросли, приспособленные к большим колебаниям влажности и солености.
По условиям существования бентосных водорослей и животных литораль делится на три горизонта. Это связано с тем, что уровень моря во время приливов и отливов не постоянен. Два раза в месяц, в новолуние и полнолуние, Земля, Луна и Солнце оказываются примерно на одной линии; при этом происходят наибольшие приливы и отливы, называемые сизигийными. Ими определяются верхняя и нижняя границы литоральной зоны. В фазе первой и последней четвертей Луны, когда силы притяжения Луны и Солнца направлены под прямым углом друг к другу, амплитуда приливов и отливов наименьшая (квадратурные приливы и отливы) (рис. 33). В соответствии с этим различают: I, верхний горизонт - от верхнего уровня сизигийного прилива до верхнего уровня квадратурного прилива; II, средний горизонт - от верхнего уровня квадратурного прилива до нижнего уровня квадратурного отлива; III, нижний горизонт - от нижнего уровня квадратурного отлива до нижнего уровня сизигийного отлива (рис. 34). Как видим, время обсыхания водорослей сокращается от верхнего горизонта к нижнему. Если верхний горизонт заливается водой полностью только в сизигийный прилив, то нижний горизонт, наоборот, полностью обнажается только в сизигийный отлив.
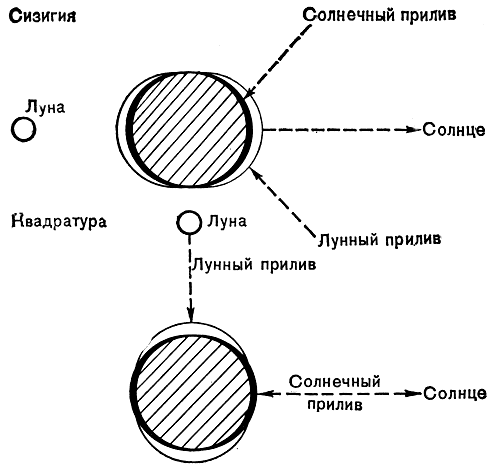
Рис. 33. Объяснение сизигийных и квадратурных приливов
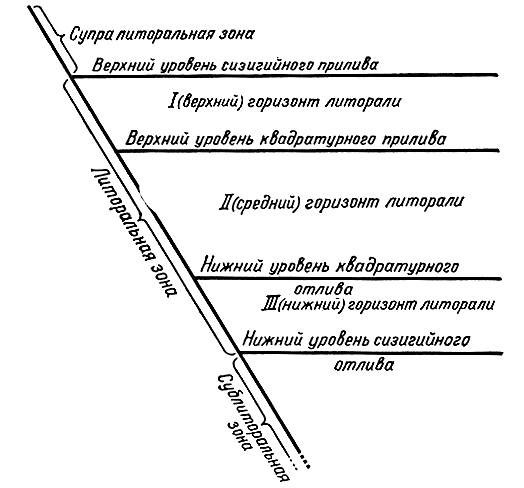
Рис. 34. Колебания уровня моря при приливах и расчленение прибрежной части на зоны
В зависимости от уклона дна и высоты приливов ширина литоральной зоны бывает от нескольких метров до нескольких километров. В среднем и нижнем горизонтах литорали растут уже заросли макроскопических бентосных водорослей, среди которых в морях СССР есть промысловые растения: виды фукуса, аскофиллум узловатый (Ascophyllum nodosum). Для литоральных водорослей характерна способность переносить высыхание, сильные колебания солености и температуры.
Сублитораль простирается от нижнего уровня сизигийного отлива до нижней границы произрастания водорослей. В северных морях ее нижняя граница проходит на глубине 40-50 м; в тропических и субтропических морях при высокой прозрачности воды она может пролегать на глубине до 200 м.
Сублитораль делится на два горизонта: верхнюю сублитораль и нижнюю сублитораль. Для верхней сублиторали характерны густые заросли макроскопических водорослей на твердых грунтах. Это связано с достаточным освещением и постоянным движением воды, вызванным разрушением волн у берега. Нижняя граница верхней сублиторали в морях СССР проходит на глубине 6-10 м, реже 25 м. Нижняя сублитораль характеризуется большим постоянством факторов среды и незначительным освещением. Здесь преобладают красные водоросли, лучше приспособленные к фотосинтезу на больших глубинах, когда остаются в основном зеленые и синие лучи; зеленых и бурых водорослей встречается меньше. В тропиках, однако, на больших глубинах (50-70 м) преобладают сифоновые зеленые водоросли, на этих же глубинах встречаются бурые водоросли, в частности виды рода ламинария. Находки Laminaria rodriguezii в Адриатическом море известны с глубины около 200 м.
В сообществах бентосных водорослей, в основном морских, когда в их состав входят виды с крупными слоевищами, хорошо выражена ярусность. Иногда можно насчитать до 5 ярусов, нижний из которых образован видами с корковидными слоевищами.
Почти на всех макроскопических бентосных водорослях всегда присутствуют эпифиты. Это могут быть тоже макроскопические виды (высотой 10-20 см), но еще больше полумакроскопических (0,5-1 см) и микроскопических водорослей; последние растут как на растении хозяине, так и на других более крупных эпифитах. Эпифитные водоросли могут настолько плотно обрастать слоевище хозяина, что происходит его отмирание и разрушение. Особенно много эпифитов развивается на водорослях с кустистыми слоевищами, растущими в местах с ослабленным движением воды и повышенной концентрацией биогенных веществ.
Из-за широко распространенного у бентосных водорослей явления эпифитизма их заросли можно сравнить с тропическими лесами. Наиболее крупные морские водоросли растут в умеренных и холодных водах. Заросли их бывают настолько густыми, что при погружении в них аквалангисты испытывают чувство неуверенности и страха. Это настоящие подводные джунгли. Только на суше джунгли находятся в экваториальном поясе, а в морях они встречаются в субарктическом, субантарктическом и умеренных поясах.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'