Планктонные водоросли (И. И. Николаев)
Термин планктон (греч. "планктон" - блуждающее) впервые был введен в науку Гензеном в 1887 г. и по первоначальному представлению означал совокупность организмов, парящих в воде. Несколько позднее в составе планктона стали различать фитопланктон (растительный планктон) и зоопланктон (животный планктон). Следовательно, фитопланктоном называют совокупность свободноплавающих (в толще воды) мелких, преимущественно микроскопических, растений, основную массу которых составляют водоросли. Соответственно каждый отдельный организм из состава фитопланктона называют фитопланктером.
Экологи считают, что фитопланктон в жизни больших водоемов выполняет ту же роль, что и растения на суше, т. е. производит первичное органическое вещество, за счет которого прямо или косвенно (через цепь питания) существует весь остальной живой мир на суше и в воде. Это справедливо. Однако следует помнить, что в состав фитопланктона, так же как и в состав наземных растительных сообществ, входят грибы и бактерии, которые, за редким исключением, не способны сами создавать органическое вещество. Они относятся к той же экологической группе гетеротрофных организмов, питающихся за счет готового органического вещества, к которой принадлежит весь животный мир. Грибы и бактерии участвуют в разрушении мертвого органического вещества, тем самым выполняя хотя и очень важную роль в круговороте веществ, но принципиально иную, чем зеленые растения. Несмотря на это, основной функцией фитопланктона в целом следует все же признать создание органического вещества водорослями. Поэтому далее мы будем говорить здесь лишь о микроскопических водорослях, входящих в состав фитопланктона. Это тем более оправдано, что состав грибов в сообществе фитопланктона изучен еще очень слабо, а планктонные бактерии (бактериопланктон) в экологии водоемов рассматриваются обычно отдельно.
Существование планктонных организмов во взвешенном состоянии в воде обеспечивается некоторыми специальными приспособлениями. У одних видов образуются разного рода выросты и придатки тела - шипы, щетинки, роговидные отростки, перепонки и т. п. (рис. 27); у других видов происходит накопление в теле веществ с удельным весом меньше единицы, например капель жира, газовых вакуолей (у некоторых сине-зеленых водорослей, рис. 28) и т. д. Облегчается масса клетки также путем уменьшения ее размеров: размеры клеток у планктонных видов, как правило, заметно меньше, чем у близкородственных донных водорослей. В планктоне широко распространены и мельчайшие организмы, размером в несколько микрометров, образующие так называемый наннопланктон.
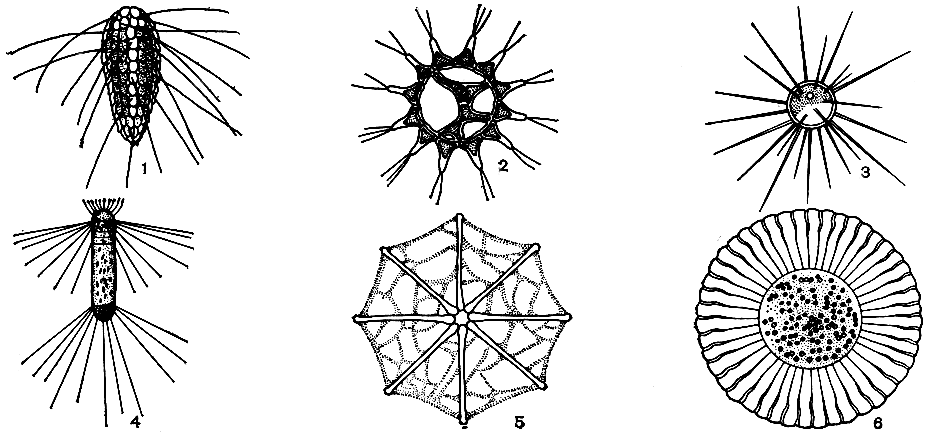
Рис. 27. Внешние приспособления к планктонному образу жизни у водорослей из разных систематических групп: 1-4 - шиповатые формы (1 - Mallomonas, одноклеточный жгутиконосец из золотистых водорослей с окремнелыми чешуйками на оболочке, снабженными отростками; 2 - колониальная зеленая водоросль педиаструм (Pediastrum) с шипами на краевых клетках; 3 - одноклеточная зеленая водоросль голенкиния (Golenkinia) с шипами, усеивающими оболочку; 4 - одноклеточная диатомея коретрон (Corethron) с тремя венчиками отростков на панцире); 5-6 - парашютные формы (5 - звездчатая колония диатомеи астерионеллы (Asterionella) со слизистыми тяжами между клетками, образующими парашют; 6 - одноклеточная диатомея планктониелла (Planktoniella) с плоской формой панциря)

Рис. 28. Планктонные сине-зеленые водоросли с газовыми вакуолями в клетках, вызывающие 'цветение' воды: 1 - две колонии микроцистиса (Microcystis aeruginosa), образованные бесструктурной слизью; 2 - колония воронихинии (Woronichinia naegeliana) с штриховатой наружной слизью; 3, 4 - афанизоменон (Aphanizomenon flos-aquae) (3 - чешуйки из нитей в натуральную величину, 4 - участки нитей при большом увеличении); 5 - собранные в клубочек нити анабены (Anabaena lemmermannii); 6 - плавающие отдельные нити анабены (Anabaena scheremetievii); 7, 8 - колония и отдельная нить глеотрихии (Gloeotrichia echinulata) при разных увеличениях. Газовые вакуоли под микроскопом кажутся черными
Состав и экология отдельных представителей водорослевого фитопланктона в разных водоемах чрезвычайно разнообразны. Фитопланктон существует в водоемах самой разной природы и самых разных размеров - от океана до маленькой лужи. Его нет лишь в водоемах с резко аномальным режимом, в том числе в термальных (при температуре воды выше + 70, + 80 °С), заморных (зараженных сероводородом), чистых приледниковых водах, не содержащих минеральных питательных веществ. Отсутствует живой фитопланктон также в пещерных озерах и на больших глубинах водоемов, где недостает солнечной энергии для фотосинтеза. Общее число видов фитопланктона во всех морских и внутренних водоемах достигает 3000.
В разных водоемах и даже в одном и том же водоеме, но в разные сезоны года число и соотношение видов отдельных таксономических групп весьма различны. Рассмотрим основные его комплексы по главным экологическим категориям водоемов.
Морской фитопланктон состоит в основном из диатомовых и перидиниевых водорослей. Применение методов центрифугирования и осаждения помогло обнаружить в планктоне существенное число мелкоразмерных видов, ранее неизвестных. Из диатомовых водорослей в морском фитопланктоне особенно многочисленны представители класса центрических диатомей (Centrophyceae), в частности роды хетоцерос (Chaetoceros), ризосоления (Rhizosolenia), талассиозира (Thalassiosira), коретрон (Corethron), планктониелла (Planktoniella) и некоторые другие (рис. 29, 1-6), совсем отсутствующие в пресноводном планктоне или представленные в нем лишь небольшим числом видов.
Очень разнообразен в морском фитопланктоне состав жгутиковых форм пирофитовых водорослей, особенно из класса перидиней (рис. 29, 7-10). Эта группа и в пресноводном фитопланктоне довольно разнообразна, но все же насчитывает меньшее число видов, чем в морском, а некоторые роды представлены только в морях: динофизис (Dinophysis), гониаулакс (Goniaulax) и некоторые другие. Весьма многочисленны в морском фитопланктоне также известковые жгутиковые - кокколитофориды, представленные в пресных водах лишь несколькими видами, и встречающиеся исключительно в морском планктоне кремнежгутиковые, или силикофлагеллаты (табл. 9).
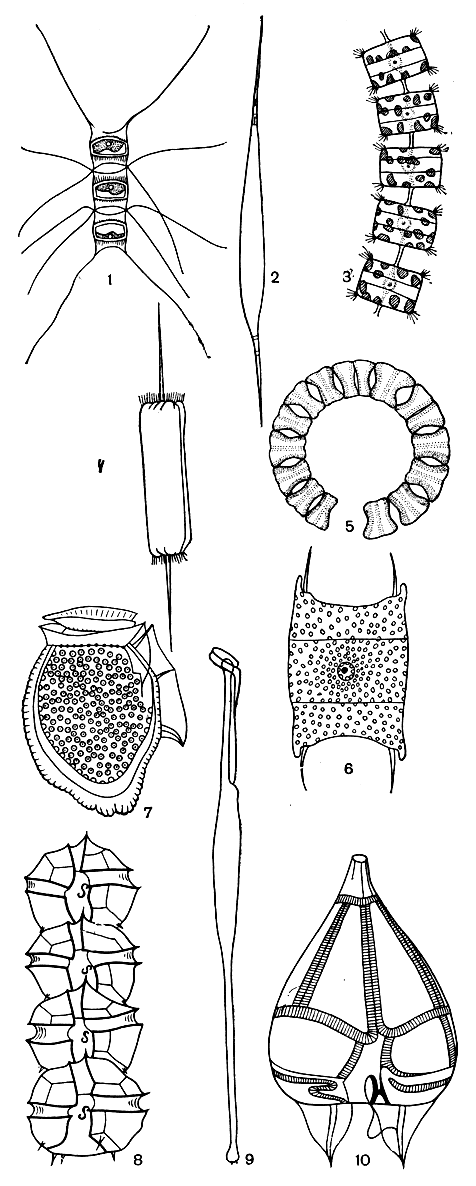
Рис. 29. Морские планктонные диатомеи (1-6) и перидинеи (7-10): 1 - хетоцерос (Chaetoceros); 2 - ризосоления (Rhizosolenia); 3 - талассиозира (Thalassiosira); 4 - дитилиум (Ditylium); 5 - эвкампия (Eucampia); 6 - биддульфия (Biddulphia); 7 - динофизис (Dinophysis); 8 - гониаулакс (Goniaulax); 9 - амфисоления (Amphisolenia); 10 - подолампас (Podolampas)
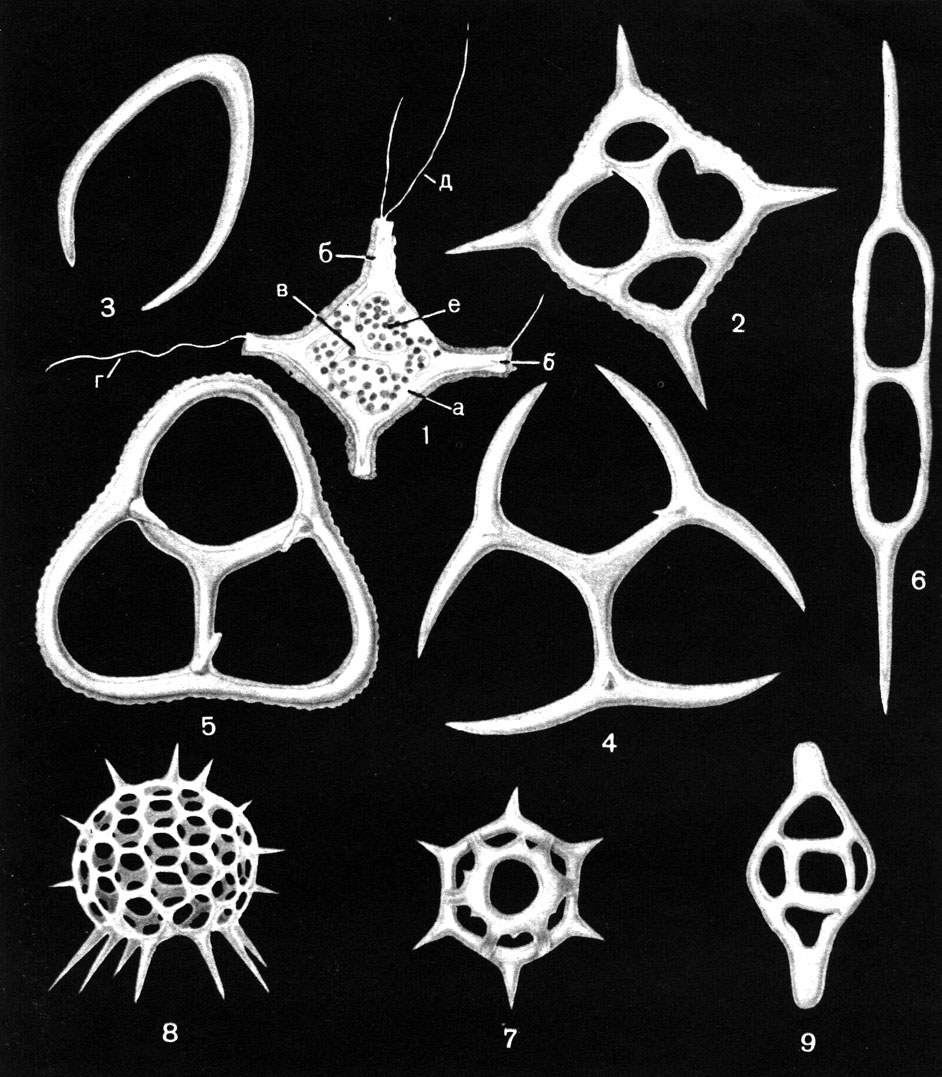
Таблица 9. Основные представители современных и ископаемых силикофлагеллат: 1 - Dictyocha fibula, строение клетки: а - базальное кольцо, б - радиальный рог, в - апикальное образование, г - жгутик, д - псевдоподии, е - хлоропласты; 2 - D. fibula, скелет сверху (палеоген, Западно-Сибирская низменность); 3 - Lyramula simplex, скелет (поздний мел, восточный склон Урала); 4 - Cornua poretzkajae, скелет снизу (поздний мел, восточный склон Урала); 5 - Corbisema apiculata, скелет снизу (палеоген, восточный склон Урала); 6 - Naviculopsis biapiculata, скелет сверху (палеоген, восточный склон Урала); 7 - Distephanus speculum, скелет сверху (неоген, Сахалин); 8 - Cannopilus sphaericus, скелет сбоку (неоген, Венгрия); 9 - Deflandryocha naviculoidea, скелет сверху (неоген, Югославия) (х 1000)
Характернейшей морфологической особенностью представителей морского фитопланктона является образование у них разного рода выростов: щетинок и острых шипов у диатомовых, воротничков, лопастей и парашютов у перидиней. Похожие образования встречаются и у пресноводных видов, но там они выражены значительно слабее. Например, у морских видов церациума (Ceratium) роговидные отростки не только значительно длиннее, чем у пресноводных, но у многих видов они еще и загнуты. Предполагается, что подобные выросты способствуют парению соответствующих организмов. По другим представлениям выросты типа шипов и роговидных образований сформировались как защитное приспособление от выедания фитопланктеров рачками и другими представителями зоопланктона.
Хотя морская среда на значительных пространствах относительно однородна, монотонного распределения фитопланктона не наблюдается. Неоднородность видового состава и различия в численности нередко выражены даже на сравнительно небольших акваториях морских вод, но особенно они рельефны в крупномасштабном географическом распределении. Здесь проявляется экологическое действие основных факторов среды: солености воды, температуры, условий освещенности и содержания питательных веществ.
Для морского тропического фитопланктона характерны наибольшее видовое разнообразие, в целом наименьшая продуктивность (за исключением районов апвеллинга, о которых речь будет далее) и наиболее ярко выраженные морфологические особенности морского фитопланктона (упомянутые выше разного рода выросты). Здесь чрезвычайно разнообразны перидинеи, среди которых встречаются не только отдельные виды, но и целые роды, распространенные исключительно или преимущественно в тропических водах. Тропическая зона - оптимальный биотоп (место существования) и для известковых флагеллат - кокколитофорид. Здесь они наиболее разнообразны и местами развиваются в такой массе, что их известковые скелеты образуют особые донные отложения. Тропические воды по сравнению с холодными водами северных и арктических морей значительно беднее диатомовыми водорослями. Сине-зеленые, как и в других морских районах, представлены очень небольшим числом видов, и только один из них, относящийся к роду осциллатория (Oscillatoria erythraea), в некоторых районах тропиков развивается в таком количестве, что вызывает "цветение" воды.
В отличие от тропиков в полярных и приполярных морских водах в фитопланктоне господствуют диатомовые. Это они создают ту огром-ную массу первичной растительной продукции, на базе которой образуются мощные скопления зоопланктона, в свою очередь служащего пищей самым крупным стадам китов в Антарктике, сельдей и китов в приполярных водах Арктики.
Перидинеи в арктических водах представлены значительно беднее, чем в морях умеренных широт и тем более тропических. Редки здесь и кокколитофориды, зато силикофлагеллаты разнообразны и местами многочисленны. Морские сине-зеленые отсутствуют, в то же время некоторые виды зеленых водорослей развиваются в значительном количестве.
Не менее значительны различия в составе и продуктивности водорослей и в двух других больших биотопах морей, разграниченных в широтном направлении - океанической и неритовой областях, особенно если к последней отнести все внутренние моря. Особые черты океанического планктона перечислены выше. Хотя они и различны в тропических и приполярных водах, но в целом отражают характерные особенности морского фитопланктона. Океанический планктон, и только он, состоит исключительно из таких видов, которые весь свой жизненный цикл совершают в толще воды - в пелагической зоне водоема, без связи с грунтом. В неритовом планктоне таких видов уже значительно меньше, а в планктоне континентальных вод они могут встречаться лишь как исключение.
Неритовая, или шельфовая, зона - это область моря, простирающаяся от берега до конца материковой отмели, что обычно соответствует глубине около 200 м. В некоторых местах она узкая, в других простирается на многие сотни и даже тысячи километров. Главные экологические особенности этой зоны определяются более выраженной связью с берегом и дном. Здесь наблюдаются значительные отклонения от океанических условий в солености воды (чаще в сторону понижения); пониженная прозрачность вследствие минеральной и органической взвеси (нередко за счет более высокой продуктивности планктона); отклонения в температурном режиме; более выраженное турбулентное перемешивание вод и, что особенно важно для растительного планктона, повышена концентрация питательных веществ.
Эти особенности определяют следующие характерные черты в составе и продуктивности фитопланктона неритовой зоны: 1) многие океанические виды выпадают из состава этого сообщества, другие представлены в той или иной степени измененными формами (разновидностями); 2) появляется немало специфических морских видов, не встречающихся в океаническом планктоне; 3) складывается комплекс солоноватоводных видов, полностью отсутствующих в океаническом планктоне, а в сильно опресненных водах некоторых внутренних морей, при солености воды ниже 10-12‰ (‰, промилле - тысячная доля какого-либо числа, десятая доля процента), значительного разнообразия достигают пресноводные виды, которые при опреснении воды до 2-3‰ становятся преобладающими; 4) близость дна и берегов способствует обогащению неритового фитопланктона временнопланктонными (меропланктонными) видами.
Благодаря разнообразию биотопов неритовый фитопланктон в целом значительно богаче по видовому составу, чем океанический. В фитопланктоне неритовой зоны умеренных широт господствуют диатомовые и перидинеи, но среди них немало видов солоноватоводных, в массе развивающихся в опресненных водах внутренних морей (Балтийского, Черного, Азовского и др.). В жизненном цикле многих видов неритового планктона хорошо выражена донная фаза (стадия покоя), что в условиях умеренных широт определяет более четкую сезонную сменность (сукцессию) фитопланктона. В целом неритовый фитопланктон в несколько раз продуктивнее океанического.
Фитопланктон опресненных внутренних морей по составу и продуктивности существенно отличается не только от планктона океанического, но и от типичного неритового. Примером может служить фитопланктон Балтийского моря. Соленость воды в верхнем слое центральной части Балтики 7-8‰, что примерно в 4,5-5 раз меньше солености океана, но в 20-40 раз больше солености пресных вод. В заливах Рижском, Финском и Ботническом соленость понижается до 5-6‰, у берегов - до 3-4‰га в устьях рек и в некоторых бухтах-лиманах (Невская губа, Куршский залив и др.) вода совсем пресная.
Хотя в фитопланктоне Центральной Балтики и даже в открытой части Рижского, Финского и Ботнического заливов преобладает морской комплекс видов, в строгом смысле морским его можно называть лишь по происхождению. Типичные океанические виды здесь отсутствуют полностью. Даже морской неритовый планктон здесь крайне обеднен и представлен лишь эвригалинными видами - способными переносить широкие колебания солености, хотя и предпочитающими низкие значения ее. В этом, морском по происхождению, но солоноватоводном по экологии комплексе балтийского фитопланктона преобладают виды диатомовых: хетоцерос талассиозира, сцелетонема (Sceletonema), актиноциклус (Actinocyclus). Регулярно встречаются, но не достигают большой численности перидинеи: гониаулакс, динофизис (Dinophysis baltica) и несколько видов силикофлагеллат.
В фитопланктоне Центральной Балтики и особенно ее заливов важную роль играет комплекс видов пресноводного происхождения, преимущественно из сине-зеленых: анабена (Anabaena), афанизоменон (Aphanizomenon), нодулария (Nodularia), микроцистис (Microcystis), которые летом в устойчивую солнечную погоду развиваются в такой массе, что даже в центральной части моря образуют "цветение" воды (в основном за счет развития афанизоменона и нодуларии, а в южной части моря также микроцистиса).
В пресноводном комплексе нередки и зеленые водоросли: ооцистис (Oocystis) (по всему морю), виды сценедесмуса (Scenedesmus) и педиаструма (Pediastrum), более многочисленные в заливах.
Пресноводный фитопланктон отличается от типично морского огромным разнообразием зеленых и сине-зеленых водорослей. Особенно многочисленны среди зеленых одноклеточные и колониальные вольвоксовые и протококковые: виды хламидомонад (Chlamydomonas), гониума (Gonium), вольвокса (Volvox), педиаструма, сценедесмуса, ооцистиса, сфероцистиса (Sphaerocystis) и др. (рис. 30). Среди сине-зеленых многочисленны виды анабены, микроцистиса, афанизоменона, глеотрихии (Gloeotrichia) и др.
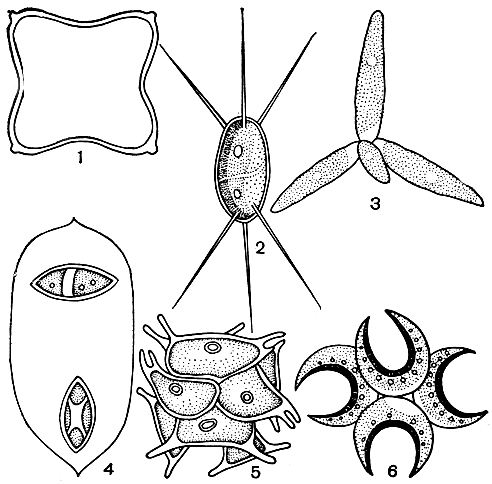
Рис. 30. Пресноводные планктонные протококковые водоросли: 1 - тетраэдрон (Tetraedron); 2 - шодателла (Chodatella); 3 - актинаструм (Actinastrum); 4 - ооцистис (Oocystis); 5 - сораструм (Sorastrum); 6 - селенаструм (Selenastrum)
Видовое разнообразие диатомовых здесь меньше, чем в морях (если не принимать в расчет большое разнообразие временно планктонных видов) (рис. 31); по продуктивности же на единицу поверхности воды роль диатомовых в пресных и морских водах в среднем сопоставима.
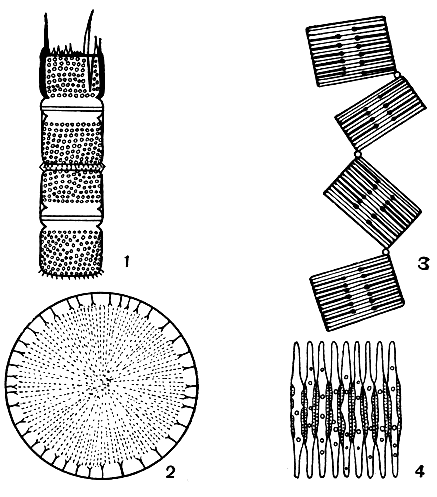
Рис. 31. Пресноводные планктонные диатомеи: 1 - мелозира (Melosira); 2 - стефанодискус (Stephanodiscus); 3 - табеллария (Tabellaria); 4 - фрагилария (Fragilaria)
Характернейший в морском фитопланктоне род хетоцерос (Chaetoceros) в озерах и прудах совсем отсутствует, а многочисленные в морях ризосолении (Rhizosolenia) в пресных водах представлены лишь несколькими видами.
В пресноводном фитопланктоне значительно беднее качественно и количественно представлены перидинеи. Обычны среди них виды церациума (Ceratium) и перидиниума (Peridinium, рис. 64). В пресных водах отсутствуют кремнежгутиковые и очень редки кокколитофориды, зато некоторые другие жгутиковые представлены здесь разнообразно и нередко в большом количестве. В основном это хризомонады - виды динобриона (Dinobryon), малломонаса (Mallomonas), уроглены (Uroglena) и др. (рис. 68, 69), а также эвгленовые - эвглена (Euglena), трахеломонас (Trachelomonas) и факус (Phacus) (рис. 195, 201, 202); первые преимущественно в холодных водах, а вторые - в теплых.
Одной из существенных особенностей пресноводного фитопланктона является обилие в нем временнопланктонных водорослей. Ряд видов, которых принято считать типично планктонными, в прудах и озерах имеют донную или перифитонную (прикрепление к какому-либо предмету) фазу в своем жизненном цикле. Таким образом, разнообразие экологических условий во внутренних водоемах определяет и значительно большее разнообразие по сравнению с морями экологических комплексов и видового состава пресноводного планктона.
В крупных глубоких озерах выражены менее ярко черты отличия пресноводного фитопланктона от морского. В таких озерах-гигантах, как Байкал, Великие озера, Ладожское, Онежское, в фитопланктоне почти круглогодично преобладают диатомовые. Здесь они, как и в морях, создают основную продукцию. Видовой состав диатомового озерного планктона отличен от морского, но в их экологии имеется немало общего. Например, мелозира исландская (Меlosira islandica) - массовый вид фитопланктона Ладожского и Онежского озер, а также мелозира байкальская (Melosira baicalensis) из Байкала в фазу покоя после весенней вспышки не опускаются на дно (или опускаются лишь частично), как это наблюдается у других пресноводных видов в водоемах меньшего размера, а удерживаются в толще воды, образуя на некоторой глубине характерные межсезонные скопления. В больших озерах, как и в морях, велики различия в продуктивности фитопланктона: в центральной части водоема продуктивность очень низка, а у берегов, особенно в мелководных заливах и против устьев рек, она резко повышается.
Еще больше сходства с морским в фитопланктоне двух самых крупных озер мира с соленой водой - Каспии и Арале. Хотя соленость воды в них значительно ниже морской (в Каспии 12-13‰, в Арале 11-12‰), но в составе фитопланктона здесь преобладают водоросли морского происхождения, особенно среди диатомовых: виды хетоцероса (Chaetoceros), ризосолении (Rhizosolenia) и др. Из жгутиковых характерны солоноватоводные виды эксувиеллы (Exuviella) и др. В опресненных зонах этих озер господствуют пресноводные виды, однако при солености воды даже 3-5‰ еще весьма разнообразен солоноватоводный фитопланктон морского происхождения.
В наиболее типичном виде пресноводный фитопланктон как по составу и экологии, так и по продукционным свойствам представлен в озерах среднего размера умеренной зоны, например в озерах Балтийского бассейна. Здесь в зависимости от типа озера и сезона года в фитопланктоне преобладают диатомовые, сине-зеленые или зеленые водоросли. Из диатомовых характерны виды мелозиры (Melosira), астерионеллы (Asterionella), табеллярии (Tabellaria), фрагилярии (Fragilaria), циклотеллы (Cyclotella) и др.; среди сине-зеленых - виды микроцистиса (Microcystis), анабены (Anabaena), афанизоменона (Aphanizomenon), глеотрихии (Gloeotrichia). Основными представителями зеленых водорослей в озерном планктоне являются перечисленные выше протококковые, а в водах с очень мягкой водой, находящихся под влиянием болот, многочисленны десмидиевые: виды космариума (Cosmarium), стаураструма (Staurastrum), клостериума (Closterium), эуаструма (Euastrum) и др. В мелководных озерах и прудах из зеленых водорослей нередко преобладают вольвоксовые: вольвокс (Volvox), хламидомонада (Chlamydomonas), пандорина (Pandorina), эвдорина (Eudorina). В фитопланктоне озер тундры и северной тайги весьма разнообразны хризомонады: виды динобриона (Dinobryon), синуры (Synura), урогленопсиса (Uroglenopsis), малломонаса (Mallomonas). Характернейшая для морского фитопланктона группа перидиней в пресных водах представлена повсюду (во всех водоемах), но относительно небольшим числом видов, которые везде, за редким исключением, достигают невысокой численности. В самых малых водоемах - в небольших озерах и прудах - весьма разнообразны и нередко многочисленны эвгленовые, особенно виды трахеломонаса (Trachelomonas), а в теплых водоемах тропиков и субтропиков также эвглены (Euglena), лепоцинклис (Lepocynclis), факус (Phacus) и др.
В каждом отдельном водоеме в зависимости от физических и химических особенностей режима и от сезона года преобладает одна или другая из перечисленных групп водорослей, а в периоды очень интенсивного развития господствует нередко всего один вид.
В мелких временных водоемах - лужах, выкопанных ямах - весьма обычны мелкие вольвоксовые из рода хламидомонас (Chlamydomonas), от массового развития которых вода нередко окрашивается в зеленый цвет.
В литературе часто выделяют в особую категорию пресноводного планктона речной фитопланктон. В больших реках с очень медленным течением, конечно, водоросли успевают размножаться в пределах ограниченного участка реки при относительно однородных условиях. Следовательно, здесь может сформироваться до некоторой степени особый для данных условий состав фитопланктона. Однако даже в этом случае исходным "материалом" для данного речного сообщества являются организмы, занесенные течением из выше расположенного участка реки или из боковых притоков. Чаще же всего в реке состав фитопланктона формируется как смесь фитопланктона притоков, в той или иной степени преобразованная под влиянием условий реки.
Преобразующая роль условий реки в формировании ее фитопланктона хорошо проявляется, когда большая равнинная река протекает через город или мимо крупного завода, которые загрязняют воду бытовыми и промышленными стоками. В этом случае состав фитопланктона в реке выше города характеризует чистую воду, а в черте города и сразу за его окраиной под влиянием органических загрязнений фитопланктон сильно обедняется и в нем преобладают так называемые сапробные виды - индикаторы сапробных, т. е. загрязненных, вод. Однако ниже, отчасти в связи с осаждением взвешенных органических веществ, отчасти же в связи с распадом их в результате микробиологических процессов, вода становится вновь чистой, а фитопланктон приобретает примерно тот же вид, что и выше города.
На состав и распределение фитопланктона по отдельным водоемам, на его изменение в пределах одного водоема влияет большой комплекс факторов. Первостепенное значение из физических факторов имеют световой режим, температура воды, а для глубоких водоемов - вертикальная устойчивость водных масс. Из химических факторов основное значение имеют соленость воды и содержание в ней питательных веществ, в первую очередь солей фосфора, азота, а для некоторых видов также железа и кремния. Рассмотрим некоторые из перечисленных факторов.
Влияние освещенности как экологического фактора наглядно проявляется в вертикальном и сезонном распределении фитопланктона. В морях и озерах фитопланктон существует лишь в верхнем слое воды. Нижняя граница его в морских, более прозрачных водах находится на глубине 40-70 м и лишь в немногих местах достигает 100-120 м (Средиземное море, тропические воды Мирового океана). В озерных, значительно менее прозрачных водах фитопланктон существует обычно в верхних слоях, на глубине 10-15 м, а в водах с очень малой прозрачностью встречается на глубине до 2-3 м. Лишь в высокогорных и некоторых крупных озерах (например, Байкале) с прозрачной водой фитопланктон распространен до глубины 20-30 м. Прозрачность воды в данном случае влияет на водоросли не прямо, а косвенно, поскольку она определяет интенсивность проникновения в водную толщу солнечной радиации, без которой невозможен фотосинтез. Это хорошо подтверждает сезонный ход развития фитопланктона в водоемах умеренных и высоких широт, замерзающих в зимний период. Зимой, когда водоем покрыт льдом, часто еще со слоем снега, несмотря на самую высокую в году прозрачность воды, фитопланктон почти отсутствует - встречаются лишь весьма редкие физиологически неактивные клетки некоторых видов, а у отдельных водорослей - споры или клетки в стадии покоя.
При общей большой зависимости фитопланктона от освещенности оптимальные значения последней у отдельных видов варьируют в довольно широких пределах. Особенно требовательны к этому фактору зеленые водоросли и большинство видов сине-зеленых, в значительном количестве развивающихся в летний сезон. Некоторые виды сине-зеленых в массе развиваются только у самой поверхности воды: осциллатория (Oscillatoria) - в тропических морях, многие виды микроцистиса (Microcystis), анабены (АпаЬаепа) и др. - в мелких внутренних водоемах.
Менее требовательны к условиям освещенности - диатомовые. Большинство из них избегает ярко освещенного приповерхностного слоя воды и более интенсивно развивается лишь на глубине 2-3 м в малопрозрачных водах озер и на глубине 10-15 м в прозрачных водах морей.
Температура воды - важнейший фактор общего географического распределения фитопланктона и сезонных циклов его, но действует этот фактор во многих случаях не прямо, а косвенно. Многие водоросли способны переносить большой диапазон колебаний температуры (эвритермные виды) и встречаются в планктоне разных географических широт и в разные сезоны года. Однако зона температурного оптимума, в пределах которого наблюдается наибольшая продуктивность, для каждого вида обычно ограничена небольшими отклонениями температуры. Например, широко рас-пространенная в озерном планктоне умеренной зоны и субарктики диатомея мелозира исландская (Melosira islandica) обычно присутствует в планктоне (например, в Онежском и Ладожском озерах, в Неве) при температуре от + 1 до + 13 °С, максимальное же размножение ее наблюдается при температуре от +6 до +8 °С.
Температурный оптимум у разных видов не совпадает, чем и определяется смена видового состава по сезонам, так называемая сезонная сукцессия видов. Общая схема годового цикла фитопланктона в озерах умеренных широт имеет следующий вид. Зимой подо льдом (особенно когда лед покрыт снегом) фитопланктон почти отсутствует в связи с недостатком солнечной радиации. Вегетационный цикл фитопланктона как сообщества начинается в марте - апреле, когда солнечной радиации достаточно для фотосинтеза водорослей даже подо льдом. В это время бывают довольно многочисленными мелкие жгутиковые - криптомонас (Cryptomonas), хромулина (Chromulina), хризококкус (Chrysococcus) - и начинается повышение численности холодноводных видов диатомовых - мелозиры (Melosira), диатомы (Diatoma) и др.
Во вторую фазу весны - с момента вскрытия льда на озере до установления температурной стратификации, что обычно бывает при прогреве верхнего слоя воды до + 10, + 12 °С, наблюдается бурное развитие холодноводного комплекса диатомовых. В первую фазу летнего сезона, при температуре воды от +10 до + 15 °С, холодноводный комплекс диатомовых прекращает вегетацию, в планктоне в это время еще многочисленны диатомовые, но уже другие виды - умеренно тепловодные: астерионелла (Asterionella), табеллария (Tabellaria). Одновременно повышается продуктивность зеленых и сине-зеленых водорослей, а также хризомонад, часть видов которых достигает значительного развития уже во вторую фазу весны. Во вторую фазу лета, при температуре воды выше +15 °С, наблюдается максимум продуктивности сине-зеленых и зеленых водорослей. В зависимости от трофического и лимнологического типа водоема в это время может наблюдаться "цветение" воды, вызванное видами сине-зеленых (Anabaena, Aphanizomenon, Microcystis, Gloeotrichia, Oscillatoria) и зеленых водорослей (Scenedesmus, Pediastrum, Oocystis).
Диатомовые летом, как правило, занимают подчиненное положение и представлены тепловодными видами: фрагиларией (Fragilaria) и мелозирой (Melosira granulata). Осенью, с понижением температуры воды до +10, +12 °С и ниже, снова наблюдается подъем продуктивности холодноводных видов диатомовых. Однако, в отличие от весеннего сезона, в это время заметно большую роль играют сине-зеленые водоросли.
В морских водах умеренных широт весенняя фаза в фитопланктоне также выделяется вспышкой диатомовых водорослей; летняя же - повышением видового разнообразия и обилия перидиней при депрессии продуктивности фитопланктона в целом.
Из химических факторов, влияющих на распределение фитопланктона, на первое место следует поставить солевой состав воды. При этом общая концентрация солей является важным фактором качественного (видового) распределения по типам водоемов, а концентрация питательных солей, прежде всего солей азота и фосфора,- количественного распределения, т. е. продуктивности.
Общая концентрация солей нормальных (в экологическом смысле) природных вод варьирует в очень широких пределах: примерно от 5-10 до 36 000-38 000 мг/л (от 0,005-0,01 до 36-38‰). В этом диапазоне солености выделяются два основных класса водоемов: морские с соленостью 36-38‰, т. е. 36 000- 38 000 мг/л, и пресные с соленостью от 5-10 до 400-500 и даже до 1000 мг/л. Промежуточное положение по концентрации солей занимают солоноватые воды. Этим классам вод, как было показано выше, соответствуют и основные группы фитопланктона по видовому составу.
Экологическое значение концентрации биогенных веществ проявляется в количественном распределении фитопланктона в целом и составляющих его видов.
Продуктивность, или "урожайность", микроскопических водорослей фитопланктона, как и урожайность крупной растительности, при прочих нормальных условиях в очень большой степени зависит от концентрации питательных веществ в окружающей среде. Из минеральных питательных веществ для водорослей, как и для наземной растительности, в первую очередь необходимы соли азота и фосфора. Средняя концентрация этих веществ в большинстве естественных водоемов очень мала, и поэтому высокая продуктивность фитопланктона, как устойчивое явление, возможна лишь при условии постоянного поступления минеральных веществ в верхний слой воды - в зону фотосинтеза.
Правда, некоторые сине-зеленые водоросли способны еще усваивать элементарный азот из растворенного в воде воздуха, однако таких видов немного и их роль в обогащении азотом бывает существенной лишь для очень мелких водоемов, в частности на рисовых полях.
Внутренние водоемы удобряются азотом и фосфором с берега, за счет приноса питательных веществ речной водой с водосборной площади всей речной системы. Поэтому наблюдается четкая зависимость продуктивности озер и мелководных внутренних морей от плодородия почв и некоторых других факторов, действующих в пределах водосборной площади их бассейнов (речных систем). Наименее продуктивен фитопланктон приледниковых озер, а также водоемов, расположенных на кристаллических породах и в районах с большим количеством болот в пределах водосборной площади. Примером последних могут служить озера Северной Карелии, Кольского полуострова, Северной Финляндии, Швеции и Норвегии. Наоборот, водоемы, расположенные в пределах высокоплодородных почв, отличаются высоким уровнем продуктивности фитопланктона и других сообществ (Азовское море, нижневолжские водохранилища, Цимлянское водохранилище).
Продуктивность фитопланктона зависит и от динамики воды, динамического режима вод. Влияние может быть прямым и косвенным, что, однако, не всегда легко различить. Турбулентное перемешивание, если оно не слишком интенсивно, при прочих благоприятных условиях прямо способствует повышению продуктивности диатомовых водорослей, так как многие виды этого отдела, обладая относительно тяжелой оболочкой из кремния, в спокойной воде опускаются на дно. Поэтому ряд массовых пресноводных видов, в частности из рода мелозира, интенсивно развиваются в планктоне озер умеренных широт лишь весной и осенью, в периоды активного вертикального перемешивания воды. При прекращении такого перемешивания, наступающем при прогреве верхнего слоя до +10, + 12 °С и образовании при этом во многих озерах температурного расслоения водной толщи, эти виды из планктона выпадают.
Другие водоросли, прежде всего сине-зеленые, наоборот, не выносят даже относительно слабого турбулентного перемешивания воды. В противоположность диатомовым многие виды сине-зеленых наиболее интенсивно развиваются в предельно спокойной воде. Причины высокой чувствительности их к динамике вод не вполне установлены.
Однако в тех случаях, когда вертикальное перемешивание вод распространяется на большую глубину, оно подавляет развитие даже относительно теневыносливых диатомовых. Связано это с тем, что при глубоком перемешивании водоросли периодически выносятся токами воды за пределы освещенной зоны - зоны фотосинтеза.
Косвенное влияние динамического фактора на продуктивность фитопланктона состоит в том, что при вертикальном перемешивании воды питательные вещества поднимаются из придонных слоев воды, где они не могут быть использованы водорослями вследствие недостатка света. Здесь проявляется взаимодействие нескольких экологических факторов - светового и динамического режимов и обеспеченности питательными веществами. Такая взаимосвязь характерна для природных процессов.
Уже в начале нашего века гидробиологи открыли особое значение фитопланктона в жизни водоемов как основного, а на обширных океанических просторах и единственного производителя первичного органического вещества, на базе которого создается все остальное многообразие водной жизни. Это определило повышенный интерес к изучению не только качественного состава фитопланктона, но и количественного распределения его, а также факторов, регулирующих это распределение.
Элементарный метод количественной оценки фитопланктона, который на протяжении нескольких десятилетий был основным, да и теперь еще не полностью отвергнут,- метод отцеживания его из воды с помощью планктонных сеток. В сконцентрированной таким путем пробе просчитывают количество клеток и колоний по видам и определяют общую численность их на единицу поверхности водоема. Этот простой и доступный метод имеет, однако, существенный недостаток - он не полностью учитывает даже относительно крупные водоросли, а самые мелкие (наннопланктон), которые во многих водоемах значительно преобладают, планктонные сетки не улавливают.
В настоящее время пробы фитопланктона берут в основном батометром или планктобатометром, позволяющим "вырезать" монолит воды с заданной глубины. Сгущение пробы производится методом осаждения в цилиндрах или фильтрацией через микрофильтры: то и другое гарантирует учет водорослей всех размеров.
Когда определились огромные различия в размерах водорослей, составляющих фитопланктон (от нескольких до 1000 мкм и более), стало ясно, что для сравнительной оценки продуктивности фитопланктона по водоемам величинами численности пользоваться нельзя. Более реальным показателем для этой цели является общая биомасса фитопланктона на единицу площади водоема. Однако в дальнейшем и этот метод был забракован по двум основным причинам: во-первых, расчеты биомассы клеток, имеющих у разных видов разную конфигурацию, очень трудоемки; во-вторых, вклад мелких, но быстро размножающихся водорослей в общую продукцию сообщества за единицу времени может быть значительно большим, чем крупных, но медленно размножающихся.
Истинным показателем продуктивности фитопланктона является скорость образования им вещества за единицу времени. Для определения этой величины пользуются физиологическим методом. В процессе фотосинтеза, происходящем только на свету, поглощается углекислота и выделяется кислород. Наряду с фотосинтезом происходит и дыхание водорослей. Последний процесс, связанный с поглощением кислорода и выделением углекислоты, превалирует в темноте, когда фотосинтез прекращается. Метод оценки продуктивности фитопланктона основан на количественном сопоставлении результатов фотосинтеза (процесса продукции) и дыхания (процесса деструкции) сообщества по балансу кислорода в водоеме. Для этой цели используются пробы воды в светлых и темных склянках, экспонируемых в водоеме обычно на сутки на разных глубинах.
Для повышения чувствительности кислородного метода, непригодного для малопродуктивных вод, стали применять изотопную (радиоуглеродную) разновидность его. Однако впоследствии выявились недостатки кислородного метода в целом, и в настоящее время широко применяют хлорофилльный метод, основанный на определении содержания хлорофилла в количественной пробе фитопланктона.
В настоящее время уровень продуктивности фитопланктона многих внутренних водоемов определяется не столько природными условиями, сколько общественно-экономическими, т. е. плотностью населения и характером хозяйственной деятельности в пределах водосборной площади водоема. Эта категория факторов, именуемая в экологии антропогенными, т. е. происходящими от деятельности человека, приводит к обеднению фитопланктона в одних водоемах, а в других, наоборот, к значительному повышению его продуктивности. Первое происходит в результате сброса в водоем токсических веществ, содержащихся в сточных водах промышленного производства, а второе - при обогащении водоема биогенными веществами (особенно соединениями фосфора) в минеральной или органической форме, содержащимися в больших концентрациях в водах, стекающих с сельскохозяйственных территорий, из городов и мелких селений (бытовые стоки). Биогены содержатся и в сточных водах многих промышленных производств.
Второй вид антропогенного влияния - обогащение водоема биогенными веществами - повышает продуктивность не только фитопланктона, но и других водных сообществ, до рыб включительно, и его следовало бы рассматривать как благоприятный с экономической точки зрения процесс. Однако во многих случаях стихийное антропогенное обогащение водоемов первичными питательными веществами происходит в таких масштабах, что водоем как экологическая система оказывается перегруженным биогенами. Следствием этого является чрезмерно бурное развитие фитопланктона ("цветение" воды), при разложении которого выделяется сероводород или другие токсические вещества. Это приводит к гибели животного населения водоема и делает воду непригодной для питья.
Нередки случаи и прижизненного выделения водорослями токсических веществ. В пресноводных водоемах чаще всего это наблюдается при массовом развитии сине-зеленых водорослей, в частности видов рода микроцистис (Microcystis). В морских водах отравление воды нередко вызывается массовым развитием мелких жгутиковых. В таких случаях вода иногда окрашивается в красный цвет, отсюда и название этого явления - "красный прилив".
Понижение качества воды в результате антропогенной перегрузки водоема биогенными веществами, вызывающей чрезмерное развитие фитопланктона, принято называть явлением антропогенной эвтрофикации водоема. Это одно из печальных проявлений загрязнения окружающей среды человеком. О масштабах этого процесса можно судить по тому, что загрязнение интенсивно развивается в таких огромных пресных водоемах, как озеро Эри, и даже в некоторых морях.
Естественное плодородие морских поверхностных вод определяется разными факторами. Пополнение питательными веществами мелко-водных внутренних морей, например Балтийского, Азовского, происходит в основном за счет приноса их речными водами.
Поверхностные воды океанов обогащаются питательными веществами в районах выхода глубинных вод на поверхность. Явление это вошло в литературу под названием апвеллинга. Очень интенсивен апвеллинг у перуанского побережья. На базе высокой продукции фитопланктона здесь чрезвычайно высока продукция беспозвоночных, а за счет этого растет численность рыб. Небольшая страна, Перу в 60-х годах по уловам рыбы вышла на первое место в мире.
Мощная продуктивность фитопланктона в холодных водах арктических морей и особенно в водах Антарктики определяется также подъемом глубинных вод, обогащенных биогенными веществами. Подобное явление наблюдается и в некоторых других районах океана. Противоположное явление, т. е. обеднение поверхностных вод питательными веществами, тормозящее развитие фитопланктона, наблюдается в районах с устойчивой изоляцией поверхностных вод от глубинных.
Таковы основные особенности типичного фитопланктона.
Среди сообществ мелких растений и животных, населяющих толщу воды, существует комплекс организмов, обитающих только у самой поверхности воды - в зоне поверхностной пленки. Этому не столь значительному по видовому составу, но очень своеобразному сооб-ществу в 1917 г. Науман дал особое название - нейстон (греч. "неин" - плавать), хотя, очевидно, оно является лишь составной частью планктона.
Жизнь нейстонных организмов связана с поверхностной пленкой воды, причем одни из них находятся над пленкой (эпинейстон), другие - под пленкой (гипонейстон). Помимо микроскопических водорослей и бактерий здесь обитают и мелкие животные - беспозвоночные и даже личинки некоторых рыб.
Большие концентрации нейстонных организмов вначале были обнаружены в мелких водоемах - в прудах, выкопанных ямах, в небольших заливах озер - в тихую погоду при спокойной поверхности воды. Позднее разнообразные нейстонные организмы, в основном мелкие животные, были найдены и в крупных водоемах, в том числе в морях.
В состав пресноводных водорослей нейстона входят виды разных систематических групп. Здесь обнаружен ряд представителей золотистых водорослей - хромулина (Chromulina), кремастохризис (Kremastochrysis); из эвгленовых - эвглена (Euglena), трахеломонас (Trachelomonas), а также некоторые зеленые - хламидомонада (Chlamydomonas), кремастохлорис (Kremastochloris) - и мелкие протококковые, отдельные виды желто-зеленых и диатомовых водорослей.
У некоторых видов нейстонных водорослей существуют характерные приспособления к существованию у поверхности воды. Например, у видов наутококкуса (Nautococcus) имеются слизистые парашюты, удерживающие их на поверхностной пленке. У кремастохризиса (рис. 32, 1) для этого служит чешуйчатый парашют; у одного вида из зеленых водорослей такой микроскопический парашютик выступает над пленкой поверхностного натяжения в виде конусовидного колпачка (рис. 32, 2).
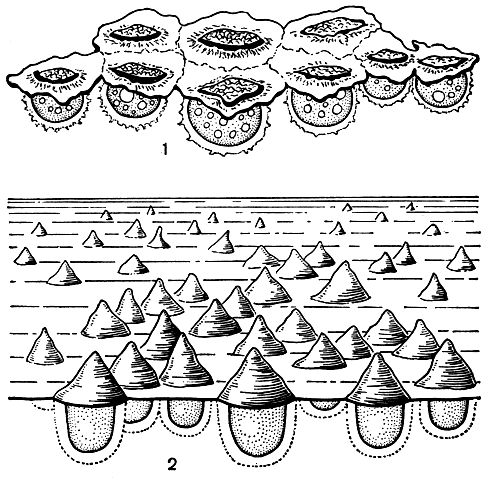
Рис. 32. Водоросли нейстона: 1 - чешуйка кремастохризиса (Kremastochrysis) из нескольких 'парашютов' с висящими под ними клетками, плавающая на поверхности воды; 2 - конические 'парашюты' кремастохлориса (Kremastochloris) на поверхности воды с подвешенными к ним клетками
Преимущества существования нейстонных организмов на границе водной и воздушной сред неясны, тем не менее в отдельных случаях они развиваются в таком количестве, что покрывают воду сплошной пленкой. Нередко и планктонные водоросли (особенно из сине-зеленых) в период массового развития всплывают к самой поверхности воды, образуя огромные скопления. Обнаружены резко повышенные концентрации и водных бактерий. В сообществе нейстона довольно разнообразны и микроскопические животные, которые даже в морях, в условиях почти постоянно волнующейся поверхности, временами образуют значительные скопления у нижнего края поверхности воды.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'