2. Основные экологические группировки водорослей
В силу приспособленности водорослей к различным внешним условиям, они распределяются в разнообразные экологические группировки, характеризующиеся определенной амплитудой экологических факторов и более или менее определенным составом слагающих их водорослей.
Одним из важнейших факторов в распределении водорослей является общая соленость воды. Основную массу природных бассейнов составляют бассейны морские, со средней соленостью в 35 г солей на литр воды, и бассейны пресноводные, в которых содержание солей колеблется в пределах 0,01-0,5 г на литр воды. Такое резкое различие в количестве солей определяет разделение водорослей на морские и пресноводные, причем только очень немногие могут жить как в одной, так и в другой воде. Ниже мы увидим, что по солености можно различить и промежуточные типы водоемов.
В морской воде, вследствие большого постоянства ее химического состава, решающими факторами распределения являются свет и температура. В континентальных водоемах, характеризующихся разнообразными химическими свойствами воды, очень большое значение, наряду с температурой, имеет и химизм.
Однако водоросли, как уже указывалось, могут развиваться не только в водоемах. При наличии хотя бы периодического увлажнения, некоторые из них успешно произрастают на различных наземных предметах, как-то - скалах, деревьях и т. д. Вполне благоприятной средой для развития водорослей во многих случаях является также толща почвенного слоя. Наконец, известны водоросли, обитающие внутри каменистых (известковых) субстратов.
Таким образом, мы должны рассмотреть целый ряд экологических группировок водорослей или, иначе говоря, познакомиться с основными ценозами (сообществами) водорослей. Последние могут быть объединены в две большие группы: 1) водоросли, обитающие в воде и 2) водоросли, обитающие вне воды.
Значительное разнообразие континентальных водоемов в отношении химического состава воды и температурного фактора делает необходимым выделить в пределах этой группы ряд экологических группировок, которые кратко характеризуются ниже.
Прежде всего следует остановиться на водорослевом населении пресных водоемов, физические (температура) и химические (солевой состав) свойства которых лежат в пределах норм, обычных для нашей территории (гидрофитон), В зависимости от характера водоемов (реки, озера, пруды, мелкие водоемы), состав водорослей в них может быть различным, но обычно в пределах каждого бассейна (пресноводного и морского) водоросли могут населять толщу воды или поселяться на дне и на погруженных в воду предметах. В первом случае они входят в состав планктона, как принято называть совокупность организмов, живущих во взвешенном состоянии в воде*, во втором - в состав бентоса, т. е. комплекса форм, в своей жизни тесно связанных с дном водоема.
* (Планктонные организмы, как мы увидим ниже, лишены всяких органов активного движения или обладают ими, но органы эти настолько слабы, что не могут противостоять силе ветра и течений. Живущие в толще воды сильные пловцы (например, рыбы) выделяются в особую группу, называемую нектоном, естественно, не содержащую водорослей.)
К бентосным водорослям (фитобентос) принадлежат все те водоросли, жизнь которых в той или иной мере связана с дном водоемов. Сюда, следовательно, относятся формы, прикрепленные ко дну или целиком лежащие на дне, а также поселяющиеся на различных погруженных в воду предметах. По отношению к обрастанию предметов, введенных в воду человеком - как подвижных (лодки, пароходы и пр.), так и неподвижных (сваи, бакены и т. п.), - часто употребляется специальный термин - перифитон.
В процессе своего расселения бентосные водоросли, даже на небольшом пространстве, встречаются с весьма различными местообитаниями - заиленные участки дна, лежащие на дне камни и т. п. В соответствии с этим и видовой состав их разнообразен. Пресноводные бентосные водоросли в подавляющем большинстве мелкие и даже микроскопические, и только харовые достигают крупных размеров, не уступающих размерам высших водных растений. Многие из них растут, оставаясь всю жизнь прикрепленными ко дну или к погруженным в воду предметам (из зеленых водорослей, например, Stigeoclonium, Ulothrix, многие Cladophora, а также харовые). Другие же (например, зеленые: Spirogyra, Mougeotia, Zygnema некоторые Cladophora и др.) вначале развиваются на дне (не прикрепляясь к нему), а затем крупными дерновинками всплывают на поверхность, куда их поднимает волнение или многочисленные пузырьки кислорода, выделенные ими в процессе фотосинтеза и застрявшие между нитями. Диатомовые водоросли, за исключением небольшого числа колониальных прикрепленных форм, массами лежат на дне или ползают по дну.
Планктонные водоросли (фитопланктон), обитающие в толще воды, очень разнообразны. К тому же в зависимости от характера водоема состав планктона может быть существенно различным, и можно различать планктон мелких водоемов - луж, ям и т. п. (фоссопланктон), прудов (гелеопланктон), озер (лимнопланктон) и рек (потамопланктон).
Основную массу растительного планктона составляют диатомовые и синезеленые водоросли. Все активно плавающие формы тоже являются планктонными, как, например, большинство эвглен, хризомонады, перидинеи и т. п., а также подвижные одноклеточные и колониальные зеленые водоросли монадной структуры. В меньшем количестве встречаются здесь и мелкие неподвижные зеленые водоросли.
Удельный вес протоплазмы несколько больше удельного веса воды. В связи с этим, нахождение в воде во взвешенном состоянии влечет за собой необходимость наличия ряда приспособлений, препятствующих опусканию на дно. Общей особенностью планктонных организмов, способствующей их пловучести, являются их малые размеры. Все они микроскопические или реже видимые невооруженным глазом и измеряемые одним или несколькими миллиметрами. В зависимости от размеров, планктонные организмы могут быть разделены на несколько групп. Среди представителей пресноводного планктона обычно различаются мезопланктонные, микропланктонные и наннопланктонные организмы.
Мезопланктонные организмы имеют тело 1-5 мм величины. Из планктонных водорослей только немногие колониальные формы достигают таких размеров (например, Sphcieronostoc Kihlmani, образующий шаровидные слизистые колонии, содержащие большое количество густо переплетающихся многоклеточных нитей, рис. 68). Большинство же представителей фитопланктона характеризуется размерами, типичными для микропланктонных организмов и лежащими в пределах от 50 μ* до 1 мм. Наконец, к наннопланктонным организмам относятся формы менее 50 μ. Последние, в отличие от двух предшествующих групп, проходят через мелкоячеистую шелковую ткань, применяемую для изготовления сеток, служащих для сбора планктона (см. главу V).
* (Греческой буквой μ (ми) обозначается мера длины микрон, равная 0,001 мм.)
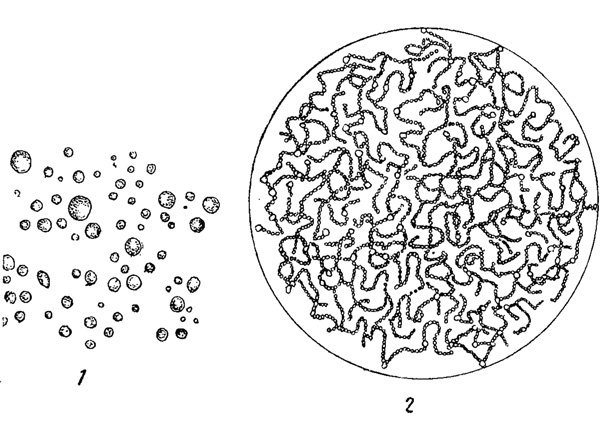
Рис. 68. Синезеленая мезопланктонная водоросль Sphaeronostoc Kihlmani. 1 - колонии в натуральную величину. 2 - колония при небольшом увеличении
Благодаря мелким размерам, а, следовательно, и небольшому весу, планктонные водоросли в воде не так быстро тонут. При малых размерах тела, по сравнению с крупными, как известно, на единицу объема приходится большая поверхность и, вследствие этого, увеличивается трение организма о воду. Но все же мелкие размеры сами по себе еще не могут обеспечить "парения" планктонных организмов в толще воды, в связи с чем мы находим у них и ряд других, весьма разнообразных приспособлений к тому своеобразному образу жизни, который они ведут.
Все планктонные водоросли, прежде всего, делятся на имеющих жгутики, активно плавающих, и лишенных органов движения, причем первых значительно меньше. Однако наличие жгутиков не всегда обеспечивает постоянное поддержание тела во взвешенном состоянии, так что даже у жгутиковых организмов имеется ряд других приспособлений.
У лишенных активного движения планктонных водорослей увеличение пловучести в значительной мере достигается соответствующей формой тела и наличием разнообразных отростков, способствующих "парению" в воде наподобие парашюта. Так, например, имеются планктонные диатомовые, клетки которых слагаются в плоские лентовидные колонии (Tabellaria и Fragilaria, рис. 69). Весьма многочисленны виды (из диатомовых, зеленых водорослей и др.), снабженные длинными шипами (рис. 70). У колониальных форм сами клетки могут слагаться наподобие парашюта, образуя пластинки, как у зеленой водоросли Pediastrum, часть видов которой снабжена еще отростками, или у диатомовой водоросли Asterionella, клетки которой слагаются в форме звезды (рис. 71). Интересно, что между клетками Asterionella протянуты тонкие, нежные слизистые нити (обычно незаметные в воде), способствующие ее поддержанию в водной среде.
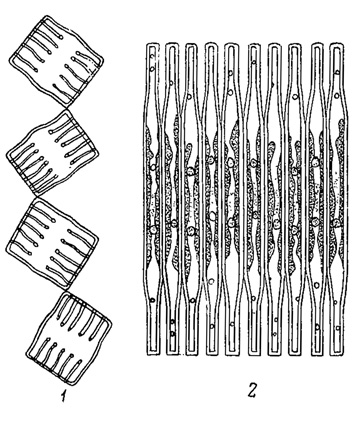
Рис. 69. Планктонные диатомовые водоросли, образующие лентовидные колонии. 1 - Tabellaria flocculoea. 2 - Fragilaria crotoneneis
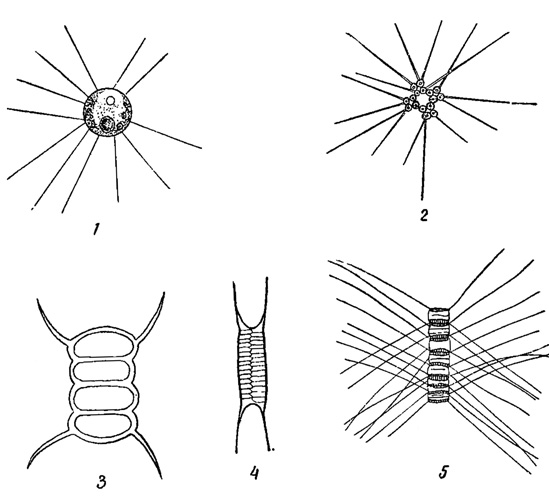
Рис. 70. Планктонные водоросли, снабженные шипами. 1-3 - зеленые водоросли: 1 - Golenkinia radiata, 2 - Richteriella botryoides, 3 - Scenedesmus quadricauda. 4-5 - диатомовые водоросли: 4 - Attheya Zachariasii, 5 - Cyclotella chaetoceros
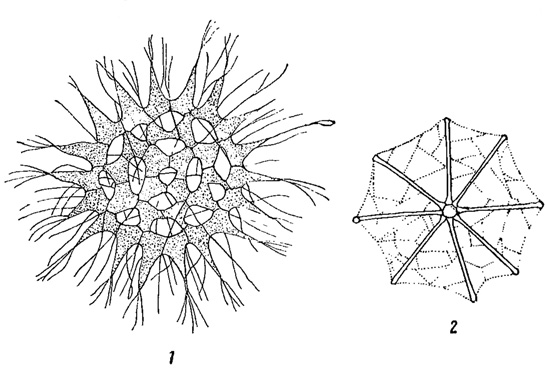
Рис. 71. Планктонные колониальные водоросли парашютной формы. 1 - Pediastrum. 2 - Asterionella
Другим весьма существенным приспособлением к увеличению пловучести является наличие в клетках включений, уменьшающих удельный вес планктонного организма. К числу таких включений относятся жиры, широко распространенные у планктонных форм в качестве запасных питательных веществ, а также газовые вакуоли у синезеленых водорослей (см. выше). Известное значение в этом смысле имеет и слизь, выделяемая многими зелеными и особенно синезелеными планктонными водорослями. Не увеличивая сильно веса, слизь значительно увеличивает объем водоросли, а, следовательно, и ее поверхность трения о воду (рис. 72).
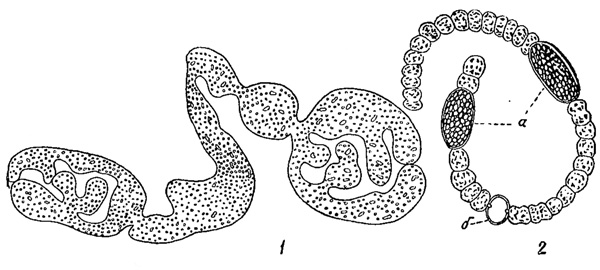
Рис. 72. Приспособления к планктонному образу жизни у синезеленых водорослей. 1 - слизистая колония Coelosphaerium dubium. 2 - газовые вакуоли у Anabaena Scheremetievi. а - споры; б - гетероциста (пограничная клетка)
Как видно, сходные условия существования во взвешенном состоянии в толще воды вызвали схожие приспособления у целого ряда систематически далеких форм.
Не следует думать, что планктонные организмы равномерно развиты во всей толще воды в водоеме. Несмотря на сравнительно большую однородность водной толщи как среды обитания, распределение отдельных экологических факторов в воде может быть неоднородным и меняться с течением времени. Развитие планктона, прежде всего, ограничивается глубиной проникновения света, и в мутных водах он сосредоточен лишь в небольшом слое в несколько метров от поверхности. Развиваясь вначале в поверхностных, лучше освещенных и прогретых слоях воды, планктонные организмы быстро исчерпывают здесь основные питательные соли азота и фосфора, вследствие чего отдельные азотолюбивые формы в дальнейшем становятся более обильными у дна, где имеется подток питательных солей. Иначе говоря, различные планктонные организмы, в зависимости от своих индивидуальных потребностей, могут обильно развиваться в различных участках водяной толщи, меняя местопребывание с течением времени по мере изменения условий. Конечно, в мелких водоемах, где условия значительно более однородны, такие различия в составе и распределении планктона не будут столь резкими или он вообще будет однородным.
Большой интерес представляют также факты вертикальной суточной миграции планктонных водорослей. Так, отмечалось, что фитопланктон в некоторых водоемах, содержась днем в поверхностном слое, вечером и ночью спутается в более глубокие слои воды. Это указывает на то, что даже неподвижные планктонные водоросли обладают известной долей "активности", чутко реагируя своим перемещением в толще воды на внешние условия. В основе подобной миграции, очевидно, лежит изменение удельного веса клеток, но механизм его не вполне ясен. Возможно, что он связан с отдачей кислорода, выделяющегося в процессе фотосинтеза.
Жизнь планктонных организмов в значительной мере протекает сезонно и состав планктона существенно меняется по временам года: одни формы исчезают или сохраняются только в покоящихся стадиях, другие появляются, причем общее количество планктона также не остается постоянным. Основное влияние на эти сезонные изменения оказывает температура, меняющаяся по временам года, а с температурой меняется также и ряд других факторов.
Годовые изменения в составе планктона протекают весьма неодинаково в разных водоемах и в разные годы. Наиболее резко и быстро чередование сезонов сказывается в мелких водоемах.
Как правило, большая часть форм интенсивно размножается в теплое летнее время (например, некоторые синезеленые), но имеются и такие, для которых наиболее благоприятны низкие температуры (например, некоторые хризомонады из золотистых водорослей), так что максимум их развития падает на холодное время года - весну, осень и даже зиму (вегетируют подо льдом).
Весьма интересное явление в жизни планктона представляет так называемое "цветение воды"*. "Цветение воды" наступает тогда, когда внешние условия становятся особенно благоприятными для развития одного, редко двух-трех планктонных форм и неблагоприятными для остальных. Эти немногие формы начинают тогда столь интенсивно размножаться, что в короткое время вытесняют все или почти все другие планктонные организмы и своей массой окрашивают воду в свойственный им цвет. К такому интенсивному развитию способны далеко не все планктонные водоросли, но все же число форм, вызывающих "цветение воды", уже не столь мало, причем вызываемая ими окраска воды весьма разнообразна.
* (Случаи "цветения воды", вызванные водорослями, нужно отличать от "ложного цветения", обусловленного скоплением на поверхности воды пыльцы растений (сосна, ель и др.). Не следует также смешивать с ним часто наблюдающееся в мелких водоемах массовое развитие ряски, относящейся, как известно, к высшим растениям.)
Зеленое "цветение" в мелких водоемах - канавах, лужах, ямах - чаще всего вызывается видами Euglena и Clilamydomonas (рис. 73, 1-2). Особенно быстро и обильно эти организмы развиваются в воде, богатой органическими соединениями. В более крупных водоемах зеленое "цветение" часто обусловливается массовой вегетацией близких к хламидомонадам зеленых колониальных водорослей из порядка вольвоксовых (Volvox, Eudorina и др., рис. 73, 3, 4).
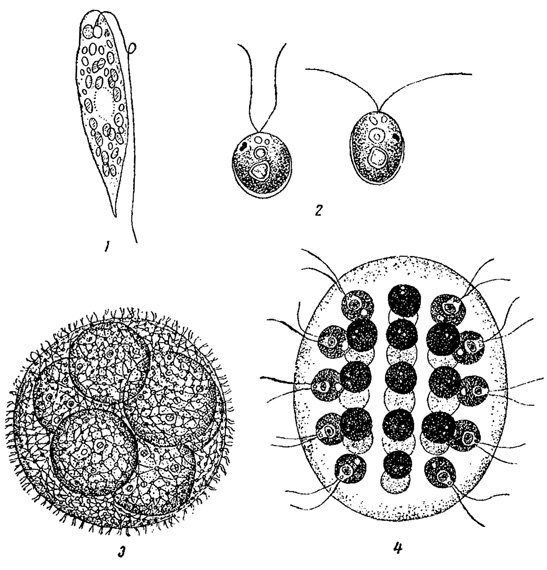
Рис. 73. Водоросли, вызывающие зеленое 'цветение воды'. 1 - Euglena proxima. 2 - Chlamydomonas Reinhardii. 3 - Volvox aureus. 4 - Eudorina elegans
Обычными возбудителями золотисто-желтой или желтовато-коричневатой окраски воды являются некоторые, преимущественно колониальные хризомонады (Synura, Uroglena, Uroglenopsis, Dinobryon, рис. 74), относящиеся к типу золотистых водорослей. "Цветение воды", вызванное хризомонадами, легко узнать по характерному неприятному запаху трескового жира, который они, при массовом развитии, придают воде.
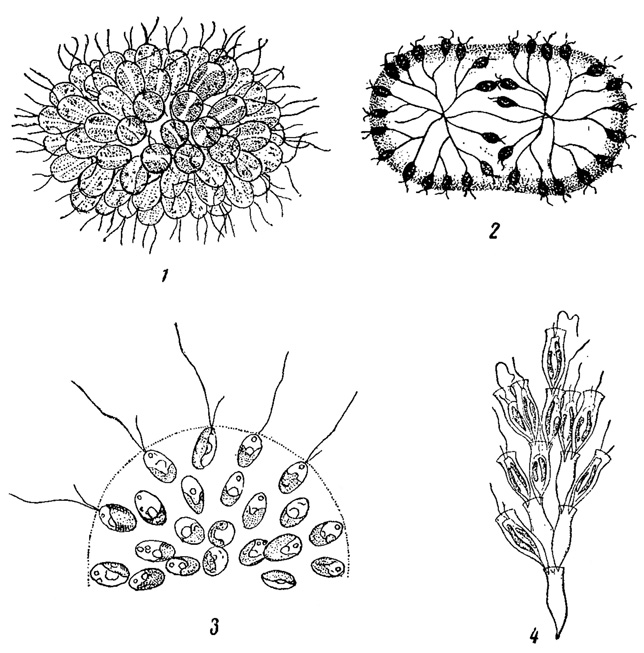
Рис. 74. Водоросли, вызывающие желтовато-коричневатое 'цветение воды'. 1 - Synura uvella. 2 - Uroglena volvox. 3 - Uroglenopsis americana (часть колонии). 4 - Dinobryon sertularia
Коричневатый или буроватый цвет воды часто связан с интенсивным размножением некоторых перидиней (из пирофитовых водорослей) и диатомовых водорослей (рис. 75).
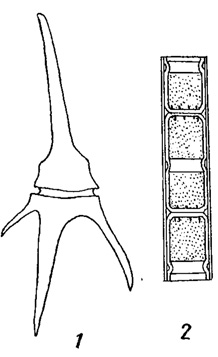
Рис. 75. Водоросли, вызывающие коричневато-буроватое 'цветение воды'. 1 - пирофитовая водоросль Ceratium hirundinella. 2 - часть нити диатомовой водоросли Melosira italica
Наконец, наибольшее количество форм, вызывающих "цветение воды", дают сине-зеленые водоросли, при массовом развитии которых в планктоне вода приобретает сине-зеленый или грязнозеленый цвет (рис. 76). Возбудителями "цветения" могут быть как колониальные (Microcystis, Woronichinia Naegelicina и др.), так и многоклеточные нитчатые формы (Anabaena, Aphcinizomenon flos-aquae, Gloeotrichia echinulata и др.). Клетки планктонных сине-зеленых водорослей обычно забиты многочисленными, кажущимися под микроскопом черными, газовыми вакуолями.
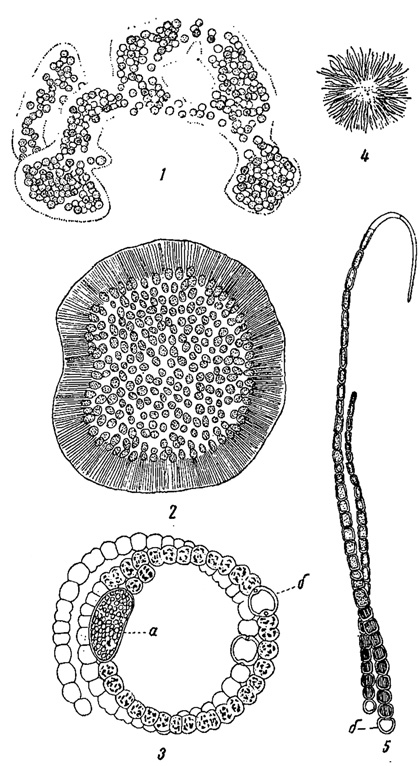
Рис. 76. Водоросли, вызывающие сине-зеленое и грязнозеленое 'цветение воды'. 1 - Microcystis aeruginosa. 2 - Woronichinia Naegeliana. 3 - Anabaena flos-aquae. 4, 5 - Gloeotrichia echinulata: 4 - колония при небольшом увеличении, 5 - две нити из колонии при сильном увеличении, а - спора; б - гетероциста (пограничная клетка)
"Цветение воды" - временное явление в жизни планктона. Это понятно, так как при столь интенсивном развитии особей одного или немногих видов необходимые им питательные вещества будут быстро потребляться, а, кроме того, массовое скопление водорослей создает и неблагоприятные физические условия, как нарушенный газовый обмен, помутнение воды и т. п. Поэтому с известного момента максимального "цветения", из-за недостатка питательных веществ и других причин, неизбежно начинается отмирание части водорослей, что влечет за собой заражение воды продуктами гниения, а это, в свою очередь, еще более усиливает неблагоприятные для данной водоросли условия и ускоряет ее гибель. С другой стороны, по мере освобождения воды от массы особей этого одного или немногих видов, начнут размножаться другие формы, для которых эти условия являются благоприятными. Таким путем постепенно восстанавливаются обычные соотношения организмов, характерные для данного водоема.
Кроме бентоса и планктона в качестве третьей, равноценной им экологической группировки, можно выделить еще нейстон, понимая под последним совокупность организмов, населяющих поверхностную пленку воды. В состав нейстона входят и некоторые, правда, немногочисленные, виды водорослей (некоторые хризомонады, эвглены).
К числу факторов, создающих особые условия для жизни водорослей и обусловливающих своеобразные группировки их, относится повышенное содержание в воде солей, свойственное морским и некоторым континентальным водоемам. В соответствии с задачей настоящего "Определителя", мы ниже будем иметь в виду только последние. Другой ряд своеобразных группировок обусловливает температура.
Флора водорослей водоемов повышенной солености составляет особую группу, которую под именем гидрогалофитона можно противопоставить кратко охарактеризованному нами гидрофитону. Дело в том, что среди континентальных водоемов нередко встречаются такие, в которых концентрация солей значительно выше, чем в обычных пресноводных бассейнах, причем водоемы эти относятся к различным категориям: озерам, прудам, рекам, лужам и болотам. Степень их солености может быть весьма значительной, вплоть до выпадения солей из насыщенного раствора. Так, например, в озерах с преобладанием поваренной соли содержание солей доходит до 285 г на литр воды. И все-таки, даже при таких условиях, живут водоросли*.
* (Для контраста не лишним будет напомнить, что высокое содержание солей настолько неблагоприятно для развития даже таких стойких организмов, как бактерии, что засолка является обычным способом предохранения многих продуктов от порчи.)
В зависимости от солености бассейны делятся на ряд типов, из которых отметим следующие.
1. Пресноводные (олигогалинные), населенные пресноводными видами. Их соленость, как уже отмечалось выше, не превышает 0,5 г на литр воды. Такова большая часть континентальных водоемов.
2. Солоноватые (мезогалинные), с характерными для них солоноватоводными формами (мезогалобами). Соленость колеблется в пределах от 0,5 до 16 г на литр воды. К этой группе относятся многие континентальные водоемы и некоторые моря.
3. Соленые (полигалинные), характеризующиеся содержанием солей от 16 до 47 г (чаще 32-40° г) на литр воды. В эту группу входят моря с населяющими их морскими формами, а также и континентальные водоемы указанной солености. Живущие в них виды мы будем называть солоноводными (эвгалобами).
4. Пересолоненные (ультрагалинные) с соленостью свыше 47 г на литр воды. Формы, обитающие в подобных водоемах, называются ультрагалинными.
Разные виды водорослей по-разному относятся к степени солености и к составу солей в воде. В отношении концентрации солей одни виды выносят широкий диапазон солености и могут жить в бассейнах различной солености (широкосолевые или эвригалинные виды). Другие - наоборот, приспособлены к определенной концентрации солей в воде и развиваются лишь в бассейнах определенного типа солености (узкосолевые или стеногалинные виды).
В общем можно сказать, что количество видов водорослей по мере увеличения солености выше морской убывает и очень высокую соленость переносят только очень немногие из них, но в целом солевыносливых форм не так уж мало. Одним из известнейших представителей последних является, например, Dunaliella salina - одноклеточная зеленая водоросль монадного строения, относящаяся к порядку полиблефаридовых, и представляющая собой типичный узкосолевой ультрагалинный вид (рис. 77, 1). Она обладает красной окраской благодаря наличию в клетках красного пигмента гематохрома, маскирующего хлорофилл. При размножении в массовом количестве, когда клетки Dunaliella отмирают, ее пигменты сообщают солевому раствору (рапе) и выпадающей из него соли в пересолоненных водоемах характерную розовую до красной окраску.
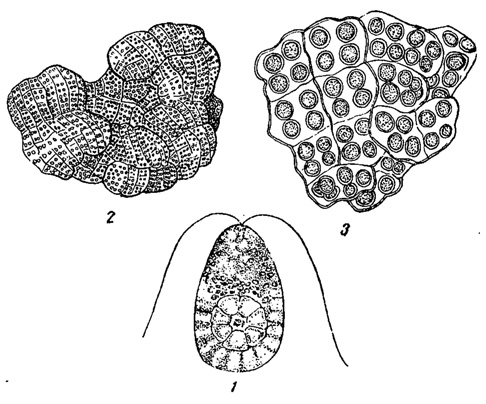
Рис. 77. Водоросли соленых водоемов. 1 - зеленая водоросль Dunaliella salina. 2, 3 - сине-зеленая водоросль Chlorogloea sarcinoides: 2 - общий вид колонии при небольшом увеличении, 3 - часть колонии при сильном увеличении
В водорослевом населении водоемов повышенной солености, так же как и в пресноводных бассейнах, можно различить планктонные и бентосные формы, среди которых, как это вообще характерно для этого рода водоемов, наряду со специфическими обитателями соленой воды, встречаются и пресноводные виды. Последние относятся к эвригалинным организмам, развитие которых стимулируется повышением минерализации (галофилы), или же в известном проценте представляют собой формы, случайно попадающие в соленую воду и могущие жить здесь.
Различное отношение разных видов водорослей к составу солей отчетливо проявляется прежде всего в том, что в континентальных соленых водоемах типично морские формы встречаются редко. В противоположность морским водам, соленые континентальные бассейны отличаются гораздо большим разнообразием своего солевого состава. Кроме вод, богатых хлористым натрием, встречаются водоемы, содержащие большое количество глауберовой соли, карбоната натрия и др. Их концентрация также может иногда достигать весьма значительных размеров. Так, содержание глауберовой соли в некоторых озерах доходит до 347 г на литр воды, что, однако, не исключает развития водорослей. И можно видеть, что в зависимости от химического состава солей в тех или иных водоемах видовой состав водорослей бывает весьма различным.
В жизни соленых водоемов водоросли играют выдающуюся роль. Сочетание образуемой ими органической массы и большого количества солей в воде обусловливает ряд своеобразных биохимических процессов, свойственных этим водоемам. В частности некоторые из водорослей, например, сине-зеленая Clilorogloea sarcinoides (рис. 77, 2, 3), размножаясь в массе, участвуют в образовании лечебных грязей, на чем мы еще остановимся ниже (см. гл. IV).
Гидрогалофитон был противопоставлен нами гидрофитону по признаку повышенного содержания солей в обитаемых им бассейнах. Исходя из другого признака - температуры водной среды, - ему можно противопоставить растительность, с одной стороны, теплых и горячих водоемов, а с другой - снега и льда.
Водоросли теплых и горячих водоемов (гидротермофитон) представляют большой биологический интерес. Связанные с подземными водами горячие источники имеют иногда очень высокую температуру воды, доходящую почти до точки кипения, и оказывается, что даже в таких условиях могут поселяться водоросли, нередко разрастающиеся здесь большими дерновинами сине-зеленого, зеленого или коричневого цвета, плавающими на поверхности воды или выстилающими дно. В более крупных теплых водоемах водоросли обитают и в толще воды, так что здесь также можно различить бентосные и планктонные формы.
Предельные температуры воды, при которых еще были находимы живые водоросли, разными исследователями указываются разные. Для одних источников предел жизни определяется температурой в +52°С, тогда как для других он значительно выше - не менее +80°С и даже до +85°С, а по некоторым данным до +93°С. Кроме высокой температуры эти источники обычно отличаются также высоким содержанием солей, т. е. относятся к числу так называемых минеральных источников. Водоросли нередко поселяются также в сильно загрязненных горячих водах, спускаемых фабриками и заводами.
Самыми типичными обитателями горячих вод являются сине-зеленые водоросли. Среди них имеется наибольшее число видов, живущих в этих условиях и выносящих наиболее высокую температуру. В значительном количестве встречаются также диатомовые, но максимальная температура, допускающая их развитие, ниже, чем для сине-зеленых, и они обычно ютятся в более холодных местах по окраинам водоемов. Среди зеленых водорослей термофильных форм сравнительно немного, но некоторые из них выносят более высокие температуры, чем диатомовые, хотя и уступают в этом отношении сине-зеленым.
Значительный интерес представляет вопрос о видовом составе гидротермофитона. Своеобразие водорослевого населения горячих источников не исключает того, что большинство обитающих здесь водорослей может существовать и при более низких температурах, относясь, следовательно, к эвритермным организмам. Специфически термофильных форм среди них не так уж много, и до настоящего времени неизвестно ни одного вида, в отношении которого можно было бы утверждать, что он не может жить при температуре ниже +30°С.
Условия жизни термофильных водорослей отличаются рядом особенностей. Помимо высокой температуры воды, наибольшее значение имеет то обстоятельство, что они не подвергаются резким переменам температуры, остающейся даже в зимние месяцы выше 0°С, и, следовательно, вегетируют круглый год. Именно в этом некоторые исследователи и усматривают главную отличительную черту термофильных ценозов. К чему они должны быть более всего приспособлены, - это к перенесению высоких температур. Однако никакими специальными внешними признаками, связанными с этим, термофильные водоросли не отличаются, и их исключительная выносливость по отношению к данному фактору, очевидно, обусловливается внутренними физиологическими изменениями в клетках.
Наконец, интересной особенностью многих представителей гидротермофитона является их способность к выделению из воды известковых и кремнистых отложений, на чем мы еще остановимся ниже (см. главу IV).
Прямую противоположность теплолюбивым (термофильным) водорослям представляют водоросли холодолюбивые (криофильные или термофобные), обитающие всю жизнь при низкой температуре, колеблющейся около 0° и еще ниже. Они живут на поверхности снега или на небольшой глубине (несколько сантиметров) в его толще, а также на поверхности льда. Эта своеобразная группировка водорослей может быть, по аналогии с гидротермофитоном, названа гидрокриофитоном.
Мы уже указывали, что многие водоросли, будучи вмерзшими в лед, долгое время могут находиться в состоянии скрытой жизни и, после оттаивания, продолжать свою вегетацию. Вмерзание водорослей в лед - часто встречающееся явление; оно распространяется как на планктонные, так и на бентосные формы, относящиеся к различным типам (зеленые, диатомовые, эвгленовые, сине-зеленые и др.). Подобные комплексы водных организмов, вмерзших в лед (известные под именем пагона), следует, однако, отличать от группировки, обозначенной нами как гидрокриофитон. Для последнего, как отмечено выше, характерно постоянное пребывание при низких температурах, тогда как в состав пагона могут входить случайные формы, вегетирующие в летнее время при температурах значительно выше 0°, хотя в холодных районах криофильные водоросли тоже вмерзают в лед, т. е. входят в состав пагона.
Криофильные водоросли распространены среди снегов и льдов Арктики и Антарктики, а также горных массивов, и развиваются, казалось бы, в крайне неблагоприятных условиях. Большую часть года они находятся в замерзшем состоянии и начинают вегетировать только летом, когда под влиянием солнечных лучей снег и лед подтаивают. При этом размножение их нередко происходит столь интенсивно, что поверхность снега или льда окрашивается в тот или иной цвет, соответственно окраске самих водорослей.
Видовой состав криофильных водорослей довольно разнообразен и включает главным образом представителей зеленых, диатомовых и сине-зеленых водорослей. Некоторые из них распространены широко и в других условиях, но есть и такие, которые встречаются только на снегу и льду.
Наибольшую известность с давних пор получило явление так называемого "красного снега", встречаемого во многих местах (в том числе и в пределах СССР - на Кавказе, в Сибири, па Камчатке, в Арктике), преимущественно по склонам гор. В зависимости от степени развития водорослей, вызывающих эту окраску, она может меняться в своей интенсивности от бледнорозовой до кровяно-красной и темномалиновой и распространяется иногда на огромные пространства (до нескольких квадратных километров).
Главным организмом, вызывающим покраснение снега, является зеленая водоросль монадной структуры (из порядка хламидомонадовых) Chlamydomonas nivalis (рис. 78). Большую часть времени она находится, впрочем, в состоянии неподвижных шаровидных клеток, густо заполненных внутри красным пигментом гематохромом, и подвижность приобретается ею только при оттаивании верхних слоев снега. К Chlamydomoncis nivalis обычно примешиваются и другие виды.
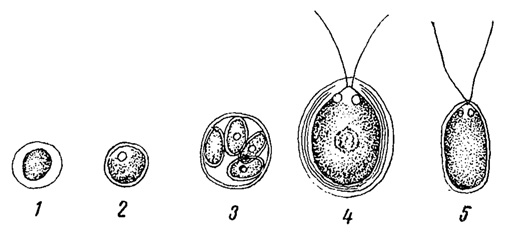
Рис. 78. Водоросль красного снега Chlamydomonas nivalis. 1-3 - в неподвижном состоянии. 4, 5 - в подвижном состоянии
Кроме "красного снега", в настоящее время известно еще много других случаев, когда снег становится окрашенным от развития в нем водорослей. Окраска может быть зеленой, желтой, бурой и даже черной. Интересно, что схожая окраска вызывается иногда разными видами. На льдах в арктических и антарктических бассейнах, главным образом на погруженных в морскую воду частях их, особенно обильно развиваются некоторые диатомовые водоросли, окрашивающие лед в грязнобурый цвет.
Криофильные водоросли подвергаются в зимние стужи очень сильному охлаждению, а в летнее время живут и размножаются в талой воде, т. е. при температуре около 0°. Однако многие из них (например, диатомовые) вовсе не образуют покоящихся стадий, и какие-либо видимые специальные приспособления для перенесения низких температур (так же, как и приспособления к перенесению высоких температур у термофильных водорослей) у них отсутствуют. Следует, однако, отметить, что сама их окраска способствует поглощению солнечных лучей, что обеспечивает более быстрое таяние снега и льда весной в тех местах, где развивается гидрокриофитон.
Водоросли, обитающие вне воды, также весьма разнообразны и развиваются в различной внешней среде. Поэтому и здесь, так же как и по отношению к водорослям, живущим в воде, необходимо указать их главнейшие экологические группировки.
Остановимся сначала на воздушных водорослях (аэрофитон), основной жизненной средой которых является воздух. Они обитают на различных возвышающихся над почвой твердых субстратах, на коре деревьев, на заборах и т. д., причем этот субстрат не оказывает на них сколько-нибудь отчетливого физико-химического воздействия. Источники увлажнения - различны, в зависимости от того, где именно они поселяются. Иногда это атмосферная влага - дождь и роса, иногда же - брызги водопада, прибоя и пр. Увлажнение может достигать разной степени и быть временным или постоянным.
Воздушные водоросли развиваются нередко в массовом количестве. Условия их жизни своеобразны и, в первую очередь, характеризуются частой и резкой сменой основных факторов - температуры и влажности. Днем эти водоросли сильно прогреваются, ночью охлаждаются, а зимой промерзают. В засушливые периоды многие из них высыхают настолько, что могут быть растерты в порошок. Правда, не все упомянутые местообитания в этом отношении одинаковы, и, например, на стенах оранжерей или на орошаемых брызгами скалах условия жизни довольно постоянны, но все же существенно отличаются от водной среды. Понятно, что к таким неблагоприятным условиям приспособились сравнительно немногие организмы, хотя количество видов водорослей в аэрофитоне в общем довольно значительно.
Систематический состав воздушных водорослей различен на разных субстратах. На коре деревьев чаще всего развиваются зеленые водоросли, из которых наиболее широко распространены одноклеточный Pleurococcus и нитчатая Trentepohlici, образующие - первый яркозеленый, а вторая (вследствие наличия в ее клетках гематохрома) - кирпично-красный налет. Часто встречаются также Chlorococcum, Stichococcus, Chlorella и др. (рис. 79).
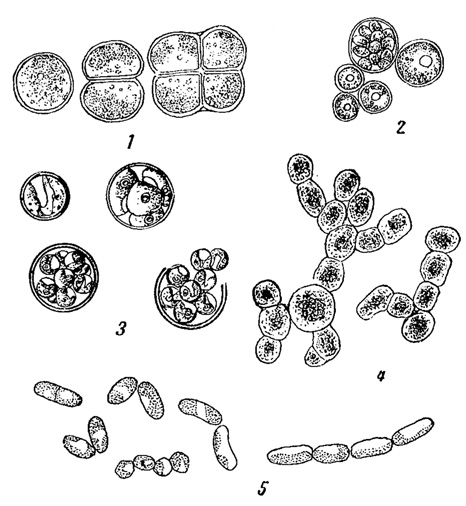
Рис. 79. Водоросли с коры деревьев. 1 - Pleurococcus vulgaris. 2 - Chlorococcum humicola. 3 - Chlorella vulgaris. 4 - Trentepohlia umbrina. 5 - Stichococcus bacillaris
На влажных скалах, наряду с некоторыми, преимущественно одноклеточными зелеными и диатомовыми, особенно обильны синезеленые водоросли, образующие здесь разнообразные налеты и корочки, в сухом состоянии почти черные и крошащиеся, а при увлажнении - слизистые, разнообразной окраски (рис. 80). Тут встречаются как одноклеточные и колониальные (особенно, Gloeocapsa), так и многоклеточные нитчатые их представители (Stigonema, а также Tolypothrix, Scytonemci и др.).
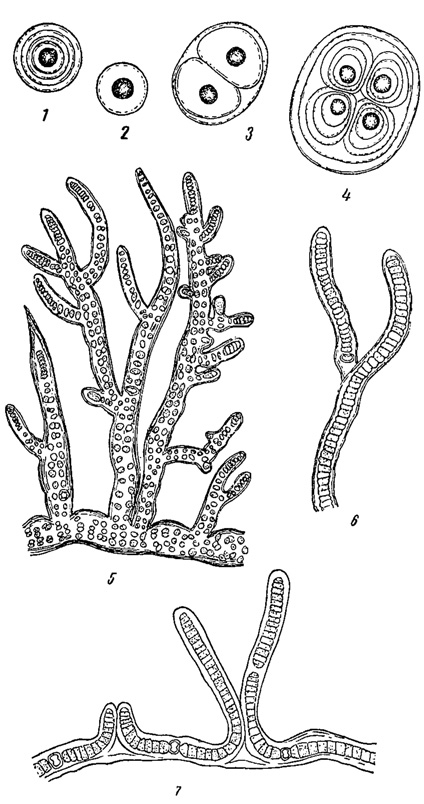
Рис. 80. Водоросли с влажных скал. 1-4 - Gloeocapsa rupestris. 5 - Stigonema minutum. 6 - Tolypothrix byssoidea. 7 - Scytonema ocellatum
Как же все эти водоросли приспосабливаются к наземным условиям существования? Очевидно, что в условиях частой и беспорядочной смены благоприятных и неблагоприятных воздействий, характерных для многих их местообитаний, наилучшим приспособлением будет такое, при котором водоросль целиком может быстро переходить от покоя к вегетации и наоборот. Мы и видим это в действительности у наиболее типичных воздушных водорослей, не образующих, как правило, специальных покоящихся стадий. Выше было указано (см. выше), что приспособление выражается здесь в физиологических изменениях внутри клеток, повышающих их стойкость, которые в разных случаях могут быть различны. Так, у Trentepohlia защитную роль играет накапливающееся в клетках масло, у Pleurococcus - повышение вязкости протоплазмы и т. п. Определенное значение имеют также сильно утолщенные, часто слоистые оболочки (например, у Trentepohlia) или образование широких слизистых обверток, легко пропитывающихся водой, но не так легко ее отдающих, что характерно для многих воздушных синезеленых водорослей (Gloeocapsa, Stigonema и др.).
От воздушных водорослей следует отличать наземные водоросли (геофитон), обитающие непосредственно на почве. В отличие от аэрофитона, геофитон показывает ясную зависимость от химических свойств субстрата (почвы), специфически благоприятный химизм которого и обусловливает иногда его массовое разрастание, что удается наблюдать, например, на подсыхающем илу, на почве, богатой аммиачными соединениями и т. п. Аэрация здесь также обильная, а увлажнение может быть временным (в период интенсивных дождей) или более или менее постоянным (почвы, всегда пропитанные водой). Если влажность увеличивается и создается постоянный слой воды, - эти ценозы естественно переходят в водные, что, впрочем, в более редких случаях встречается и у воздушных водорослей. При непериодическом увлажнении ценозы наземных водорослей часто эфемерны - быстро развиваются и сохраняются недолго. При постоянном увлажнении они могут существовать длительное время.
Систематический состав геофитона различен в зависимости от почвы. На глинистой почве, наиболее благоприятной для развития водорослей, обычным поселенцем является Microcoleus, темно-сине-зеленые слизистые шнуры которого, содержащие внутри множество нитей, расползаются по почве в виде сеточки, заметной простым глазом. Весьма своеобразна флора водорослей на почве степей и пустынь. На некоторых почвах пустынной зоны, в частности, на так называемых такырах, часто встречающихся в Средней Азии, водоросли являются господствующей растительностью, пышно развивающейся во влажные периоды и замирающей летом. Сплошная неисчезающая корочка из отмерших и живых водорослей (преобладают синезеленые) служит характерным признаком такыров. Обильно развиваются водоросли и в полупустынях нашего юго-востока, особенно на поверхности корково-столбчатых солонцов. Наибольшее значение имеет здесь синезеленая водоросль Stratonostoc commune, образующая крупные (до 3-4 см) закругленно-лопастные слизистые колонии, весной и осенью при хорошем увлажнении разрастающиеся иногда, вместе с некоторыми другими водорослями, настолько обильно, что сама почва только проглядывает через образуемый ими налет (рис. 81). Однако в сухое время года эти колонии делаются почти совершенно незаметными, так как высыхают и сморщиваются до состояния тонких, покрывающихся пылью, крошащихся корочек, при увлажнении снова разрастающихся.
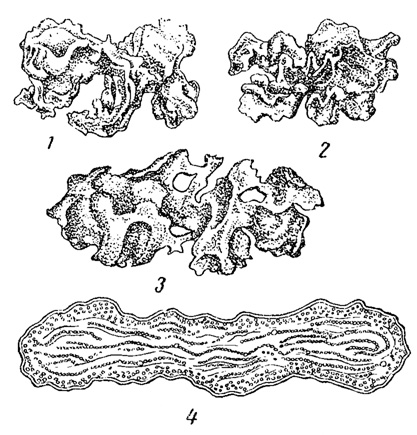
Рис. 81. Наземная водоросль Stratonostoc commune. 1-3 - слоевища в натуральную величину. 4 - поперечный разрез слоевища при небольшом увеличении
Особую флору водорослей мы находим на подсыхающем илу на дне канав, по берегам прудов и т. п. Здесь главенствующую роль играет Botrydium - водоросль сифональной структуры из желтозеленых, темнозеленые шарики которой (размером с булавочную головку) иногда развиваются сплошной массой. Позднее, при подсыхании почвы, Botrydium часто сменяется нитями Vaucheria (зеленая водоросль также сифональной структуры) и некоторыми другими (рис. 82).
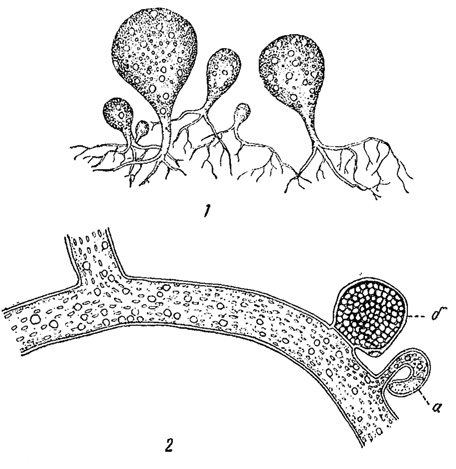
Рис. 82. Водоросли подсыхающего ила. 1 - Botrydium granulalum. 2 - часть нити Vaucheria sp. а - антеридий; б - оогоний
Наконец, можно упомянуть еще водоросли, специфически характерные для почвы, пропитанной азотистыми, в частности аммиачными соединениями (у птичьих базаров, вблизи конюшен, уборных и т. п.). К ним относятся Prasiola (из зеленых), некоторые виды Phormidiam (из синезеленых) (рис. 83) и другие так называемые нитрофильные водоросли.
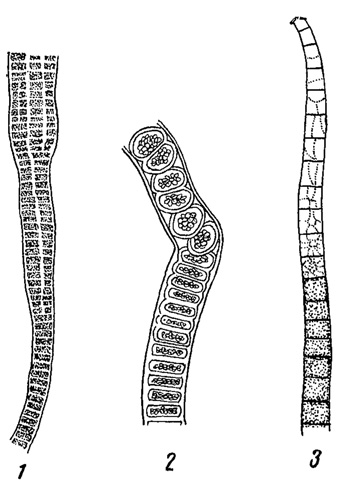
Рис. 83. Водоросли загрязненной почвы. 1, 2 - зеленая водоросль Prasiola сrispa. 3 - синезеленая водоросль Phormidium autumnale
Промежуточное положение между аэрофитоном и геофитоном занимают водоросли влажных мхов. Влажные мхи являются весьма благоприятным субстратом для поселения водорослей, что хорошо можно видеть, например, на севере на огромных пространствах тундры. На них развиваются самые разнообразные водоросли, в частности тот же Stratonostoc commune, широко распространенный в Арктике. Мхи также часто бывают покрыты мезотениевыми и десмидиевыми водорослями (из зеленых водорослей подтипа Conjugatae), нередко образующими здесь, слизистые скопления.
В сравнительно недавнее время было выяснено, что и толща почвенного слоя содержит водоросли. Правда, в большинстве случаев они не видны на глаз, но если небольшую порцию любой, кажущейся совершенно бесплодной почвы поместить в колбу с водой, содержащей необходимые питательные соли (см. гл. V), то в этих благоприятных условиях водоросли быстро размножаются и их яркозеленая масса становится ясно заметной. В естественных условиях такое интенсивное размножение наблюдается только в редких случаях, - например в песках вблизи водоемов или при высоких грунтовых водах, когда на некоторой глубине от поверхности развиваются целые зеленые горизонты, образованные множеством разнообразных водорослей.
Условия существования водорослей в почве крайне своеобразны. Они живут здесь среди почвенных частиц в окружении непрозрачной среды, очень сложной по комплексу факторов, и составляют особую экологическую группировку почвенных водорослей (эдафон), естественно связанную переходами с наземными водорослями (геофитоном).
Состав почвенных водорослей разнообразен, причем количество обнаруженных здесь до сих пор видов более или менее равномерно распределяется между зелеными, синезелеными и диатомовыми водорослями. Большой интерес представляет вопрос о том, насколько специфичен видовой состав эдафона. Исследования последних лет показывают, что только сравнительно немногие виды водорослей могут считаться исключительными обитателями почвенного слоя, не найденными в других экологических условиях, тогда как остальные, более многочисленные виды относятся к числу таких, которые известны как обычные представители водных и воздушных ценозов. Однако далеко не все виды водорослей, живущие в этих условиях, могут развиваться и в почве. Здесь нацело отсутствуют типично планктонные виды, а также некоторые воздушные формы.
Почвенные водоросли обитают преимущественно в поверхностном слое почвы в несколько сантиметров глубины, но некоторые из них могут проникать и значительно глубже. Специальными исследованиями было доказано нахождение водорослей на глубине до 50 см и даже до 1 м, а в отдельных случаях до 2,8 м. Конечно, для водорослей, как фототрофных организмов, поверхностные, освещенные, и глубокие, лишенные света слои почвы являются резко различными местообитаниями. Однако мы уже указывали, что многие водоросли легко переходят на сапрофитное питание, что, в связи с обилием в почве органических веществ, весьма возможно и имеет место в глубоких почвенных слоях. Наряду с этим высказывалось также предположение, что в глубине почв, при полном выключении видимой части спектра, водоросли способны синтезировать органические вещества за счет использования невидимой лучистой энергии и, следовательно, питаться автотрофно.
Наиболее обильно почвенные водоросли развиваются весной и осенью, а в жаркую засушливую погоду и зимой количество их значительно убывает. Интенсивность их развития во многом зависит и от характера почвы. Одним из наиболее существенных факторов здесь является влажность, а также богатство основными питательными веществами, как азот и фосфор. В частности, оказывается, что обрабатываемые удобренные почвы, находящиеся в комковатом состоянии, богаче водорослями, чем необработанные, бесструктурные, а меньше всего видов удалось выявить в сухих песчаных почвах.
Толща почвенного слоя как среда для обитания водорослей по температурно-водному режиму занимает промежуточное положение между вневодными и водными местообитаниями. Самый поверхностный слой почвы по резкости смен температуры и влажности приближается к наземным и даже воздушным условиям, не вполне, однако, им соответствуя. Более глубокие слои почвы отличаются большим постоянством температурно-водных условий, не достигающим все же постоянства, свойственного крупным водным бассейнам. В этом отношении их можно скорее сблизить с мелкими стоячими водоемами. В связи с этим почвенные водоросли приспосабливаются к перенесению неблагоприятных условий частично как вневодные, т. е. при помощи физиологических изменений содержимого клеток или образования слизистых обверток, частично посредством развития специальных покоящихся стадий.
В заключение нашего обзора основных экологических группировок водорослей скажем несколько слов о водорослях, поселяющихся внутри известкового субстрата (эндолитофитон). Это, с одной стороны, так называемые "сверлящие" водоросли, внедряющиеся внутрь известкового субстрата, а, с другой, - водоросли, отлагающие вокруг себя известь. Эти водоросли встречаются как в субстратах, погруженных в воду, т. е. относятся собственно к бентосу, так и вне воды, включаясь тем самым в группировку воздушных водорослей, но в обоих случаях отличаются своеобразным образом жизни внутри каменистого субстрата.
К сверлящим водорослям относятся немногочисленные виды зеленых и синезеленых водорослей, распространенные весьма широко на известковых скалах, камнях, раковинах и т. п. Все они - микроскопические организмы, отличающиеся замечательной особенностью, поселившись сначала на поверхности известкового субстрата, постепенно внедряться в него на глубину до 10 мм и более (рис. 84). Механизм их проникновения состоит в том, что в процессе своей жизнедеятельности они выделяют органические кислоты, растворяющие известь. В результате некоторый слой известковой породы, - а тонкие известковые раковины животных часто и насквозь, - оказывается пронизанным многочисленными каналами. Иными словами, сверлящие водоросли разрушают известковый субстрат, в котором они поселяются. Проникая внутрь субстрата, сверлящие водоросли сохраняют, однако, связь с наружной средой через те каналы, которые они образовали.
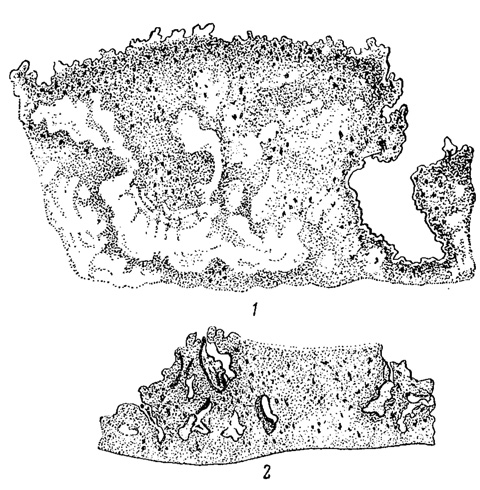
Рис. 84. Два продольных шлифа известковой породы со сверлящими синезелеными водорослями внутри субстрата. Черные точки - колонии Gloeocapsa, черные линии - пучки нитей Scytonema
Прямо противоположный процесс созидания известковых пород осуществляется водорослями, способными выделять известь, что свойственно некоторым синезеленым, зеленым и красным водорослям. Источником извести для них является окружающая вода. В пресной воде, наряду с другими солями, всегда имеется некоторое количество растворимого бикарбоната кальция [Са(НСO3)2], особенно, значительное в жестких водах. В результате отщепления от него углекислоты (используемой в процессе фотосинтеза) выделяется вода и выпадает нерастворимый карбонат (СаСО3)*.
* (В морской воде процесс протекает несколько иначе, и основным источником кальция для известковых водорослей служит сернокислый кальций, превращаемый ими в углекальциевую соль.)
Количество выделяемой водорослями извести различно. Некоторые формы выделяют очень небольшое количество углекислого кальция, который в виде мелких кристаллов располагается между особями или образует футляры вокруг них. Другие же выделяют известь настолько обильно, что частично оказываются совершенно погруженными, как бы "замурованными" в нее, и тогда отмирают, сохраняясь живыми лишь в самых поверхностных слоях тех, подчас очень мощных, отложений, которые они образуют. Так, например, происходит образование известковых туфов в горячих источниках.
В процессе завоевания условий жизни, многие водоросли приспособились не только к обитанию на поверхности или внутри тех или иных субстратов, но и к сожительству с различными другими организмами. Такое сожительство может иметь разный характер. Весьма многочисленны примеры, когда одни водоросли поселяются на других организмах (растениях или животных), пользуясь ими только как местом прикрепления. Эти так называемые эпифитные водоросли широко распространены в природе, встречаясь как в воде, так и вне воды. Однако водоросли могут проникать и внутрь других организмов, сохраняя самостоятельное питание (эндофитные водоросли) или превращаясь в паразитов (например, зеленая водоросль Cephalearos, развивающаяся в листовой ткани некоторых тропических растений). Иногда же они, напротив, сами становятся пищей для той особи, в которой живут (см. ниже). Если такие тесные взаимоотношения делаются постоянными, то возникает симбиоз, под которым мы понимаем тесное сожительство двух или нескольких организмов с прижизненным обменом веществ между ними*. Впрочем, провести резкую границу между всеми этими явлениями трудно.
* (Термин симбиоз иногда применяется и в более узком смысле для обозначения такого рода сожительств, которые обоюдно выгодны для симбионтов. В последнем усматривается отличие симбиоза от паразитизма, при котором выгоду извлекает только один сожитель, тогда как другой, наоборот, является страдающим элементом. Однако, симбиоз в таком понимании вообще в чистом виде едва ли встречается в природе и отграничить симбиоз от паразитизма не представляется возможным.)
Случаи симбиоза встречаются среди водорослей, обитающих как в воде, так и вне воды. Наиболее известным примером симбиоза у воздушных водорослей могут служить многочисленные лишайники, представляющие собой сожительство грибов и водорослей (зеленых или сине зеленых)*. Однако известно немало и других случаев тесного сожительства водорослей с различными растениями и животными. Так, зеленая водоросль из рода Chlorella постоянно поселяется внутри амёб, инфузорий, в клетках гидр, губок и некоторых червей, вследствие чего она получила название зоохлореллы (Zoochlorella). Взаимоотношения обоих симбионтов сводятся к тому, что животное частично переваривает клетки водоросли, а также, вероятно, использует вырабатываемые ею углеводы, а водоросль в известной мере питается азотсодержащими органическими веществами животной протоплазмы. Точно так же некоторые синезеленые водоросли сожительствуют с одноклеточными бесхлорофилльными организмами, часто представляющими собой тоже водоросли, но утратившие хлорофилл. В подобных случаях они функционально заменяют отсутствующие здесь хроматофоры, за которые их иногда ошибочно и принимают.
* (Сравнительно небольшое число лишайников живет в воде.)
Интересны постоянно наблюдающиеся факты развития бактерий внутри слизи различных водорослей (синезеленых и зеленых), что также представляет собой случай симбиоза. Однако здесь не водоросль поселяется внутри других организмов (как в вышеприведенных примерах), а, наоборот, микроорганизмы живут в теле водоросли. Среди обитающих в слизи водорослей бактерий имеются и такие, которые фиксируют свободный азот атмосферы, так что данное сожительство может принести водоросли определенную пользу. Наряду с бактериями, в этих условиях иногда поселяются и мелкие водоросли, т. е. имеет место сожительство разных водорослей друг с другом.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'