3. Строение клетки водорослей
При определении водорослей большое и даже руководящее значение часто имеют признаки, касающиеся строения клетки как основной структурной единицы тела этих растений. Из приведенной выше краткой характеристики типов водорослей видно, что значительное число их является одноклеточными организмами, которые или живут самостоятельно, или образуют временные или постоянные колонии. Далее мы находим огромное количество форм различного облика, показывающих ясно выраженное многоклеточное строение. Только у сравнительно немногих водорослей тело не поделено на клетки (неклеточное или сифональное строение). Но и у этих последних имеются все основные элементы клетки, не отграниченные, однако, клеточными оболочками и многократно повторяющиеся. Важно также, что в цикле их развития нередко имеются одноклеточные стадии, так что клетка и здесь сохраняет свое значение как основной элемент, благодаря развитию и дифференциации которого образуются своеобразно построенные слоевища этих организмов.
Клетки разных водорослей имеют весьма различное строение. Ниже приводится только краткая характеристика основных особенностей клетки водорослей, в которой преимущественное внимание обращено на признаки, имеющие систематическое значение. Более подробное описание строения клетки применительно к отдельным типам будет дано в соответствующих выпусках "Определителя".
Клетки водорослей во многом напоминает клетки высших растений, но, вместе с тем, обладают и рядом специфических черт. Размеры и форма их чрезвычайно разнообразны и являются существенным систематическим признаком. Наибольшего разнообразия достигает форма клеток у зеленых водорослей, особенно в порядке десмидиевых, где в ряду видовых различий она играет исключительно важную роль (рис. 1).
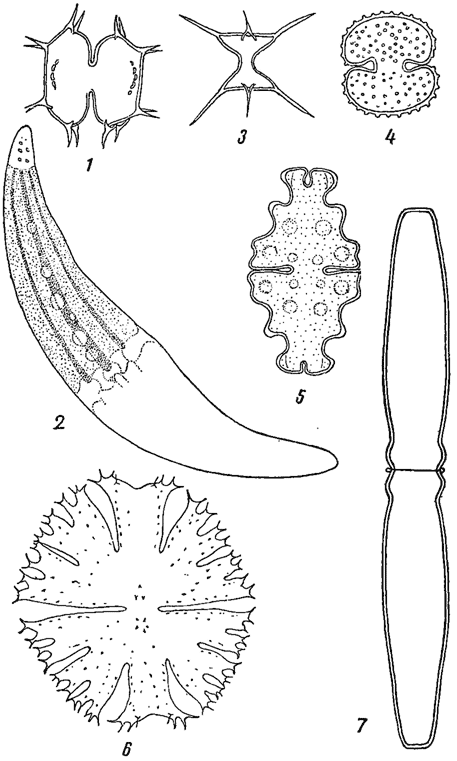
Рис. 1. Одноклеточные десмидиевые водоросли. 1 - Xanthidium antilopaeum var. polymazum. 2 - Closterium moniliferum. 3 - Staurastrum curvatum. 4 - Cosmarium reniforme. 5 - Euastrum affine. 6 - Micrasterias apiculata. 7 - Pleurotaenium trabecula
У подавляющего большинства водорослей протопласт, как и у высших растений, дифференцирован на цитоплазму и ядро, и только у синезеленых водорослей типичные ядра всегда отсутствуют (см. выше). В большинстве случаев клетка содержит одно ядро, но иногда их бывает и несколько. Ядра у многих (но не у всех) водорослей заключают одно или более ядрышек (часто называемых кариозомами) и по своему строению подобны ядрам высших растений или более или менее существенно от них отличаются (рис. 2). Интересной особенностью ядер многих водорослей является то, что хроматин в них часто, даже после окраски, неотличим от ядерного сока (кариолимфы), так что ядро оказывается почти гомогенным (оптически пустым). Форма и местоположение ядер в клетках в некоторых случаях имеют систематическое значение.
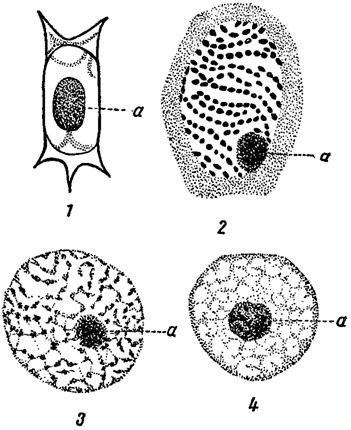
Рис. 2. Ядра из клеток разных водорослей. 1 - Spirogyra. 2 - Glenodinium. 3 - Oedogonium. 4 - Chara. а - ядрышко
Ядро всегда погружено в цитоплазму, выполняющую всю клетку или оставляющую полости, заполненные обычно клеточным соком и называемые вакуолями. Последние достигают иногда значительных размеров и занимают большую часть клетки (например, в порядке зигнемовых из зеленых водорослей). В других случаях вакуоли вовсе отсутствуют, как это характерно для большинства клеток синезеленых водорослей. У некоторых форм вакуоли отличаются некоторыми особенностями, заслуживающими упоминания. Так, у многих видов Closterium (порядок десмидиевых) на концах клетки, имеющей здесь удлиненную форму, часто изогнутой и утончающейся к обоим концам, - располагаются довольно крупные шаровидные вакуоли, в которых видны мельчайшие твердые частицы (кристаллы гипса), находящиеся в интенсивном, так называемом броуновском движении (рис. 1, 2).
Особым типом вакуолей являются пульсирующие вакуоли, нормально присутствующие в подвижных, имеющих жгутики клетках (в частности, в зооспорах и гаметах) во всех группах водорослей и иногда достигающие весьма сложного строения. Они могут встречаться и в неподвижных клетках (у некоторых зеленых и диатомовых водорослей). Эти вакуоли ритмически пульсируют, т. е. попеременно сокращаются и расширяются. Их строение различно. В простейших случаях (у многих золотистых и зеленых водорослей) имеется одна или несколько простых вакуолей, при своем сокращении выбрасывающих содержащуюся в них жидкость наружу - в цитоплазму (откуда она затем поступает в окружающую среду) или, через специальные выводные канальцы, непосредственно за пределы клетки. В других (например, у эвгленовых) - налицо целая система вакуолей, состоящая из главной вакуоли, в которую открывается одна или несколько, вакуолей второго порядка, часто в свою очередь окруженных вакуолями третьего порядка (рис. 3). При сокращении последних жидкость поступает в вакуоли второго порядка, которые, сокращаясь, наполняют главную вакуоль. Выведение жидкости из главной вакуоли осуществляется через выводной канал. У некоторых перидиней встречается еще более сложное строение.
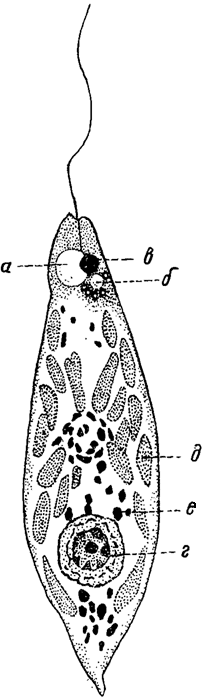
Рис. 3. Euglena. а - главная вакуоль; б - вакуоли второго и третьего порядка; в - стигма; г - ядро; а - хроматофоры; в - парамилоновые зерна
Функция сократительных вакуолей - удаление из клетки излишка воды и, тем самым, осморегуляция. С этим хорошо согласуется тот факт, что они обычно встречаются только у пресноводных водорослей, живущих в среде с незначительным содержанием растворенных веществ, вследствие чего их клетки впитывают большое количество воды.
Наряду с этим, пульсирующие вакуоли, невидимому, выполняют и экскреторную функцию, т. е. выбрасывают из клетки ненужные ей продукты обмена веществ.
Своеобразную форму вакуолей представляют собой газовые вакуоли, находимые в клетках многих синезеленых водорослей (рис. 4). Это - полости в цитоплазме, заполненные газом. Наличие газовых вакуолей уменьшает удельный вес водорослей и они поднимаются в верхние слои воды. Именно в этом многие и усматривают биологическое значение газовых вакуолей. Однако они встречаются и у многих глубоководных синезеленых. В связи с этим было высказано предположение, что газовые вакуоли образуются в результате внутриклеточного брожения, имеющего место в специфических условиях глубинной жизни (в первую очередь, недостатка кислорода). С этой точки зрения "парение" водорослей, обладающих газовыми вакуолями, представляет собой вторичное явление, и они сначала обитают на дне водоема.
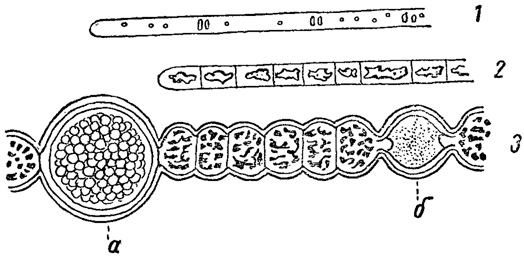
Рис. 4. Газовые вакуоли у синезеленых водорослей. 1 - Оscillatoria putrida. 2 - Oscillatoria Lauterbornii. 3 - Anabaena Scheremetievi. а - спора (покоящаяся клетка); б - гетероциста (пограничная клетка)
Протопласт в большинстве случаев окружен клеточной оболочкой. Впрочем, у водорослей известно не малое количество голых клеток, лишенных настоящих оболочек. Таковы амебоидные клетки, многие водоросли "флагеллатной" организации (эвгленовые, перидинеи и др.), а также большей частью зооспоры, В этих случаях наружный слой цитоплазмы, уплотняясь, образует перипласт (иногда называемый пелликулой). В зависимости от его плотности, клетки или могут изменять свою форму, или она прочно фиксирована. В некоторых случаях перипласт показывает определенную скульптуру (например, у эвгленовых) в виде различным образом расположенных (часто спиральных) штрихов, бородавочек, бугорков и т. п. (рис. 5). У разных форм скульптура перипластов различна и имеет поэтому систематическое значение. У представителей подкласса Coccolithophoreae (относящихся к типу золотистых водорослей) перипласт снаружи окружен более или менее сплошным слоем из округлых известковых телец (кокколитов), лежащих в слизи, выделенной перипластом (рис. 6).
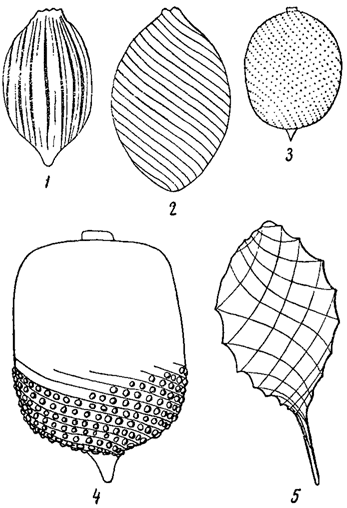
Рис. 5. Скульптура перипласта у эвгленовых водорослей. 1 - Lepocinclis Steinii. 2 - L. fusiformis. 3 - L. ovum var. punctato-striata. 4 - L. quadrata. 5 - Monomorphina pyrum
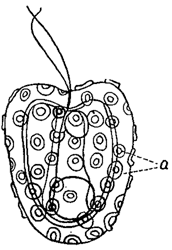
Рис. 6. Hymenomonas roseola. а - кокколиты
Голые клетки у некоторых водорослей (некоторые хризомонады, эвгленовые) выделяют на поверхности особый домик, имеющий специальное отверстие, через которое проходят жгуты (рис. 7). Протопласт часто не заполняет всей его полости и лежит свободно. В отличие от настоящей оболочки, домик не участвует в делении клетки, и образовавшиеся в результате деления два протопласта, или только один из них, выходят наружу и образуют новый домик. Поверхность домика иногда бывает покрыта шипиками, бородавочками и т. п., причем эти особенности, так же как и форма домика, играют большую роль при определении видов (например, в роде Trachelomonas из эвгленовых водорослей).
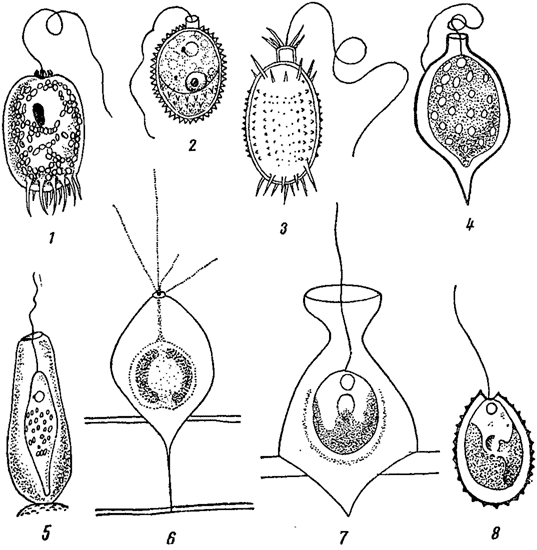
Рис. 7. Домики одноклеточных водорослей. 1 - Trachelomonas armata. 2 - Tr. hispida. 3 - Tr. mirabilis. 4 - Strombomonas acuminata var. verrucosa. 5 - Ascoglena vaginicola. 6 - Chrysopyxis stenostoma. 7 - Chr. cyathus. 8 - Chrysococcus ornatus
Клеточные оболочки у водорослей отличаются значительным разнообразием как в отношении своего строения, так и химического состава. Чаще всего они состоят из клетчатки (целлюлозы), но нередко и из других веществ, среди которых на первое место следует поставить пектиновые вещества. Тот или иной химический состав во многих случаях связан с систематической принадлежностью, и разные группы водорослей характеризуются разным химическим составом клеточных оболочек, как это было частично отмечено выше при характеристике типов этих растений.
Клеточные оболочки нередко показывают более или менее отчетливую слоистость, которая иногда связана с различием в химическом составе разных слоев. Часто внутренние слои состоят из чистой клетчатки, тогда как наружные - из пектиновых веществ. У некоторых водорослей оболочки покрыты шипиками, бородавочками, щетинками и т. п., как это наблюдается у многих хлороцокковых и десмидиевых (из зеленых водорослей, рис. 8). Подобного рода скульптура часто сильно развита на оболочках зигот* (например, у десмидиевых, рис. 9), а также на цистах (хризомонады, рис. 10). Сложная структура оболочки, воспринимаемая в микроскоп в виде штрихов, ребрышек, точек и т. п., особенно характерна для диатомовых водорослей, где она имеет руководящее значение в систематике (рис. 11).
* (См. далее.)
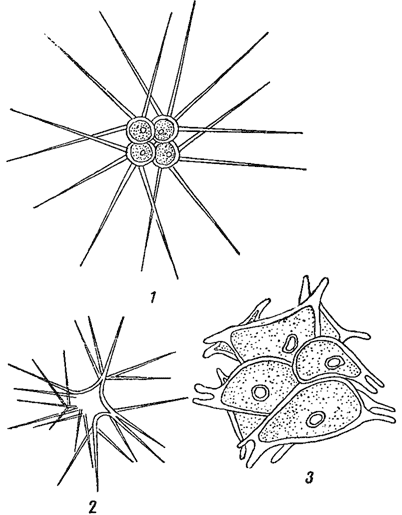
Рис. 8. Хлорококковые водоросли. 1 - Richteriella botryoides. 2 - Tetraëdron spinulosum. 3 - Sorastrum spinulosum
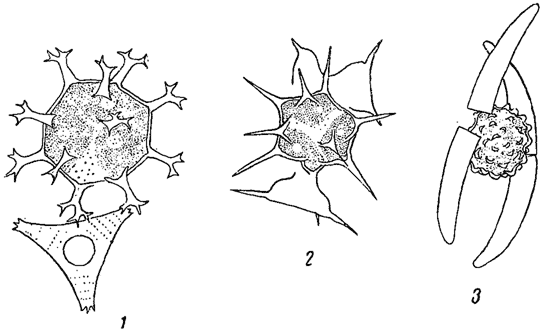
Рис. 9. Зиготы десмидиевых водорослей. 1 - Staurastrum furcigerum. 2 - Arthrodesmus incus var. extensus. 3 - Ciosterium calosporum
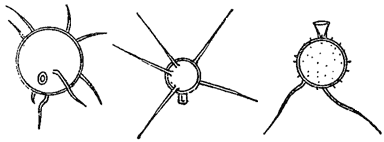
Рис. 10. Цисты хризомонад
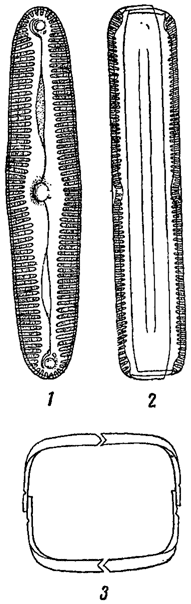
Рис. 11. Панцырь диатомовой водоросли Pinnularia viridis. 1, 2 - панцырь в разных положениях. 3 - поперечный разрез панцыря (схема)
Известны водоросли, клеточная оболочка которых состоит из двух половинок, как это имеет место, например, у желтозеленых водорослей. Оболочки, состоящие из двух (редко больше) частей, характерны и для десмидиевых. Типичной особенностью диатомовых водорослей также является наличие в оболочке двух половинок, из которых каждая, в свою очередь, слагается из двух частей; между последними часто находятся еще промежуточные образования (рис. 11). Сложно построенная составная оболочка диатомовых называется панцырем. Панцырь характерен и для некоторых других групп водорослей, где он имеет, однако, иное строение. Так, панцырем называют весьма сложно построенную оболочку у представителей порядка перидиниевых (из пирофитовых водорослей), состоящую из большого количества закономерно расположенных пластинок (рис. 12). Их число, форма и взаимное расположение является здесь основным систематическим признаком. Наконец, своеобразно построен панцырь у некоторых хризомонад, у которых в наружных слоях оболочки иногда находятся многочисленные чешуйки различной формы, черепичато накладывающиеся друг на друга и часто несущие длинные щетинки (Mallomonas, рис. 13).
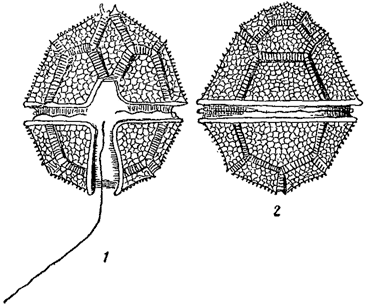
Рис. 12. Peridinium tabulation. 1, 2 - клетка в разных положениях
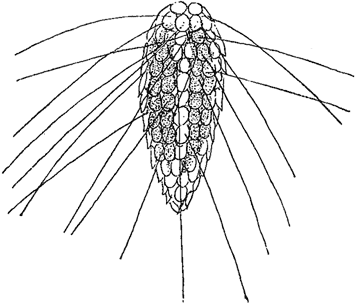
Рис. 13. Mallomonas caudata var. macrolepsis
Клеточные оболочки у водорослей нередко претерпевают разного рода метаморфозы, из которых чаще всего наблюдается ослизнение. Образование большого количества слизи в высшей степени характерно для многих водорослей, относящихся к разным типам. Так, оно сильно развито у синезеленых, а также у красных, многих зеленых и других водорослей. Ослизнение часто обусловливается видоизменением наружных слоев оболочки, состоящих из пектиновых веществ. Наряду с этим слизь может образоваться и за счет цитоплазмы, выходящей наружу через имеющиеся в оболочке поры.
Слизь часто объединяет отдельные клетки и целые многоклеточные нити в колонии. Иногда она окружает отдельные нити, образуя вокруг них слизистые футляры (рис. 14), как это имеет место у многих нитчатых синезеленых водорослей (класс Hormogoneae), а из зеленых - у представителей порядка зигнемовых и некоторых десмидиевых. Биологическое значение слизи разнообразно. Она предохраняет клетки водорослей от высыхания. У форм, обитающих в быстро текущей воде, слизь может защищать их от механических повреждений. У некоторых десмидиевых (Closterium, Cosmarium) одностороннее выделение слизи из клетки (через специальные так называемые слизевые поры) обусловливает их движение (рис. 15). Слизь способствует прикреплению некоторых водорослей к субстрату и т. п.
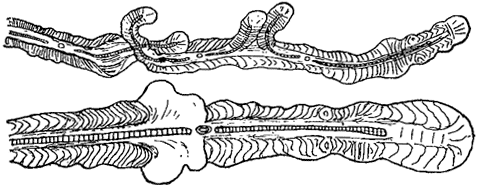
Рис. 14. Синезеленая водоросль Scytonema alatum со слизистыми футлярами
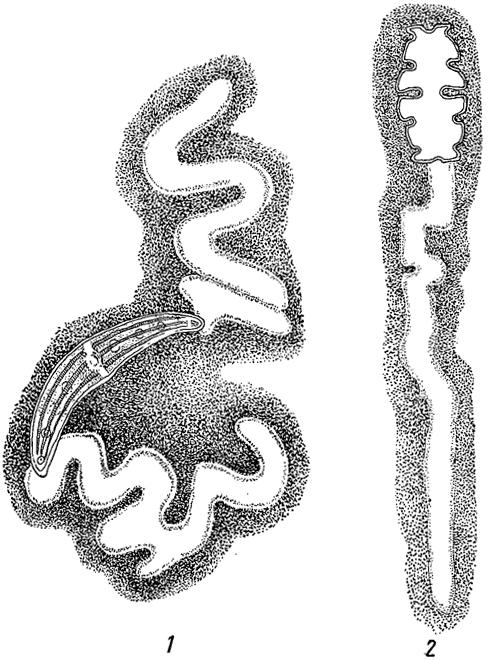
Рис. 15. Выделение слизи десмидиевыми водорослями. 1 - Closterium moniliferum. 2 - Euastrum oblongum. Клетки помещены в тушь, не проникающую внутрь слизи, вследствие чего она становится отчетливо видимой
Кроме ослизнения, клеточные оболочки могут претерпевать и другие изменения. Так, иногда наблюдается кутикуляризация наружного слоя оболочки (особенно в покоящихся спорах)*. Нередко в оболочках или на оболочках откладываются различные соли, чаще всего углекислый кальций. Это характерно для харовых, дирофитовых, некоторых синезеленых и, реже, зеленых водорослей (например, Oocardium из десмидиевых, рис. 16). Особенно сильной степени отложение извести достигает у ряда морских красных и зеленых водорослей. Клеточные оболочки могут также пропитываться кремнеземом. Его присутствие в высшей степени типично для диатомовых водорослей, у которых внутренний слой оболочки слагается из пектинового вещества, а наружный - из кремнезема. Отложение кремнезема наблюдается и у золотистых водорослей - им пропитаны оболочки цист у хризомонад, чешуйки и щетинки панцыря у Mailomonas (см. выше) и др., а также у желтозеленых водорослей. В оболочках некоторых водорослей (например, десмидиевых, трахеломонад) откладываются соли окиси железа.
* (При кутикуляризации наружный слой оболочки пропитывается особым кутикулярным веществом - кутином, близким к пробковому веществу - суберину.)
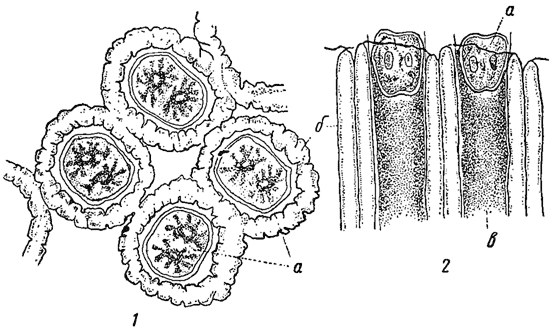
Рис. 16. Oocardium stratum. 1 - вид колонии сверху. 2 - колония в продольном разрезе. Клетки находятся в слизистых трубочках, инкрустированных известью, а - клетки водоросли; б - известковые трубочки; в - слизь
Во многих случаях в оболочках удается констатировать наличие пор, часто весьма мелких. У многоклеточных водорослей через поры во внутренних клеточных перегородках тончайшие тяжи протоплазмы (плазмодесмы) связывают между собой в единое целое протопласты отдельных клеток. Частным случаем пор являются отверстия в оболочке, через которые выходят жгуты.
Выше мы указывали, что образование слизи вокруг тела некоторых водорослей иногда обусловлено выхождением наружу цитоплазмы, осуществляемом через поры. В этом отношении особый интерес представляют, также упомянутые выше, слизевые поры, встречающиеся у десмидиевых, диатомовых и некоторых других водорослей. Слизевые поры могут равномерно распределяться по поверхности клетки или располагаться в группах, иногда вокруг бородавчатых утолщений оболочки, как это имеет место у некоторых десмидиевых (рис. 17).
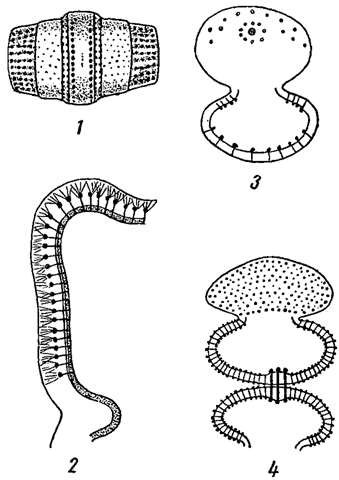
Рис. 17. Слизевые поры в оболочках клеток десмидиевых водорослей. 1 - Gomnozyga moniliformis. 2 - часть клеточной оболочки Micrasterias. 3 - Coemocladium saxonicum. 4 - Sphaerozosma
Большой сложности поровый аппарат достигает у диатомовых водорослей (строение его будет рассмотрено в выпуске "Определителя", посвященном этой группе).
Одним из наиболее существенных элементов клетки водорослей являются носители пигментов - хроматофоры. Их наличие в высшей степени характерно для водорослей как фототрофных растений. Хроматофоры отсутствуют только у небольшого числа форм, вторично утративших их и питающихся, в связи с этим, гетеротрофно. Морфологически оформленных хроматофоров лишены клетки синезеленых водорослей, у которых пигменты локализированы в хроматоплазме (см. выше). В отличие от высших растений хроматофоры водорослей отличаются большим разнообразием, и их форма и строение, а также местоположение в клетке (центральное, осевое, постенное) оказываются здесь одним из основных систематических признаков, так как в пределах видов они относительно постоянны.
Как известно, хроматофоры являются частным случаем более широкого понятия пластид и, содержа хлорофилл, представляют собой органоиды, обеспечивающие процесс фотосинтеза. О находящихся в них пигментах, различных у разных водорослей, а также о продуктах фотосинтеза, являющихся конечным результатом их деятельности, и тоже весьма разнообразных, - было сказано выше, при характеристике типов водорослей.
Хроматофоры делятся и передаются из материнских клеток в дочерние при их размножении. Они могут также образоваться из бесцветных пластид - лейкопластов в результате выработки ими пигментов, как это наблюдается у некоторых Euglena, в зависимости от условий среды то теряющих хлорофилл, то снова вырабатывающих его. Клетки многих водорослей содержат один хроматофор, но иногда их бывает и много.
Одним из простейших типов хроматофора является пластинчатый (например, у Mougeotid, Ulothrix). Близок к нему чашевидный, представляющий собой не что иное как изогнутую в соответствии с формой клетки пластинку (например, у многих видов Chlamydomonas). Края пластинки оказываются подчас более или менее изрезанными, так что хроматофор приобретает вид рассеченной пластинки (например; у Draparnaldia). Еще более сложны сетчатые хроматофоры (например, у Cladophora). Весьма характерны лентовидные хроматофоры, иногда спирально извитые (например, у Spirogyra). Известны также хроматофоры дисковидные (например, у Vaucheria, многих Euglena), звездчатые (например, у Zygnema) и мн. др. (рис. 18, 19). Приведенными примерами далеко не исчерпывается все разнообразие формы хроматофоров, особенно значительное среди зеленых водорослей.
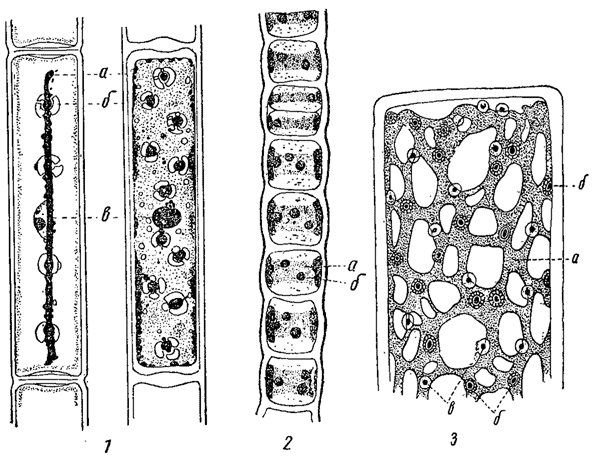
Рис. 18. Форма хроматофоров у разных водорослей. 1 - две клетки Mougeotia scalaris с хроматофором в разных положениях. 2 - часть нити Ulothrix zonata. 3 - часть клетки Cladophora. а - хроматофор; б - пиреноид; е - ядро
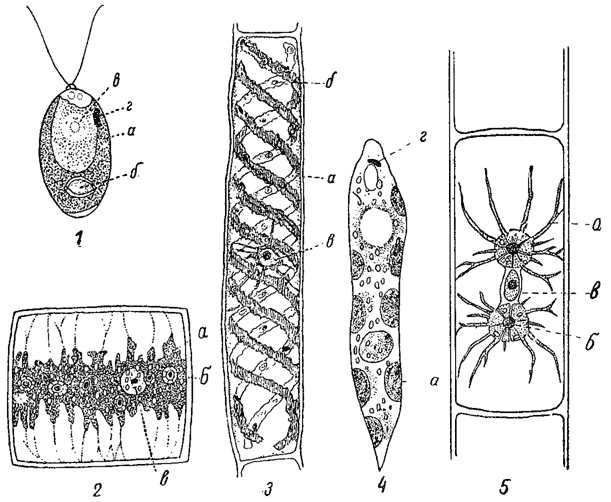
Рис. 19. Форма хроматофоров у разных водорослей. 1 - Chlamydomonas biconvexa. 2 - клетка Draparnaldia glomerata. 3 - клетка Spirogyra. 4 - Euglena deses. 5 - клетка Zygnema. а - хроматофор; б - пиреноид; в - ядро; г - стигма
Продукты фотосинтеза, образуемые в клетке, могут откладываться в хроматофорах или же вне их - в цитоплазме или же, наконец, в клеточном соку. Местом отложения крахмала являются обычно хроматофоры. Зернышки крахмала откладываются здесь или непосредственно в хроматофорах или вокруг особых телец, называемых пиреноидами*. Пиреноиды находятся внутри хроматофоров или на их поверхности; гораздо реже они расположены в цитоплазме (часто у криптомонад). Они размножаются делением, но могут возникать и путем новообразования. Форма и строение пиреноидов разнообразны. Различают пиреноиды скорлупчатые и голые (рис. 20). Первые в развитом состоянии окружены покровом из ассимилятов (крахмал или близкие к нему углеводы), вторые - лишены его и их функция не может считаться вполне выясненной. Наличие пиреноидов того или другого рода имеет известное систематическое значение. Голые пиреноиды особенно характерны для золотистых, диатомовых, бурых и красных, скорлупчатые - для большинства зеленых и пирофитовых водорослей.
* (Есть указания на то, что пиреноиды играют также роль в синтезе жиров. Высказывалось предположение, что они являются носителями ферментов, способствующих синтезу различных питательных веществ.)
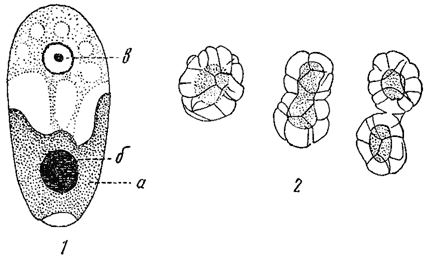
Рис. 20. Пиреноиды. 1 - отдельная клетка золотистой водоросли Hydrurus foetidus (с голым пиреноидом): а - хроматофор; б - пиреноид; в - ядро. 2 - деление скорлупчатого пиреноида зеленой водоросли Spirogyra Weberi
У большинства имеющих жгутики подвижных клеток (в том числе в зооспорах и гаметах), реже и у неподвижных клеток, чаще у их переднего конца, располагается, обычно в единственном числе, красный глазок или стигма (рис. 3, 19,1). Ее красная или коричневато-красная окраска обязана пигменту каротину. В ряде случаев установлена генетическая связь стигмы с пластидами: стигма образуется, благодаря накоплению каротина, из части пластиды или из целой пластиды и может рассматриваться как специализированная пластида. Однако в других случаях никакой связи между пластидами и стигмой констатировать не удается. Стигма в большинстве случаев состоит из двух частей: бесцветной и окрашенной. Первая имеет двояковыпуклую, чечевицеобразную форму и чаще обращена кнаружи; вторая - чашевидной формы, вогнутой стороной ориентирована к бесцветной части, содержит пигментированные зернышки и чаще располагается под ней (рис. 21).
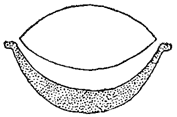
Рис. 21. Строение стигмы Volvox в поперечном разрезе (схема)
Функцией стигмы считается восприятие светового раздражения. Этому способствует вышеописанное ее строение, в котором бесцветная часть может играть роль линзы, концентрирующей световые лучи. Нужно, однако, заметить, что подвижные клетки, лишенные стигмы, также способны воспринимать световые раздражения.
Характерным элементом клеток многих водорослей - в течение всей их жизни или только на определенной стадии развития - являются органоиды движения - жгутики или жгуты. Они представляют собой тонкие цитоплазматические отростки в числе от 1 до многих и имеют довольно сложное и далеко не всегда одинаковое строение. Вдоль жгута, по-видимому, проходит эластичная осевая нить, окруженная, как чехлом, сократимой цитоплазмой. Иногда последняя не доходит до конца, и верхняя часть жгута состоит тогда только из осевой нити. В этих случаях она заметно тоньше нижней, как это имеет место у Volvocineae (рис. 22).
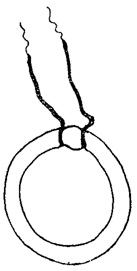
Рис. 22. Клетка Haematococcus с двумя жгутиками
Находящийся вне клетки отрезок жгута представляет собой лишь наружную часть ее жгутикового (локомоторного) аппарата. В цитоплазме жгут обычно связан с базальным зерном, располагающимся или близ поверхности клетки или глубже, около ядра. При наличии нескольких жгутов число базальных зерен или соответствует числу жгутов или меньше его. Последнее объясняется слиянием нескольких базальных зерен в одно более крупное образование - блефаропласт. Большой интерес представляет связь жгутикового аппарата с ядром, с несомненностью установленная для многих форм, у которых базальные зерна соединяются с ядром с помощью особой нити - ризопласта (рис. 23). Жгуты могут образоваться заново или делиться вместе с делением клетки.
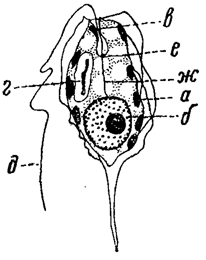
Рис. 23. Эвгленовая водоросль Phacus caudata. а - хроматофор; б - ядро; е - стигма; г - парамилоновое зерно; д - жгут; е - базальное зерно; ж - ризопласт
На других частях клетки (хондриосомы, некоторые скелетные образования и др.) мы останавливаться здесь не будем.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'