20. Отдел 11. Харовые водоросли - Charophyta (Г. М. Паламарь-Мордвинцева)
Харовые водоросли, харофиты, или лучицы, объединяют около 300 видов макроскопических водорослей, 50 из которых обнаружено в СССР. Харовые - наиболее крупные из пресноводных водорослей. Длина их таллома составляет обычно 20-30 см, но может достигать и 1-2 м. Все части их тела, включая органы размножения, хорошо различимы невооруженным глазом.
Индивиды харовых водорослей имеют довольно сложно и своеобразно устроенное тело. Наибольшего своеобразия достигает у них строение органов полового размножения, не известное более ни в одной другой группе растений.
Слоевища харовых водорослей имеют вид кустовидно ветвящихся нитевидных или стеблевидных зеленых членисто-мутовчатых побегов, укрепляющихся на дне водоемов с помощью многочисленных тонких бесцветных ризоидов (рис. 20.1). На основных побегах располагаются мутовки коротких боковых побегов, также членистого строения. Обе эти вегетативные части харовых водорослей внешне похожи на органы высших растений, и в научной литературе их условно называют стеблями (основные ветвящиеся побеги) и листьями (расположенные мутовками боковые побеги). Рост стеблей верхушечный, неограниченный, листья обладают предельным ростом. Места расположения мутовок называют узлами, а участки стебля между ними - междоузлиями. Листья могут быть осевыми, расположенными в одну линию, с "листочками" в узлах, и вильчато разделенными, когда на вершине первого членика образуются 2-4 членика второго порядка, и так до 3 раз. Между клетками узлов и междоузлий имеется существенное различие. Каждое междоузлие - это многоядерная гигантская, длиной в несколько сантиметров, вытянутая клетка, неспособная к делению (у некоторых харовых она покрыта еще корой), тогда как каждый узел состоит из собранных в диск нескольких мелких одноядерных клеток, дифференцирующихся в процессе деления и образующих как боковые ветви стебля, так и мутовку листьев.
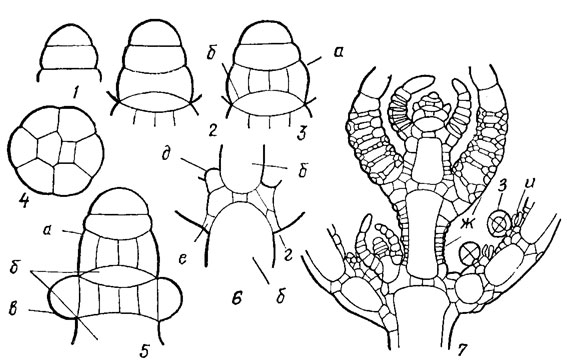
Рис. 20.1. Строение верхушки стебля Charophyta, продольный разрез: 1-6 - начальные стадии верхушечного роста (1 - двухклеточная стадия, 2 - четырехклеточная, 3 - начало формирования стеблевого узла и междоузлия, 4 - поперечный срез сформированного стеблевого узла, 5 - начальная стадия образования листьев из периферических клеток стеблевого узла, 6 - стеблевой узел с развитыми листьями и образующейся ветвью); 7 - часть стебля Chara sp. на протяжении верхних четырех мутовок; а - стеблевой узел; б - междоузлие, в - начальные листовые бугорки; г - базальный листовой узел; д - начальный бугорок боковой ветви стебля; е - клетки, образующие прилистники; ж - клетки стеблевой и листовой коры; з - антеридии, и - оогонии
Сложность строения таллома харовых водорослей определяется также дифференциацией клеток в процессе верхушечного роста основного побега (стебля). Верхушечная клетка стебля имеет характерную плоско-выпуклую форму. В течение вегетационного периода верхушечная клетка непрерывно делится горизонтальной (перпендикулярной длиной оси стебля) перегородкой на 2 клетки, из которой верхняя остается верхушечной, полусферической, а нижняя - цилиндрической (рис. 20.2, 1, 2). При следующем делений этих двух клеток такими же горизонтальными перегородками получается комплекс из четырех клеток, расположенных друг за другом, который является исходным для всего последующего процесса дифференциации и роста таллома. Новое деление верхушечной клетки повторяет описанный процесс возникновения цилиндрической клетки, а прежняя цилиндрическая клетка делится на верхнюю двояковогнутую и нижнюю двояковыпуклую клетки. Дальнейшая роль этих двух клеток различна: верхняя делится и становится исходной для формирования многоклеточного узла, а нижняя только растет в длину, образуя междоузлие. Деление исходной узловой клетки происходит не горизонтальными, а вертикальными (параллельными оси стебля) перегородками так, что после ряда делений образуются 2 клетки в центре узла и несколько клеток по его периферии (см. рис. 20.2, 3, 4). В дальнейшем центральные клетки не делятся, а периферические продолжают делиться и дают начало листьям и боковым ветвям стебля. Из каждой периферической клетки стеблевого узла возникает по одному листу.
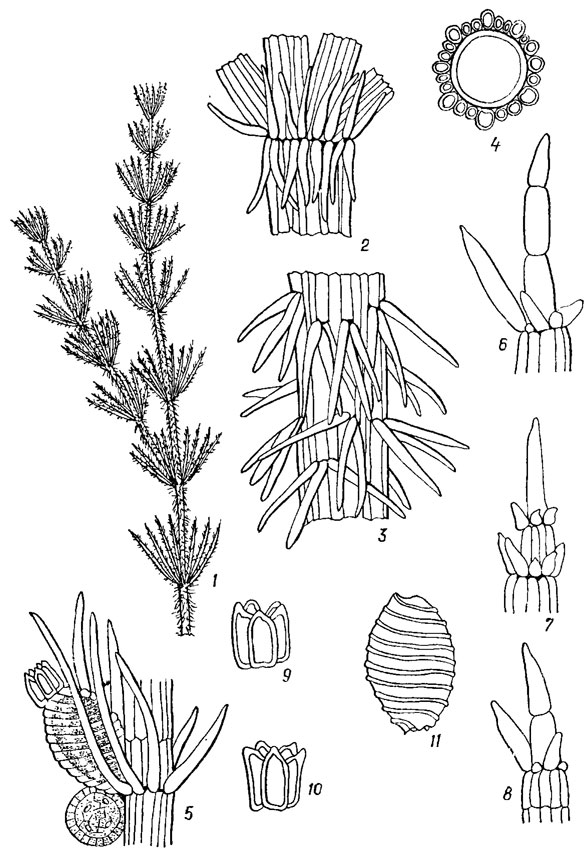
Рис. 20.2. Chara strigosa A. Br.: 1 - общий вид части растения; 2 - основные мутовки; 3 - часть стебля с коровыми шипами; 4 - стебель в поперечном разрезе; 5 - плодоносящий листовой узел; 6-8 - концы листьев; 9, 10 - коронки; 11 - ооспора
Листья закладываются первоначально как бугорки (см. рис. 20.2, 5), которые постепенно вытягиваются и развиваются в листья. При этом периферические узловые клетки стебля выполняют роль верхушечных ростовых клеток, подобным верхушечным клеткам стебля. Делятся они не беспредельно, а отчленяют от себя только определенное количество сегментов. Они, дифференцируясь, образуют членистые листья, состоящие из длинных неделящихся клеток и чередующихся с ними узлов, сложенных несколькими мелкими клетками, способными к дальнейшим делениям. Один или несколько конечных члеников листа лишены узлов. По строению листья сходны со стеблем, но функция узлов у них другая. Из периферических клеток большинства листовых узлов образуются органы полового размножения и окружающие их придаточные клетки. Только самый нижний, т. н. базальный узел, находящийся в основании каждого листа (см. рис. 20.2, 6) имеет другую функцию. Отдельные его клетки, расположенные в пазухах листьев, дают начало боковым ветвям стебля, из других образуются т. н. прилистники и кора.
У всех видов рода Chara Vaill. и у представителей некоторых других родов харофитов на стеблях при основании листовых мутовок имеется венчик более или менее вытянутых и заостренных длинных или почти шаровидных прилистников. Они образуются из двух боковых клеток базального узла каждого листа независимо от того, имеется у данного вида стеблевая кора или нет. Существует несколько вариантов строения прилистников, характерных для разных видов. Если делится одна из двух клеток, то образуется одноклеточный венчик прилистников (редкий случай). Если развиваются обе клетки по обе стороны листа, то венчик становится двукратным (частый случай). В обоих этих случаях возникает лишь один ряд прилистников, т. е. венчик может быть однократно однорядным или однократно двурядным. Чаще встречается третий случай, когда каждая начальная клетка делится дважды и развивается еще второй нижний ряд прилистников. Тогда венчик оказывается двукратно двурядным.
Междоузлия на стеблях многих харофитов не остаются голыми клетками, как у видов рода Nitella Ag., а обрастают слоем специальных клеток, получивших название коры. Стеблевая кора лучше всего развита у видов рода Chara. Она всегда построена из полос клеток, расположенных параллельными рядами вдоль междоузлия и слегка наискось. Эти полосы бывают первичными и вторичными. Количество первичных коровых полос соответствует количеству листьев в мутовке, что позволяет понять их происхождение. От основания каждого листа отходят 2 первичные коровые полосы: одна вниз по стеблю, другая - вверх. Их образуют 2 периферические клетки базального листового узла, расположенные посередине: одна под листом, другая над ним. Каждая из этих клеток выполняет функцию верхушечной ростовой клетки и развивается так же, как и при росте стебля. В итоге каждая первичная коровая полоса состоит из верхушечной клетки и чередующихся узлов и междоузлий. Узловые клетки небольшие, округлые, а клетки междоузлий крупные, вытянутые, более или менее цилиндрические. Когда встречные клетки смыкаются, рост первичных коровых полос заканчивается. Волнистая линия смыкания коровых полос бывает хорошо заметна посередине стеблевых междоузлий. Кора, составленная только из первичных коровых полос, называется однополосной. Она встречается у небольшой группы видов рода Chara.
Узловые клетки первичных коровых полос способны делиться в двух направлениях. Если они делятся параллельно поверхности стебля, то образуются шипы, одиночные или по 2-5 в пучках, направленных почти перпендикулярно к стеблю. Боковое направление делений узловой клетки приводит к появлению по обе стороны ее двух боковых клеток, которые в процессе разрастания вдоль стебля смыкаются и слагаются в полосы, названные вторичными коровыми полосами. Здесь возможны два случая. У одних видов рода Chara вторичные коровые клетки сравнительно короткие, составляющие около половины длины клеток коровых междоузлий. Они образуют лишь одну вторичную коровую полосу между двумя первичными. Такую кору называют двухполосной. Если же вторичные коровые клетки по длине равны клеткам междоузлий первичных коровых полос, то тогда происходит смыкание концов однозначных клеток, образованных от узлов одной и той же первичной коровой полосы, по обе ее стороны. В силу того что такие две вторичные коровые полосы (правую и левую) образует каждая первичная коровая полоса, по их боковым сторонам вдоль всего стеблевого междоузлия происходит смыкание двух параллельно идущих вторичных полос, происходящих от двух соседних первичных полос. Кору такого строения, когда между двумя первичными коровыми полосами находятся две вторичные (на одну первичную приходится две вторичные), называют трехполосной. Вторичные коровые полосы отличаются от первичных отсутствием округлых узловых клеток. На узловых клетках двух- и трехполосной коры, как и в первом случае, могут образовываться шипы (см. рис. 20.1, 2-5). Видов рода Chara с двух- и трехполосной корой значительно больше, чем с однополосной. Лишенных коры видов в роде Chara очень мало.
На листьях харовых кора образуется сходным образом, однако строение коровых полос очень упрощено. Листовая кора может быть только в том случае, если стебель покрыт корой. У разных видов рода Chara встречаются разные варианты развития коры.
Все клетки харовых водорослей в начальных стадиях дифференциации частей таллома одноядерны. При размножении этих клеток их ядро делится митотически. При последующей дифференциации, когда определенные клетки вытягиваются и становятся междоузлиями или другими удлиненными частями таллома, в них происходят многократные амитотические деления ядра без образования клеточных перегородок, в результате чего клетки становятся многоядерными, а ядра - вытянутыми, неправильной формы. Вытянутые, сформировавшиеся клетки харофитов имеют характерное строение. Их оболочка плотная, толстая. Внутренний слой образован целлюлозой; наружный слой состоит из кал лозы, в которой может отлагаться углекислая известь. Всю центральную часть клетки занимает центральная вакуоль с клеточным соком, а протоплазма распределена по периферии клетки. В более толстом внутреннем слое протоплазмы, граничащем с вакуолью, расположены многочисленные ядра, в тонком, примыкающем к оболочке, слое сосредоточены многочисленные хлоропласты. Они имеют форму мелких дисковидных телец, подобных хлорофилльным зернам высших растений. Хлоропласты распределяются равномерно, окрашивая всю клетку в ярко-зеленый цвет. Исключение составляет только узкая бесцветная полоса, идущая вдоль всей клетки чуть наискось - она лишена хлоропластов, но определяет их положение в остальных частях клетки: хлоропласты расположены ровными рядами параллельно этой бесцветной полосе. При достаточно большом увеличении в живых клетках видно стремительное движение протоплазмы по всей клетке, параллельно той же продольной линии. Движение протоплазмы у харовых водорослей отличается наибольшей скоростью среди всех растений: 1,5-2 мм/мин.
Набор пигментов сходен с таковыми у зеленых водорослей (хлорофиллы а и в и почти те же каротиноиды), а в качестве продуктов ассимиляции также вырабатывается крахмал.
Для харовых водорослей характерно половое и бесполое размножение. Вегетативное размножение происходит путем укоренения и последующего разрастания частей основных побегов или с помощью особых одноклеточных или многоклеточных клубеньков. Той же цели служат особые отростки, или вторичные побеги, вырастающие из старых листовых узлов и клубеньков. Половое размножение происходит с помощью половых органов: антеридиев и оогониев. Антеридии и оогонии образуются из особых клеток, отчленяемых некоторыми периферическими клетками листовых узлов. Они могут возникать или на одном талломе (однодомные виды), или на разных талломах (двудомные виды). Подробное описание органов размножения и его прохождения дано в разделе 2.2.
Харовые водоросли - очень древние растения. Их окаменевшие (фоссилизированные) оогонии и ооспоры, называемые "гирогониты", известны из силлурийских пластов мезозойской эры; обильны они в девоне. Однако вопрос о происхождении харовых водорослей не может быть разрешен на основании изучения ископаемых форм. Сравнительно-морфологический анализ также не вносит ясность в этот вопрос, так как строение вегетативных и генеративных органов харовых не имеет аналогий в других растительных группах. Только сходство с зелеными водорослями в наборе основных пигментов и мутовчатое строение побегов позволяет предполагать, что харовые водоросли произошли от каких-то высокоорганизованных мутовчатых зеленых водорослей, сходных с современными хетофоровыми из рода Draparnaldia Bory и особенно Draparnaldiopsis.
Ископаемые харовые очень многообразны. Большинство альгологов принимают классификацию харовых, согласно которой в пределах отдела Charophyta выделено несколько классов ископаемых форм. Все современные формы входят в один из этих классов - Charophyceae, в пределах которого выделен только один порядок Charales [81].
Харовые водоросли заселяют преимущественно пресные водоемы разного типа, особенно часто пруды и озера, но встречаются и во временных небольших водоемах: ямах, канавах, реже в ручьях реках. Есть виды, населяющие солоноватые водоемы. В типично морских условиях харовые не встречаются. Чаще они поселяются на мягких иловых грунтах, реже на песке на глубине 1-5 м, иногда до 40 м. В большинстве случаев они образуют обширные заросли. Все харовые водоросли предпочитают чистую спокойную воду. Для многих из них необходимо повышенное содержание в воде извести. Пределы pH составляют 5,2-9,8. Наибольшее число харовых найдено в водоемах с умеренной температурой воды, хотя некоторые встречаются и в ледяной воде на севере Норвегии и в горячих источниках Исландии и Йеллоустонского парка (США). Преимущественно это равнинные растения, но нередко обильно населяют водоемы предгорий и гор. Предельная высота, где они были найдены - 4766 м над уровнем моря в Гималаях.
Харовые водоросли распространены по всему земному шару, в водоемах всех континентов, кроме Антарктиды и островов. Однако из 300 известных видов харовых только 6 найдены на всех континентах, большинство же видов имеют ограниченное распространение.
В пределах СССР наиболее богаты харовыми водоемы европейской части СССР и Средней Азии (свыше 50 видов), значительно меньше их было найдено на Кавказе, в Западной и Восточной Сибири, менее всего - на Дальнем Востоке и в арктических районах.
Роль в природе харовых водорослей сравнительно невелика, однако там, где они поселяются, проявляется их влияние на гидрологический режим и биологические особенности водоемов. Режим водоема становится более устойчивым и в нем формируется особый биоценоз. На талломах харовых развивается множество эпифитов - микроскопических водорослей и бактерий, служащих кормом для беспозвоночных, поедаемых рыбой. В густых зарослях этих водорослей находят приют и защиту молодь рыб и мелкие животные. Замечено, что в водоемах с обильным развитием харовых водорослей отсутствуют или слабо развиты личинки комаров. Предполагают, что это обусловлено действием антибиотиков, выделяемых харовыми водорослями.
Харовые служат источником пищи для водоплавающих птиц, особенно на путях их осенних перелетов. Птицы используют главным образом ооспоры, заполненные крахмалом и каплями жира.
Хозяйственное значение харовых определяется их использованием в качестве удобрения тяжелых почв (Швейцария) благодаря обилию в них извести. Иногда при больших естественных скоплениях и образовании отложений они образуют лечебные грязи. Их также используют для очистки тяжелых органических жидкостей (например, при сахароварении). Очень широкое применение харовые водоросли получили как удобный объект для физиологических и биофизических исследований. Огромные размеры клеток, составляющие междоузлия, позволяют изучать такие явления, как проницаемость цитоплазматических мембран, закономерности движения цитоплазмы, биоэлектрические потенциалы клетки и т. п.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'