19.4.4. Порядок 4. Десмидиевые - Desmidiales
Известно более 4000 видов порядка Desmidiales. Индивиды характеризуются удивительным разнообразием очертаний, красотой форм и замечательной симметрией. Причудливость очертаний клеток в сочетании с разнообразной скульптурой и зелеными хлоропластами издавна привлекали к себе внимание не только профессиональных исследователей, но и любителей, благодаря чему эти водоросли являются наиболее хорошо изученными во всем мире.
Типичная клетка десмидиевых водорослей состоит из двух симметричных полуклеток, каждая из которых является зеркальным отражением другой (рис. 19.42). Благодаря своеобразному способу деления клеток у десмидиевых, которое будет описано ниже, одна полуклетка всегда старше другой. Обе полу клетки соединены между собой более узкой частью - перешейком. Угол, возникающий между двумя полуклетками, в результате сжатия или сужения клеток, называют синусом. Он может быть узким и длинным, закрытым или открытым, с краями, расходящимися наружу от острой или закругленной верхушки.

Рис. 19.42. Типичная клетка Desmidiales: 1 - вид спереди; 2 - вид сбоку; 3 - вид сверху (а - перешеек, б - синус)
Внешний вид клеток десмидиевых в большинстве случаев зависит от того, с какой стороны их рассматривают под микроскопом. Различают вид спереди (с широкой лицевой стороны), вид сбоку и вид сверху или снизу (см. рис. 19.42). Таким образом, клетки десмидиевых имеют три плоскости симметрии. Разнообразие очертаний клетки при взгляде с различных сторон имеет большое значение в систематике десмидиевых. Некоторые виды имеют очень сходные очертания при рассмотрении их спереди, но при этом резко отличаются друг от друга при рассмотрении сбоку или сверху. Однако клетки некоторых видов из родов Docidium Breb., Actinotaenium (Näg.) Teil. сверху округлые и, следовательно, имеют одинаковые очертания при любых поворотах вокруг продольной оси (рис. 19.43). Своеобразным строением отличаются клетки видов рода Closterium Nitzsch. (рис. 19.44, 3-5). Они обычно сужены к концам, не перешнурованы и имеют более или менее серповидную форму, в результате чего образуются неодинаковые спинной и брюшной края. Спинной край - более или менее выпуклый, а брюшной - вогнутый или почти прямой. Однако ясная симметрия полуклеток и в этом случае сохраняется.
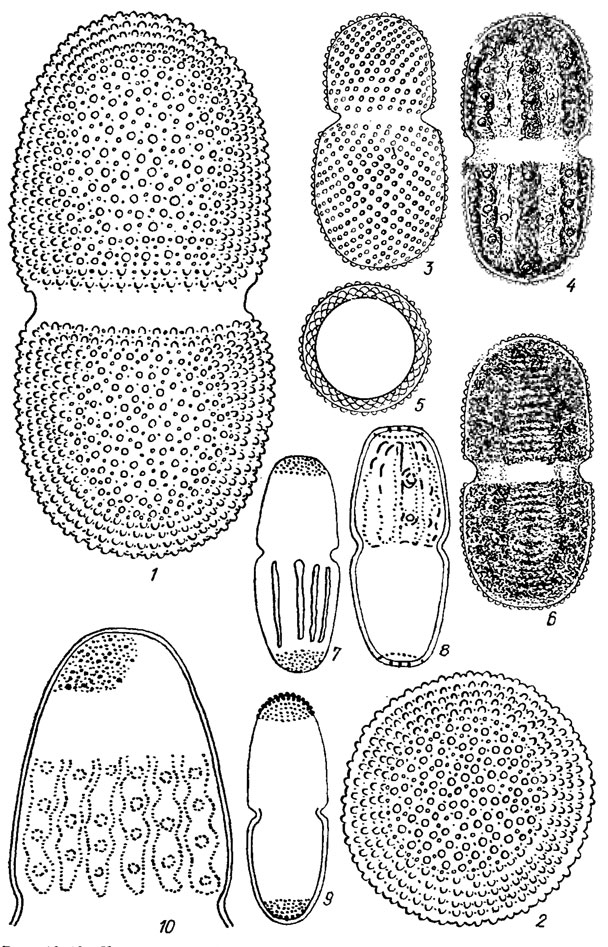
Рис. 19.43. Клетки Desmidiales (род Actinotaenium): 1-6 - A. tesellatum (Delp.) Pal.-Mordv.; 7-9 - A. clevei (Lund.) Teil.; 10 - A. turgidum (Breb.) Teil
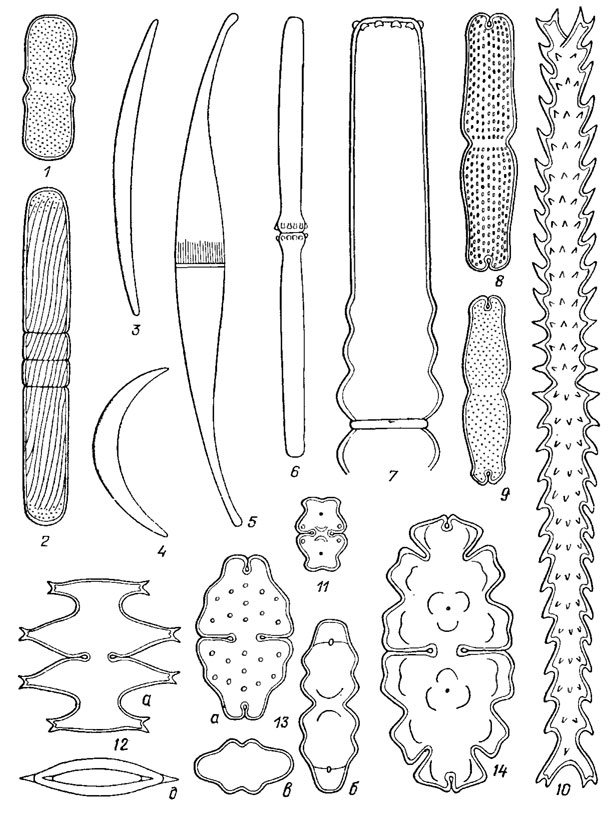
Рис. 19.44. Разнообразные формы клеток Desmidiales: 1 - Penium rufescens Cleve; 2 - P. spirostriolatum Barker; 3 - Closterium cornu Ehr., 4 - С. venus Kütz.; 5 - C. rostratum Ehr.; 6 - Docidiura baculum Breb.; 7 - Pleurotaeniura coronaturn (Breb.) Rabh.; 8 - Tetmemorus brebissonii (Menegh.) Ralfs; 9 - T. laevis (Kütz.) Ralfs; 10 - Triploceras gracile Bail.; 11 - Euastrura validutn W. et G. S. West; 12, 13 - E. sinuosum var. aboense Elfv.; 14 - E. oblongum (Grev.) Ralfs; a - вид спереди; б - вид сбоку; в, д - вид сверху
Перешнурованные виды десмидиевых, к которым относится большинство, характеризуются огромным разнообразием формы клеток (рис. 19.45). У одних видов клетки сильно вытянутые, цилиндрические, в поперечном сечении округленные или звездчатые; у других - сильно сжатые и плоские. У некоторых представителей развиваются особые отростки и шипы на углах полуклеток. Одним из наиболее характерных признаков десмидиевых является разнообразие очертаний клеток. Они бывают цилиндрическими, почти круглыми, эллиптическими, угловатыми (четырех-, шести-, восьмиугольными). Края клеток часто разделены на дольки или сегменты, или сильно изрезаны. Наиболее сложными очертаниями характеризуются клетки видов родов Micrasterias Ag., Euastrum Ehr. (см. рис. 19.44, 11-14) и Staurastrum Meyen (см. рис. 19.45, 9).
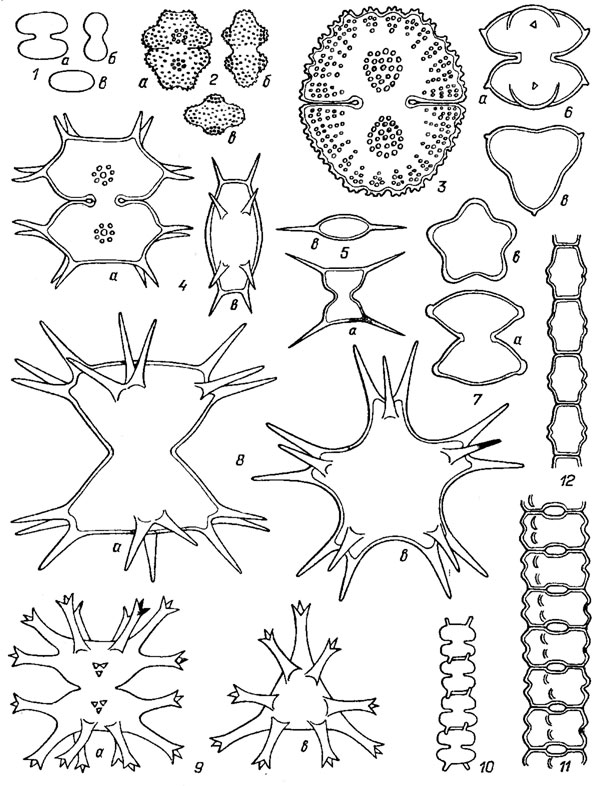
Рис. 19.45. Перешнурованные клетки Desmidiales: 1 - Cosmarium bioculatum Breb.; 2 - С. protractum (Näg.) De Bary; 3 - C. subquasillus Boldt; 4 - Xanthidium antilopaeum (Breb.) Kütz.; 5 - Staurodesraus incus (Breb.) Teil.; 6 - S. brevispinus (Brev.) Teil.; 7 - S. pachyrhynchus (Nordst.) Teil.; 8 - Raphidiastrum brasiliense var lundellii (W. West) Pal.-Mordv.; 9 - Staurastrum tohopekaligense var. trifurcatum W. et G. S. West; 10 - Teilingia filiforme (Ehr.) Bourr.; 11 - Desmidium aptogonum Breb ; 12 - Bambusina brebissonii Kütz.; a - вид спереди; 6 - вид сбоку; в - вид сверху
Оболочка клеток десмидиевых состоит из трех слоев. Самый внешний слой кажется под электронным микроскопом то петлеобразным, то гладким, или имеет вид пчелиных сот. Химическая природа этого слоя еще не известна. Предварительные исследования показывают, что он содержит пектинообразное вещество. У молодых клеток внешний слой оболочки тонкий, у старых достигает значительной толщины. За внешним слоем следует первичная оболочка, затем вторичная - самый внутренний слой. Первичная оболочка состоит из тончайших микрофибрилл, имеющих вид пучков соломы, которые перекрещиваются друг с другом. Вторичная оболочка состоит из более широких фибриллярных лент, расположенных параллельно микрофибриллам первичной оболочки.
Строение оболочки десмидиевых усложняется наличием круглых или овальных отверстий - пор. Они имеют разное назначение: 1) для выделения экскреторной слизи, которая постоянно окружает клетки десмидиевых; 2) для удаления из клетки продуктов жизнедеятельности; 3) для поступления питательных веществ из окружающей среды в клетку, так как внешний слой оболочки сам по себе непроницаем. Расположение пор в оболочке у разных видов десмидиевых чрезвычайно разнообразно (рис. 19.46). У некоторых поры очень мелкие и многочисленные, сосредоточены в определенных местах или равномерно распределены по всей оболочке. У других поры крупные и немногочисленные, располагаются в определенных местах, чаще на концах клеток. У третьих - имеются и мелкие, и крупные поры, расположенные различным образом. Поры часто имеют сложное строение.

Рис. 19.46. Расположение пор в оболочке у разных видов Desmidiales: 1 - Penium spirostriolatum Barker; 2 - Closterium dianae Ehr.; 3 - С. angustatum Kütz.; 4 - С. ralfsii Breb.; 5 - Docidium baculum Breb.; 6 - Pleurotaenium kayei (Arch.) Rabenh.; 7 - P. trabecula (Ehr.) Näg.; 8 - Triploceras gracile Bail.; 9 - Tetmemorus granulatus (Breb.) Ralfs.; 10 - Euastrum crassum (Breb.) Kütz. f. scrobiculatum Lund.; 11-12 - Cosmarium turgidum Breb.; 13 - С. praegrande Lund.; 14 - С. obsoletum Reinsch.; 15 - Xanthidum armatum (Breb.) Reiisch.; 16 - Staurastrum jaculiferum W. West; 17-19 - Cosmocladium saxonicum De Bary; 20, 21 - Oocardium stratum Nag
Известно два типа строения порового аппарата десмидиевых - тип Closterium и тип Cosmarium. Поровый аппарат типа Closterium более примитивен (рис. 19.47). Он состоит из порового канала обратноворонкообразной формы, узкая часть которого, являющаяся собственно порой, расположена во внешнем слое оболочки, а более широкая часть проходит через первичную и вторичную оболочки. Фибриллы этих оболочек пронизывают часть канала, закрывая его своеобразным ситом, через которое может проходить клеточное вещество и выбрасываться наружу через узкую часть. Поровый аппарат типа Cosmarium характеризуется наличием свободного сквозного канала, обычно снабженного различными приспособлениями (см. рис. 19.47). Самое сложное строение порового аппарата типа Cosmarium встречается в роде Micrasterias. Здесь настоящий поровый канал снабжен особыми поровыми нитями. Снизу он начинается на внутренней границе вторичной оболочки поровой луковичкой, проходит через первичную оболочку и внешний слой и заканчивается под внешним слоем конечными запонками, Поровые нити - трубкоподобные образования, которые расширяются наружу в бокаловидные конечные запонки. Поровые луковички имеют вид коротеньких гвоздиков с широкой шляпкой, которые погружены своим острием в поровый канал: они сидят в поровом канале, как затвор умывальника в отверстии для стока. Это приспособление в поровом аппарате имеет большое значение. С его помощью поры по мере надобности могут то открываться, то закрываться. В данном случае поровый механизм служит не только для выделения продуктов обмена и поступления питательных веществ, но и предохраняет клетку от нежелательной потери клеточного вещества, а также проникновения в клетку вредных веществ снаружи.
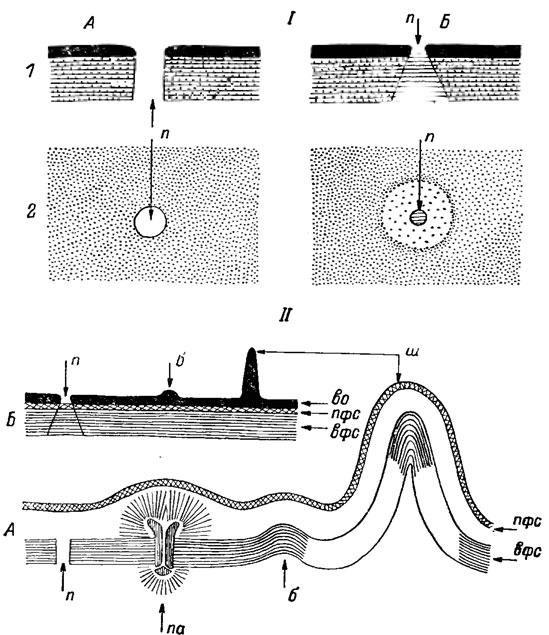
Рис. 19.47. Строение оболочки и пор у Desmidiales: I - общая схема: 1 - на срезе, 2 - в плане; II - детали строения на срезе; А - тип Cosmarium; Б - тип Closterium; п - пора; б - бородавки; ш - шипы; во - внешняя оболочка; пфс - первичный фибриллярный слой внутренней оболочки; вфс - вторичный фибриллярный слои внутренней оболочки; па - поровый аппарат
Кроме пор структура внешней оболочки десмидиевых может быть усложнена наличием особых скульптурных украшений: гранул, ямок, бородавок и шипов, в большинстве случаев расположенных в определенном порядке. Они, часто в сочетании с особыми выпуклостями, буграми и отростками оболочки, создают удивительные узоры, которые придают особую красоту и разнообразие десмидиевым. Эти скульптурные украшения являются важными систематическими признаками таксонов порядка десмидиевых.
Внешний слой оболочки у многих десмидиевых бывает пропитан соединениями железа, отложения которых располагаются у разных видов по-разному, что определяет их принадлежность к тому или иному роду. Железо часто откладывается в местах скульптурных украшений и особенно часто в районе перешейка (рис. 19.48). У отдельных видов железа откладывается так много, что оболочка их клеток приобретает желтый, ржавый или темно-коричневый цвет. Способность клеток десмидиевых к накоплению железа считается надежным систематическим признаком, однако у некоторых родов оно до сих пор не обнаружено.

Рис. 19.48. Отложения железа в оболочках клеток Desmidiales: 1 - Closterium navicula (Breb.) Lutk.; 2 - Penium cylindrus (Ehr.) Breb.; 3 - C. abruptum W. West; 4 - C. dianae Ehr.; 5 - Cosmarium contractum Kirchn. var. ellipsoideum (Elfv.) W. et G. S. West; 6 - C. tinctum Ralfs var. intermedium Nordst.; 7 - C. quadrum Lund.; 8 - Xanthidium aculeatum Ehr.; 9 - Desmidium pseudostreptonema W. et G. S. West
Внешний слой оболочки благодаря присутствию пектина и наличию пор часто образует толстый слизистый покров, полностью окутывающий особь или (у нитчатых форм) - всю нить (рис. 19.49). С помощью этого слизистого покрова многие десмидиевые приклеиваются к высшим водным растениям или к другим водорослям, другие - увеличивают свою плавучесть.
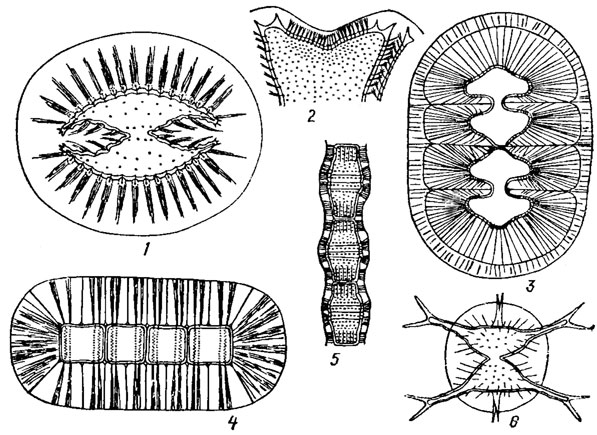
Рис. 19.49. Образование слизистого покрова у разных видов Desmidiales: 1 - S. sonthalianum Turn.; 2 - Micrasterias rotata (Grev.) Ralfs; 3 - Spondylosium nitens (Wallich.) Arch.; 4 - Hyalotheca mucosa (Mert.) Ehr.; 5 - Bambusina brebissonii Kütz.; 6 - Staurastrum pseudopelagicum W. et G. S. West
У некоторых представителей клетки объединены в нити посредством слизистых тяжей, проходящих между прилегающими верхушками смежных клеток (рис. 19.50). При этом связь клеток оказывается настолько крепкой, что нити разрываются намного легче в районе перешейка клетки, чем в точках верхушечного прикрепления. Такое склеивание посредством слизи иногда дополняется развитием верхушечных отростков оболочки клеток (Sphaerozosma Corda и Teilingia Bourr.; см. рис. 19.50, 5). У колониального рода Cosmocladium Breb. клетки удерживаются вместе с помощью слизистых стебельков, образуя небольшую разветвленную свободноплавающую колонию. У видов рода Oocardium Näg. колония имеет вид полушария, достигающего в диаметре 1-2 мм, прикрепляющегося к известковым скалам. Она состоит из более или менее параллельных, расходящихся лучами слизистых нитей значительной толщины. Каждая нить расширяется по направлению к поверхности колонии и иногда разветвляется. На свободном конце каждой слизистой нити расположена одна клетка. Она размещена так, что ее продольная ось лежит перпендикулярно оси слизистой нити. У некоторых видов образуется очень плотная слизь, которая окружает клетку в виде футляра и служит защитой при неблагоприятных условиях; она предохраняет клетку от недостатка влаги и защищает ее от высыхания.
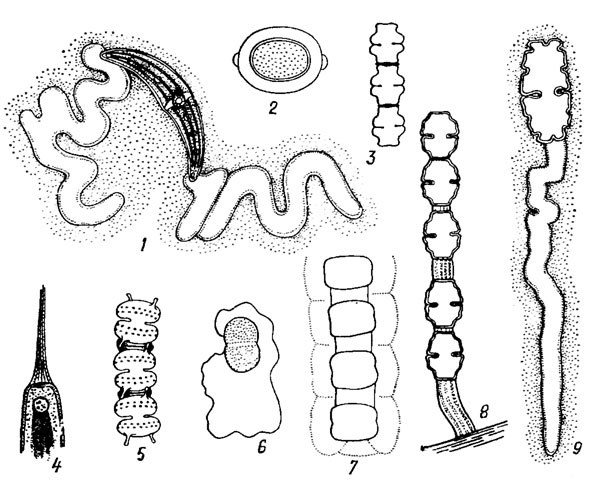
Рис. 19.50. Функции слизи у Desmidiales: 1 - Closterium sp.: 2 - Desmidium cylindricum Grev.; 3 - Spondyloslum pulchellum Arch.; 4 - Closterium moniliferum (Bory) Ehr.; 5 - Teilingia filiformis (Ehr.) Bourr.; 6 - Cosmarium pseudoconnatum Nordst.; 7 - Hyalotheca dtssiliens (Smith) Breb.; 8 - Cosmarium ruttneri Krieg.; 9 - Euastrum oblofigum (Gfev.) Ralfs
У десмидиевых имеется еще особая слизь, с помощью которой осуществляется движение клетки (см. рис. 19.50, 1, 9). Эта слизь выделяется через специальные поры, размещенные на концах клеток. Движение свойственно всем представителям десмидиевых, но особенно хорошо изучено оно у видов рода Closterium. Движение у них осуществляется таким образом. Один конец клетки клостериума временно прикрепляется к субстрату, а другой выполняет ритмические колебательные движения под углом 180°. Через некоторое время свободный конец клетки прикрепляется к субстрату, а колебательные движения начинает совершать первый конец. Так, прикрепляясь то одним, то другим концом, клетка клостериума передвигается по субстрату. При температуре воды 33°С смена позиции клетки происходит каждая 6-8 мин. У других родов также отмечены неправильные, порой спазматические движения, но иногда они имеют плавный характер.
Некоторые виды образуют слизистые "ножки", приподнимающие клетку над субстратом. Все десмидиевые положительно фототаксичны. В ясные солнечные дни они всегда собираются у поверхности воды. Если склянку с живыми десмидиевыми водорослями поставить на окно, то через некоторое время клетки сконцентрируются на освещенной стороне.
В вакуолях многих десмидиевых встречаются особые слизистые тельца в виде маленьких неподвижных зерен. Эти тельца состоят из органического вещества, так как при нагревании до 64°С, а также в кислотах и щелочах они растворяются. По-видимому, они представляют собой запасные питательные вещества, связанные каким-то образом с выделением слизи.
У десмидиевых хлоропласты бывают двух типов: осевые и пристенные. У большинства видов встречается осевой хлоропласт, причем почти всегда по одному в каждой полуклетке. У очень мелких форм рода Cosmarium Corda в центре каждой клетки имеется только один простой хлоропласт с одним пиреноидом. Хлоропласты десмидиевых имеют разнообразную форму: звездообразных тел (Penium Breb.), осевых конусов с радиальными пластинками (Closterium), стенкоположных лент (Pleurotaenium Näg.), осевых тел с радиальными лучами (Cosmarium) и др. (рис. 19.51; 19.52). Пристенные хлоропласты встречаются реже, чем осевые, и преимущественно у более крупных форм. У некоторых представителей особенно из родов Euastrum и Micrasterias, имеющих осевые хлоропласты, наблюдается тенденция к развитию и пристенных хлоропластов. Центральная часть хлоропласта становится у них более тонкой и исчезает совсем, тогда как периферическая часть развивается очень хорошо. Иногда цельные пластинки хлоропластов в периферической части клеток разделяются на дольки или многочисленные короткие отростки, направленные к оболочке клетки. Благодаря этому площадь хлоропласта значительно увеличивается.
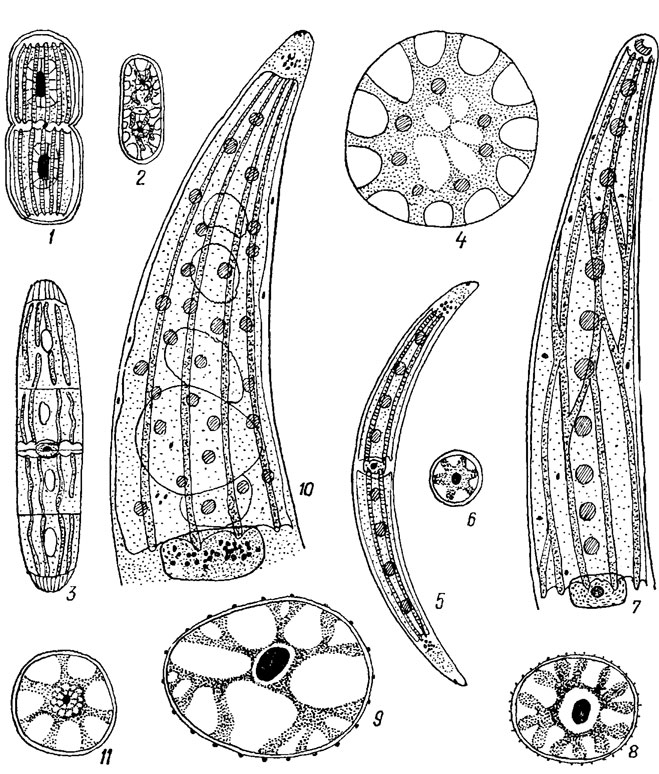
Рис. 19.51. Хлоропласта Desmidiales: 1 - Actinotaenium rufescens (Cleve) Teil.; 2 - Penium phymatosporum Nordst.; 3, 4 - P. spirostriolatum Barker (3 - общий вид, 4 - поперечный срез); 5, 6 - Closterium dianae Ehr. (5 - общий вид, 6 - поперечный срез); 7, 8 - С. striolatum Ehr. (7 - общий вид, 8 - поперечный срез); 9 - С. costatum Corda; 10, 11 - С. ehrenbergii Menegh. (10 - общий вид; 11 - поперечный срез)
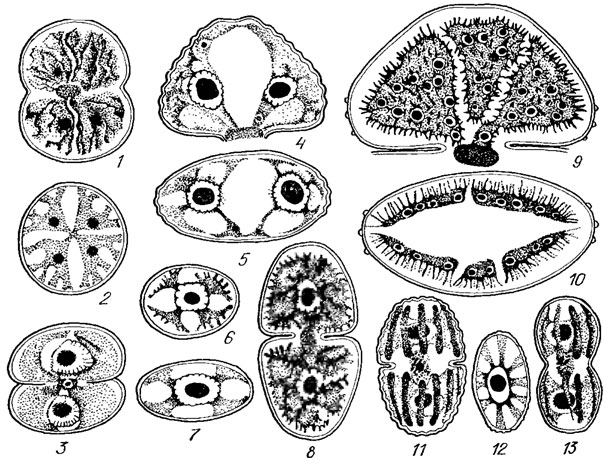
Рис. 19.52. Хлоропласты Desmidiales рода Cosmarium: 1, 2 - С. pseudoconnatum Nordst.; 3, 7 - С. depressum Lund.; 4, 5 - С. ochtodes Nordst.; 6, 8 - C. pseudopyramidatum Lund.; 9, 10 - C. askenasyi Schmidle; 11-13 - C. speciosum Lund
Размеры и характер расчленения хлоропласта могут быть различными у разных особей одного и того же вида в зависимости от условий их существования. При хорошем питании и освещении клеток хлоропласт хорошо развит и клетки окрашены в темно-зеленый цвет. В неблагоприятных условиях (при недостаточном количестве питательных веществ, отсутствии или избытке какого-либо вещества) хлоропласты развиваются слабо, преимущественно у стенок клетки. В таком случае клетки имеют бледно-зеленый или желтоватый цвет.
У разных представителей десмидиевых количество и положение пиреноидов сильно варьирует в зависимости от формы хлоропласта и условий существования клетки. У большинства видов, имеющих крупные пристенные хлоропласты, пиреноиды разбросаны без определенного порядка и порой малозаметны. В осевых хлоропластах пиреноиды обычно сосредотачиваются в наиболее массивной их части. Количество пиреноидов, внедренных в ось осевого хлоропласта, зависит от его величины. У некоторых крупных видов рода Micrasterias их может быть более 100, а у мелких видов обычно имеется только один пиреноид. В зависимости от массы накопленных веществ количество пиреноидов изменяется.
Деление клеток десмидиевых хорошо прослеживается на примере видов рода Cosmarium. Ядро, лежащее в районе перешейка, делится, образуя два, которые расходятся к центрам обеих полуклеток. После этого в середине перешейка образуется перегородка, тесно примыкающая своими краями к внутреннему слою оболочки (рис. 19.53, 1). Затем область перешейка вытягивается, в результате чего две старые полу клетки отделяются одна от другой (рис. 19.53, 2). Верхняя и нижняя части перешейка, лежащие по обе стороны поперечной перегородки, начинают раздуваться в виде пузырька, увеличиваться в размерах, после чего расщепляется сама перегородка (рис. 19.53, 3). Образовавшиеся из раздувшихся частей перешейка молодые полуклетки начинают быстро расти и достигают нормального размера и формы, оставаясь соединенными друг с другом своими вершинами (рис. 19.53, 5, 6). Когда клетки достигают зрелого возраста, их оболочка, являющая собой временное образование, отбрасывается и отпадает, наподобие линяющей кожи. Тогда клетки окончательно разъединяются и начинают самостоятельное существование (рис. 19. 53, 7). Таким образом в каждой клетке оказывается более старая и более молодая полуклетки. На ранних стадиях развития содержимое молодых полуклеток совершенно прозрачно, поэтому клеточное ядро, лежащее в области перешейка, хорошо видно без специального окрашивания. Потом протоплазма становится густозернистой, заполняется вакуолями. Наконец хлоропласты старых полуклеток удлиняются, делятся, и один из них переходит в молодую полу клетку. У неперешнурованных форм десмидиевых, например, у видов родов Closterium и Penium, клеточное деление происходит еще более сложным образом.
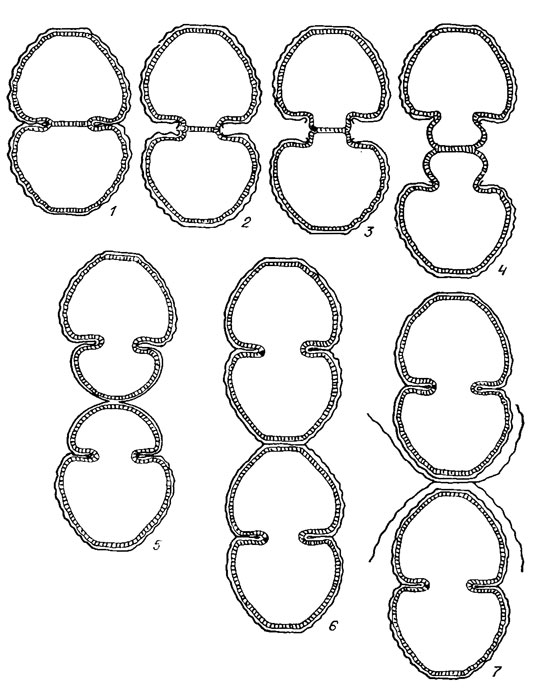
Рис. 19.53. Различные фазы деления клеток Desmidiales рода Cosmarium: 1 - возникновение поперечной перегородки; 2 - удлинение перешейка и начало развития молодых полуклеток; 3-5 - постепенный рост молодых полуклеток; 6, 7 - разделение вполне сформировавшихся молодых полу клеток, сопровождаемое сбрасыванием их временной оболочки
Половое размножение десмидиевых происходит так же, как и у других представителей класса Conjugatophyceae. Конъюгация десмидиевых совершается безжгутиковыми апланогаметами, которые выполняют функцию половых клеток. Гаметы двух клеток сливаются, образуя зиготу, которая со временем превращается в зигоспору. Зигоспоры известны у многих представителей десмидиевых, но подробности полового процесса были изучены только у нескольких видов. Это связано с тем, что половой процесс у этих водорослей довольно редко встречается в природе. Из зигоспоры развиваются 2, 4 или 1 проросток.
Порядок включает 3 семейства: Peniaceae (1 род), Closteriaceae (1 род), Desmidiaceae (более 30 родов).
Характерными представителями являются Penium spirostriolatum Bark., Р. cylindrus (Ehr.) Breb., Closterium acerosum (Schrank) Ehr.,C. leibleinii Kütz., C. ehrenbergii Menegh., Staurastrum gracile Ralfs, Xanthidium antilopaeum (Breb.) Kütz., Euastrum ansatum (Ehr.) Ralfs, Micrasterias rotata (Grev.) Ralfs, Cosmarium conspersum Ralfs, C. margaritatum (Lund) Roy et Biss.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'