18. Отдел 9. Бурые водоросли - Phaeophyta (Phaeophycophyta, Phaeophyceae) (Н. А. Мошкова)
Бурые водоросли - преимущественно морские многоклеточные растения, весьма крупные, сложно расчлененные, прикрепленные к субстрату. В настоящее время известно около 1500 видов бурых водорослей, относящихся к 240 родам. В пресных, большей частью холодных проточных водах умеренных широт до сих пор обнаружено 5 видов бурых водорослей. Благодаря незначительным размерам их талломов и редкой встречаемости, они остаются слабо изученной группой растений как в биологическом, так и в экологическом отношениях.
Общим внешним признаком индивидов бурых водорослей служит желтовато-бурая окраска их талломов, обусловленная наличием у них большого количества желтых и бурых пигментов. Талломы могут быть микроскопическими (несколько десятков микрометров) и гигантскими (30-50 м; у некоторых видов родов Laminaria Lamour., Macrocystis Ag., Sargassum Ag.). Форма талломов очень разнообразна: нитевидная, корковидная, мешковидная, пластинчатая (цельная или с разрывами, выростами и многочисленными отверстиями, гладкая или с продольными складками и ребрами), а также кустообразная.
Наиболее просто организованы талломы бурых водорослей порядка Ectocarpales. У примитивных организмов (Bodanella Zimmerm.) таллом представлен однорядными беспорядочно ветвящимися в одной плоскости нитями, плотно прилегающими к субстрату. Виды рода Ectocarpus Lyngb. имеют кустистые талломы, образованные однорядными восходящими обильно ветвящимися нитями, основанием которых служат стелющиеся ризоиды (рис. 18.1).
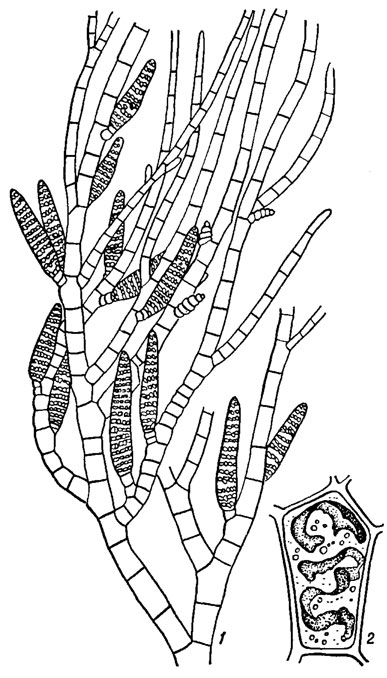
Рис. 18.1. Ectocarpus siliculosus (Dillw.) Lyngb.: 1 - фрагмент таллома с многогнездными спорангиями; 2 - клетка с лентовидными хлоропластами
У некоторых представителей порядка Chordariales восходящие нити соединены в пучки, заключенные в слизь. При этом различают одноосевой тип строения таллома, при котором от основания поднимается вверх одна нить, а от нее ответвляются другие нити, идущие рядом с ней, и многоосевой тип строения, когда от основания поднимается вверх сразу пучок однорядных нитей. У высокоорганизованных бурых водорослей (Laminaria, Fucus Tourn., Sargassum) талломы дифференцированы и напоминают цветковые растения. У них имеются стебле-, листо- и корневидные части, некоторые крупные представители имеют воздушные пузыри, удерживающие ветви в вертикальном положении.
Рост бурых водорослей интеркалярный или апикальный. У наиболее примитивных форм происходит интеркалярный диффузный рост, у более эволюционно продвинутых водорослей намечается уже интеркалярная зона роста. Она обычно размещается в базальной части многоклеточных волосков и обусловливает трихоталлический рост, характерный для бурых водорослей.
На поверхности однорядных талломов бурых водорослей образуются многоклеточные нитевидные волоски. При этом различают волоски настоящие и ложные. Настоящие волоски имеют при основании интеркалярную зону роста, где клетки делятся часто и потому они более мелкие, короткоцилиндрические или дисковидные. Ложные волоски такой специальной зоны роста не имеют и представляют собой продолжение вегетативных однорядных нитей с сильно вытянутыми клетками, лишенными хлоропластов.
В многорядных слоевищах бурых водорослей наблюдается специализация клеток с образованием тканей - паренхиматозный тип структуры тела. В простейшем случае различают кору из интенсивно окрашенных клеток, содержащих большое количество хлоропластов и особых вакуолей - физод, и сердцевину, состоящую из бесцветных, часто более крупных клеток одинаковой формы. У более сложно организованных бурых водорослей (Laminariaceae, Fucaceae) коровой слой достигает значительной толщины и состоит из интенсивно окрашенных клеток разного размера и формы (рис. 18.2). Поверхностные четыре слоя коры образованы мелкими клетками, вытянутыми по направлению к поверхности. Эти верхние слои называют меристодермой - делящейся покровной тканью. Они способны активно делиться и производить волоски и органы размножения. Настоящие волоски размещаются на поверхности меристодермы рассеянно или пучками и часто погружены своими основаниями в особые углубления - криптостомы. Глубже под меристодермой лежит кора из более крупных окрашенных клеток. В центральной бесцветной части таллома можно различать две группы клеток. В центре находятся рыхло- или плотнорасположенные нити с сильно вытянутыми клетками - сердцевина, между сердцевиной и корой лежат крупные бесцветные клетки - промежуточный слой. Сердцевина бурых водорослей служит не только для транспорта продуктов фотосинтеза, но и выполняет механическую функцию; в ней часто имеются тонкие нити с толстыми продольными оболочками. Наиболее сложным анатомическим строением отличаются представители порядка Laminariales, у которых в сердцевине развиваются слизистые каналы с особыми секреторными клетками для транспортирования продуктов фотосинтеза - сито- видные трубки и трубчатые нити.
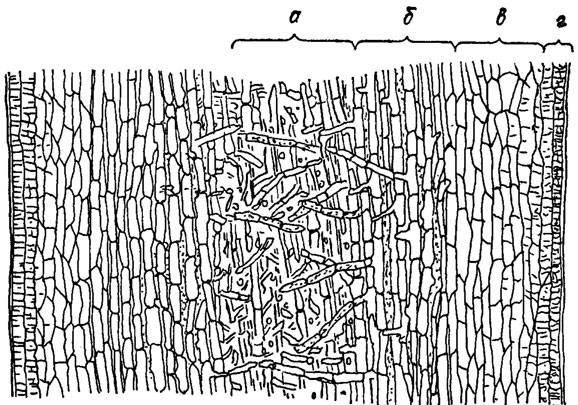
Рис. 18.2. Паренхиматозная структура бурых водорослей (продольный разрез таллома Laminaria sp.): а - сердцевина; б - промежуточный слой; в - кора; г - меристодерма
Талломы бурых водорослей прикреплены к грунту или другим субстратам и лишь иногда, вследствие механического повреждения, отрываются и свободно плавают. Органами прикрепления обычно являются длинные выросты - ризоиды, у крупных форм они массивные и представляют собой короткие корневидные выросты, охватывающие субстрат наподобие птичьих когтей. У представителей порядка Fucales и некоторых других водорослей органом прикрепления является дисковидное разрастание в основании таллома - базальный диск, уплощенный либо конический, плотно пристающий к грунту.
Ветвление бурых водорослей моноподиальное. Боковые веточки очередные, рассеянные или супротивные. При быстром дорастании их до размеров главной нити (материнских клеток) возникает дихотомовидное ветвление. Довольно часто очередные и супротивные ветви размещаются в одной плоскости и водоросли приобретают своеобразный перистый вид. Правильность размещения веточек нередко маскируется вторичными разветвлениями.
Среди бурых водорослей есть виды с эфемерными, однолетними и многолетними талломами. На продолжительность существования талломов большое влияние оказывают экологические условия. Многолетние талломы бурых водорослей бывают нескольких типов. У одних водорослей таллом многолетний, каждый год отмирают только побеги, на которых развивались органы размножения (Fucales), у других (Laminariales) многолетними являются ствол и органы прикрепления, пластинчатая часть - однолетняя. У некоторых тропических видов саргассовых водорослей многолетним бывает только диск, служащий для прикрепления таллома.
Клетки бурых водорослей одноядерные, шаровидные, эллипсоидные, боченковидные, большей частью цилиндрические, вытянутоцилиндрические или короткоцилиндрические, дисковидные, иногда многоугольные или неопределенных очертаний. Разнообразны они также по размерам. Ядро обычного для эукариот типа.
Оболочка клеток двуслойная. Внутренний слой целлюлозный, однако целлюлоза бурых водорослей по своим свойствам отличается от целлюлозы цветковых растений и потому ее иногда называют альгулезой. Внешний слой оболочки пектиновый, состоит обычно из белковых соединений альгиновой кислоты и ее солей. Благодаря такому строению оболочка бурых водорослей может сильно набухать, превращаясь в слизистую массу иногда значительного объема. У большинства бурых основой пектина является камедевидное вещество - альгин (растворимая натриевая соль альгиновой кислоты), у некоторых - фукоидин.
Содержимое соседних клеток бурых водорослей сообщается посредством плазмодесм. В клетках с толстыми оболочками (в крупных слоевищах) хорошо выражены поры.
Клетки бурых водорослей имеют одну крупную или несколько мелких вакуолей. Кроме того, встречаются физоды - очень мелкие вакуоли (до 4 мкм в диаметре), заполненные фукозаном - соединением, подобным танину. В молодых клетках физоды бесцветные, в старых - окрашены в желтый или бурый цвет.
Хлоропласты пристенные, большей частью многочисленные, мелкие, дисковидные, реже лентовидные или пластинчатые. Однако по мере старения клеток форма хлоропластов может изменяться и вместо узких лентовидно изогнутых в клетках могут появиться многочисленные дисковидные хлоропласты. Пиреноиды имеются или в хлоропластах вегетативных клеток или только в хлоропластах гамет; у ряда видов пиреноиды вообще отсутствуют или встречаются редко.
Бурые водоросли отличаются своеобразным сложным набором пигментов. В хлоропластах обнаружены хлорофиллы а, с (хлорофилл b отсутствует), β- и ε-каротины, а также несколько ксантофиллов-фукоксантин, виолаксантин, антераксантин, зеаксантин и др. Из них особенно специфичен фукоксантин интенсивно бурого цвета. Различные соотношения этих пигментов обусловливают окраску бурых водорослей от оливково-желтоватого до темно-бурого, почти черного цвета.
Продуктами ассимиляции бурых водорослей являются разные растворимые в клеточном соке углеводы - ламинарии (полисахарид), маннит (шестиатомный спирт, играющий существенную роль в обмене веществ), а также масло.
У бурых водорослей встречаются бесполое и половое формы размножения. Однако вегетативное размножение фрагментацией таллома нельзя считать безусловным. Оно наблюдается лишь тогда, когда оторванные талломы попадают в более или менее защищенные места и продолжают там вегетацию. При этом их нижние более старые части отмирают, разрушаются, а молодые ветви развиваются в самостоятельные растения, не прикрепленные, однако, к грунту. Такие растения, плавающие или лежащие на грунте, никогда не образуют органов полового и бесполого размножения.
Специальные почки вегетативного размножения имеются лишь у видов рода Sphacelaria Lyngb. (рис. 18.3).
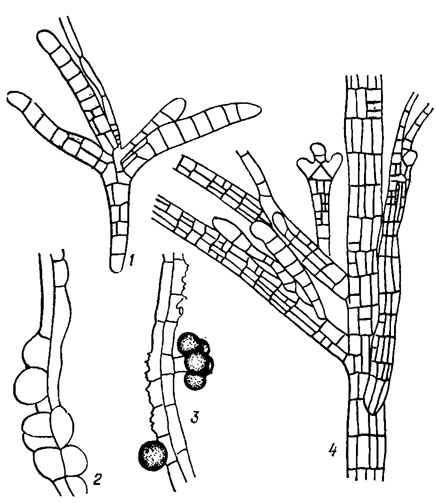
Рис. 18.3. Почки вегетативного размножения видов рода Sphacelaria Lyngb.: 1, 4 - Sphacelaria cirrhosa (Roth) Ag. (1 - почка вегетативного размножения, 4 - фрагмент слоевища с почками вегетативного размножения на разных стадиях развития); 2, 3 - Sphacelaria fluviatllis Jao (почки вегетативного размножения на разных стадиях развития)
Бесполое размножение осуществляется подвижными зооспорами, образующимися помногу в одногнездных спорангиях. У наиболее просто организованных морских и пресноводных бурых водорослей (Ectocarpus, Sphacelaria, Pleurocladia А. Br. и др.) одногнездные спорангии представляют собой шаровидные или эллипсоидные клетки, которые располагаются как боковые выросты ветвей (рис. 18.4, 1). В спорангиях происходит редукционное деление ядра с последующими многократными митотическими делениями; одновременно с ядрами делятся хлоропласты. В результате образуется большое количество зооспор, которые освобождаются через разрыв оболочки на вершине спорангия и, недолго поплавав, прорастают в новое, такое же по внешности, но уже гаплоидное растение. У видов рода Laminaria зооспорангии образуют сорусы на поверхности листовидной пластины. Сорус состоит из парафиз и зооспорангиев (см. рис. 18.4, 2, 5). Парафизы представляют собой вытянутые клетки, с хлоропластами в верхнем расширенном конце, развивающиеся на поверхности таллома между органами размножения и служащие для их защиты. Оболочка парафиз на вершине сильно ослизняется, образуя своеобразный толстый слизистый колпачок. Слизистые колпачки соседних парафиз смыкаются, благодаря чему получается сплошной толстый слой слизи, защищающий сорус. Зооспорангии вытянутоэллипсоидные, с ослизненной оболочкой на вершине. В зооспорангии развивается в зависимости от вида по 16-128 зооспор. Первое деление ядра редукционное. Некоторые бурые водоросли размножаются неподвижными, лишенными жгутиков, спорами - апланоспорами. Моноспоры наблюдаются только у видов порядка Tilopteridales, тетраспоры - у видов порядка Dictyotales (Dictyota dichotoma (Huds.) Lamour., см. рис. 18.4, 4).
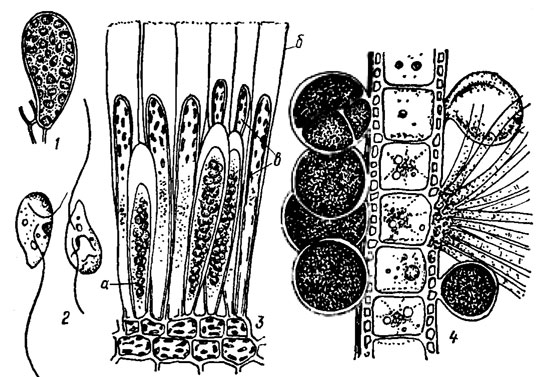
Рис. 18.4. Спорангии бурых водорослей: 1 - одногнездный зооспорангий Pleurocladia lacustrls A. Bf.; 2 - зооспоры; 3 - группа зооспорангиев (сорус) Laminarla sp. (а - зооспорангии; б - сильноослизненная оболочка парафиз, в - парафизы); 4 - тетраспорангии Djctyota djchotoma (Huds.) Lam
Половой процесс изо-, гетеро- и оогамный. Гаметы обычно образуются в многогнездных гаметангиях, по одной в каждой камере. Подвижные клетки бурых водорослей - зооспоры в гаметы имеют сходное строение - они грушевидные, с одним хлоропластом и двумя жгутиками, прикрепленными сбоку. Один жгутик более длинный, перистый, направлен вперед, другой - более короткий, гладкий, бичевидный, направлен назад. Стигма у подвижных клеток заметна не всегда. Хлоропласт мужских гамет при оогамии может быть бесцветным.
В цикле развития большинства бурых водорослей класса Phaeozoosporophyceae происходит смена форм развития и чередование полового и бесполого поколений, т. е. гаметофита (иногда еще и гаметоспорофита, если один и тот же организм может давать начало зооспорам и гаметам) и спорофита.
Эти процессы подробно описаны в разделе 3.2.3. Здесь остановимся лишь на некоторых особенностях циклов развития бурых водорослей. У наиболее примитивных морских бурых водорослей порядка Ectocarpales наблюдается изоморфная смена форм развития, однако строгое чередование поколений еще отсутствует. Из спор, образующихся спорофитом, могут развиваться как гаметофиты, так и спорофиты.
Правильная изоморфная смена форм развития наблюдается у представителей порядка Dictyotales. Наиболее широко распространенной из них является Dictyota dichotoma (Huds.) Lam., обладающая вильчато разветвленным талломом с плоскими, обычно расположенными в одной плоскости ветвями без продольного ребра (рис. 18.5).
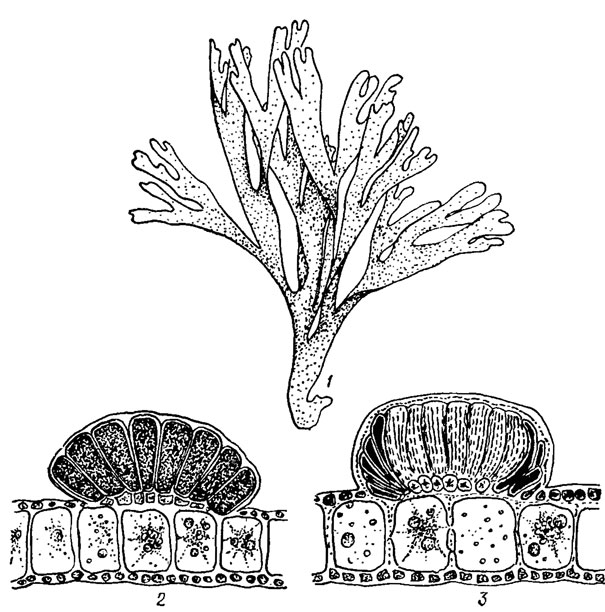
Рис. 18.5. Dictyota dichotoma (Huds.) Lamour.: 1 - общий вид таллома; 2 - продольный разрез женского гаметангия; 3 - продольный разрез мужского гаметангия
Гетероморфную смену форм развития с обязательным чередованием спорофитов и гаметофитов имеют водоросли порядка Laminariales. Цикл развития их характеризуется правильной сменой мощного спорофита и микроскопического, просто устроенного гаметофита.
К бурым водорослям, не имеющим смены форм развития, а лишь смену ядерных фаз, относятся представители семейств Fucaceae, Cystoseiraceae и Sargassaceae. Нормальное воспроизведение их возможно только половым путем. Половой процесс - типичная оогамия. Половые органы развиваются в концептакулах (рис. 18.6). Из стенки концептакула вырастают длинные волоски - парафизы, заполняющие почти всю его полость. Особенно длинные волоски развиваются в женских концептакулах, где они в виде пучка выдаются из отверстия концептакула. Среди этих волосков развиваются оогонии и антеридии (рис. 18.7, 1-5). Антеридии в большом количестве образуются на концах особых однорядных разветвленных веточек, вырастающих из стенки концептакула. В их оболочке различимы два слоя. При созревании антеридия лопается его внешняя оболочка и антерозоиды выходят наружу в виде пакета, окруженного внутренней оболочкой. В морской воде внутренняя оболочка разрывается и антерозоиды грушевидной формы с крупным ядром и оранжевой стигмой, освобождаются. Оогонии шаровидные или эллипсоидные, снабженные трехслойной оболочкой, располагаются в концептакулах на короткой одноклеточной ножке. В оогонии образуется 8 яйцеклеток, они выходят в воду, окруженные двумя внутренними слоями оболочки оогония. Когда яйцеклетки полностью освобождаются от оболочек оогония, происходит оплодотворение. Оплодотворенная яйцеклетка вырабатывает собственную толстую оболочку и тотчас же начинает прорастать, образуя новое слоевище фукуса.
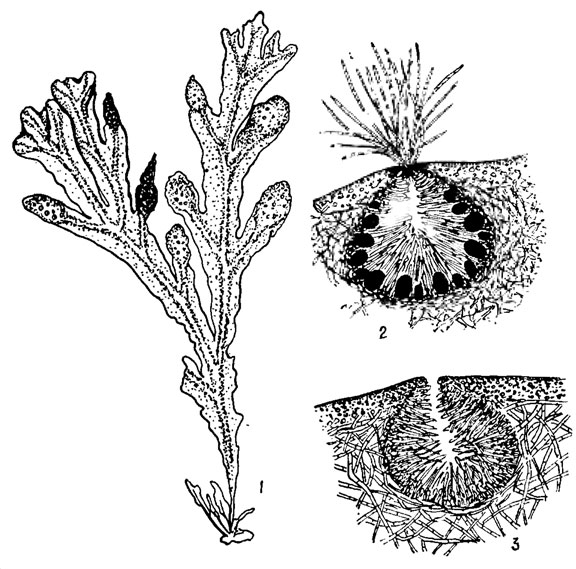
Рис. 18.6. Fucus vesiculosus L.: 1 - общий вид таллома; 2 - зрелый женский концептакул; 3 - зрелый мужской концептакул
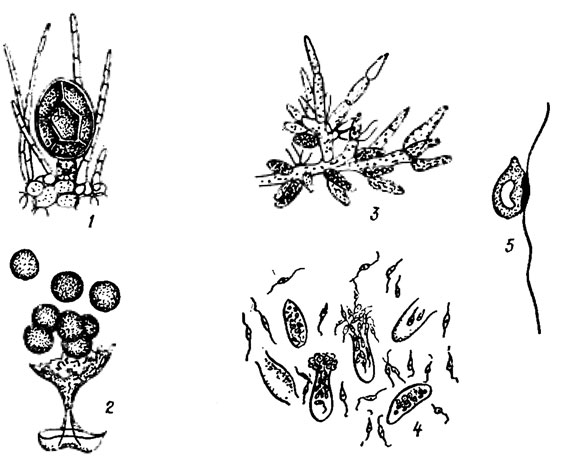
Рис. 18.7. Структуры Fucus vesiculosus L., связанные с размножением: 1 - оогоний; 2 - выход яйцеклеток из оогония; 3 - веточка с антеридиями; 4 - выход антерозоидов из антеридиев; 5 - антрозоид
У пресноводных бурых водорослей циклы развития не исследованы.
Во взглядах на классификацию бурых водорослей существуют некоторые расхождения. По мнению ряда исследователей [122, 252, 563, 578 и др.], отдел Phaeophyta делится на 2 класса: Phaeozoosporophyceae и Cyclosporophyceae. К циклоспоровым принадлежат бурые водоросли, у которых органы размножения развиваются в концептакулах и имеют крупные размеры, позволяющие видеть их на препаратах невооруженным глазом. К феозооспоровым относят все остальные бурые водоросли, многие из которых размножаются зооспорами. С 30-х годов появилась тенденция классифицировать бурые водоросли в зависимости от особенностей циклов развития. Тогда же было предложено разделить бурые водоросли на 3 класса: Isogenerate, Heterogenerate, Cyclosporae [519,571]. Предложенная классификация получила весьма широкое распространение [416, 417, 655 и др.]. Однако деление бурых водорослей на изогенератные и гетерогенератные довольно условно, поскольку в обоих классах в отдельных порядках существуют представители с противоположным типом смены форм развития. Придерживаясь взглядов отечественных альгологов, мы принимаем классификационную схему деления бурых водорослей на 2 класса - Phaeozoosporophyceae и Cyclosporophyceae.
Вопрос о происхождении бурых водорослей еще слабо разработан. А. Шерфелл [619] связывал их происхождение с золотистыми (Chrysophyta). По мнению А. Пашера [575], существует филогенетическая связь бурых с криптофитовыми (Cryptophyta). Своеобразное строение жгутиков вместе с бурой окраской позволили М. Шадефо [371] объединить в один большой отдел Chromophycophyta такие крупные таксоны, как Pyrrhophyta (куда он, кроме перидиней, включил криптофитовые и эвгленовые водоросли), Chrysophyta (к которым он отнес, кроме золотистых, желтозеленые и диатомовые водоросли) и Phaeophyta. По биохимическим свойствам из всех буроокрашенных организмов к бурым водорослям ближе всего диатомовые. Именно диатомовые и бурые водоросли характеризуются такими общими пигментами, как хлорофилле (свойствен также и перидинеям), фукоксантин (встречается и у золотистых) и неофукоксантины А и В. Принимая во внимание наличие ряда сходных черт между диатомовыми, золотистыми и бурыми водорослями, мы присоединяемся к мысли, высказанной рядом ученых [120, 176, 177 и др.], о возможности происхождения их от близких, если не общих, монадных предков.
Согласно Г. Папенфуссу [572], исходным порядком бурых водорослей является Ectocarpales. Паренхиматозное строение таллома, апикальный рост, оогамный половой процесс и гетероморфная смена форм развития в разных группах бурых водорослей развивались независимо друг от друга.
Морские бурые водоросли широко распространены во всех морях земного шара. Их заросли обычны в прибрежных водах Антарктиды и северных островов Канадского Арктического архипелага. Наибольшего развития они достигают в морях умеренных и приполярных широт, где благодаря низкой температуре и повышенной концентрации биогенных веществ, создаются наиболее благоприятные условия для их вегетации. Бурые водоросли заселяют по вертикали все горизонты шельфа. Их заросли встречаются от литоральной зоны, где они во время отлива часами находятся вне воды, до глубины 40-100 (200) м. И все же наиболее густые и обширные заросли бурых водорослей образуются в верхней части сублиторали до глубины 6-15 м. В этих местах при достаточной освещенности существует постоянное движение воды, вызываемое прибоем и поверхностными течениями, которое обеспечивает, с одной стороны, интенсивное поступление к талломам биогенных веществ, а, с другой, - ограничивает поселение растительноядных животных.
Обычно бурые водоросли обитают на скалистых или каменистых грунтах, и только в спокойных местах у берегов или на большой глубине они могут удерживаться на створках крупных раковин моллюсков или на гравии. Оторванные талломы сносятся течением в спокойные места с илистым или песчаным дном, где они при достаточной освещенности продолжают вегетировать. Виды с воздушными пузырями на талломе при отрыве от грунта всплывают к поверхности воды, образуя большие скопления (Саргассово море). Среди морских бурых водорослей имеется значительное количество эпифитных и эндофитных форм.
В морях умеренных и приполярных широт наибольшего развития бурые водоросли достигают в летние месяцы, хотя бурный рост их талломов начинается уже ранней весной, когда температура воды приближается к 0°С. В тропических морях массовое развитие бурых приурочено к зимним месяцам, когда температура воды несколько понижается. Некоторые виды морских бурых водорослей могут встречаться в сильно опресненных участках морей с соленостью меньше 5‰.
Роль в природе бурых водорослей чрезвычайно велика. Они являются одним из основных источников органического вещества в прибрежной зоне, особенно в морях умеренных и приполярных широт, где их биомасса может достигать десятков килограммов на 1 м2. Кроме того, заросли бурых водорослей служат местом размножения, укрытия и питания многих прибрежных животных; они создают также условия для поселения микроскопических и макроскопических водорослей других систематических групп.
Велико и хозяйственное значение бурых водорослей, особенно в качестве сырья для получения различного рода веществ (например, альгинатов - солей альгиновой кислоты, в частности альгината натрия). Это вещество широко используют для стабилизации разнообразных растворов и суспензий. Добавление небольшого количества альгината натрия повышает качество пищевых продуктов (консервов, мороженого, фруктовых соков и т. д.), разнообразных красящих и клеящих веществ. Альгинаты применяют в книгопечатании, при производстве пластмасс, синтетических волокон и пластификаторов, для получения стойких к атмосферным воздействиям лакокрасочных покрытий и строительных материалов. Они входят в состав высококачественных смазочных материалов для машин, растворимых хирургических нитей, мазей и паст в фармацевтической и парфюмерной промышленности. В литейном производстве с помощью альгинатов улучшают качество формовочной земли. Альгинаты находят применение в производстве электродов для электросварки, позволяющих получать более высококачественные швы. Бурые водоросли используют также как сырье для получения маннита, применяемого в фармацевтической промышленности, в пищевой промышленности - для изготовления диабетических продуктов питания, а в химической - в производстве синтетических смол, красок, бумаги, взрывчатых веществ, при выделке кож. Бурые водоросли содержат большое количество иода и других микроэлементов, поэтому они идут на приготовление кормовой муки. В свежем и переработанном виде их применяют в качестве удобрений.
Издавна бурые водоросли используют в медицине. Сейчас выявляются все новые направления их применения, например, для изготовления заменителей крови, для получения препаратов, предотвращающих свертывание крови и способствующих выведению радиоактивных веществ из организма. С давних времен бурые водоросли (преимущественно представители порядка Laminariales) употребляются человеком в пищу.
К негативным свойствам бурых водорослей относится их участие вместе с другими организмами в обрастании судов, буев, а также различных гидротехнических сооружений, погруженных в воду, что ухудшает их эксплуатационные показатели.
Интенсивное использование дикорастущих морских макрофитов, в частности бурых водорослей, привело к исчерпанию их природных запасов и поставило человечество перед необходимостью их искусственного выращивания. Поэтому в последние 30 лет значительное развитие получила аквакультура водорослей. В Норвегии и Великобритании не только успешно культивируют виды рода Laminaria, но и усовершенствуют технологию их производства [363]. Во Франции проводят работы по акклиматизации представителей рода Macrocystis [336]. Интенсивно развивается аквакультура морских водорослей в США. При этом особое внимание уделяется Macrocystis pyrifera [699]. В СССР проводят исследования по искусственному разведению Laminaria saccharina (L.) Lam. в Белом море [186]. Таким образом, выращивание морских водорослей приобретает промышленный характер и становится все более рентабельной отраслью растениеводства, несмотря на некоторые трудности экономического и экологического плана [293].
В пресных водах умеренных широт обнаружено 5 видов бурых водорослей из класса Phaeozoosporophyceae: Bodanella lauterbornii Zimmerm. (порядок Ectocarpales, семейство Ectocarpaceae) (рис. 18.8, 1), Pleurocladia lacustris A. Br. (порядок Chordariales, семейство Myrionemataceae) (рис. 18.8, 2). Heribaudiella fluviatilis (Aresch.) Sved. (порядок Chordariales, семейство Lithodermataceae (рис. 18.8, 3)), Streblonema longiseta Arnoldi (порядок Chordariales, семейство Streblonemataceae) (рис. 18.8, 4). Sphacelaria fluviatilis Jao (порядок Sphacelariales, семейство Sphacelariaceae) (рис. 18.8, 5).
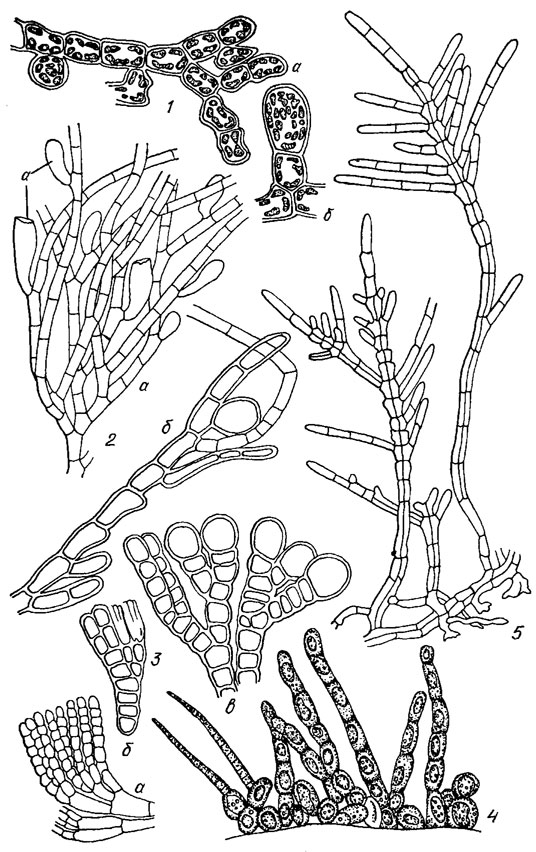
Рис. 18.8. Пресноводные бурые водоросли порядков Ectocarpales (1), Chordariales (2-4) и Sphacelariales (5): 1 - Bodanella lauterbornii Zimmerm. (а - фрагмент таллома; б - одногнездный спорангий); 2 - Pleurocladia lacustris A. Br. (а, б - фрагменты таллома с одногнездными спорангиями); 3 - Heribaudiella fluviatilis (Aresch.) Sved. (а - восходящие нити, вырастающие из стелющихся по субстрату, б, в - нити с одногнездными спорангиями); 4 - Streblonema longiseta Arnoldi, общий вид таллома; 5 - Sphacelaria fluviatilis Jao, общий вид таллома
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'