17. Отдел 8. Красные водоросли - Rhodophyta (Rhodophycophyta, Rhodophyceae) (Н. А. Мошкова)
Красные водоросли, или багрянки, - преимущественно морские растения. Это самая обширная группа растений, насчитывающая около 4000 видов, относящихся более чем к 600 родам. В пресных и солоноватых континентальных водоемах, а также в аэрофитных условиях обитает немногим более 50 видов.
Морские багрянки в подавляющем большинстве многоклеточные крупные индивиды, сложного морфологического и анатомического строения, достигающие в длину от нескольких сантиметров до метра. Окраска талломов разнообразна - от розового до темно-красного, почти черного цвета, часто с синевато-фиолетовым оттенком, в некоторых случаях голубоватого, голубовато-зеленого, иногда желтого цвета. Среди морских багрянок имеется также много мелких микроскопических форм, но лишь очень немногие, наиболее примитивные из них, имеют одноклеточные или колониальные талломы.
Форма тела красных водорослей весьма разнообразна. Здесь можно встретить нитевидные и пластинчатые, цилиндрические и корковидные, пузыревидные и кораллоподобные формы, в разной мере рассеченные и разветвленные. Нитевидные талломы могут быть очень тонкими, волосовидными, состоящими из однорядных нитей, либо более грубыми, состоящими из нескольких рядов клеток. Цилиндрические талломы плотные или с полостью внутри, слизистые, хрящеватые или кожистые, в виде шнуров или обильно разветвленных кустиков. Большого разнообразия достигают пластинчатые формы: встречаются цельные пластины, сложно рассеченные, с дополнительными выростами по краю и по поверхности таллома. Мелкие бородавчатые выросты называют папиллами, более крупные, напоминающие по форме основные ветви, - пролификациями. Некоторые пластинчатые талломы имеют листовидную форму, пронизаны "нервами" и "ребрами", напоминающими жилкование листьев цветковых растений.
Пресноводные багрянки преимущественно мелкие, нередко микроскопические водоросли, одноклеточные, колониальные или нитчатые, однорядные, простые или разветвленные; макроскопические формы - многорядно нитевидные, кустиковидные, шнуровидные или щетинковидные, высотой от нескольких сантиметров до нескольких дециметров. Встречаются также талломы в виде небольших, тонких корочек или пленочек, прижатых к субстрату. Пресноводные багрянки преимущественно оливково-зеленого, зеленовато-голубого или серовато-стального цвета.
Все многообразие внешних форм красных водорослей можно свести к нескольким основным, отчетливо выраженным, морфологическим структурам вегетативного тела.
Коккоидный тип структуры наблюдается у наиболее примитивных пресноводных багрянок класса Bangiophyceae. В виде отдельных свободно существующих шаровидных клеток встречаются Cyanidium caldarium (Tild.) Geitl. (рис. 17.1) и частично Porphyridium aeriigineum Geitl.
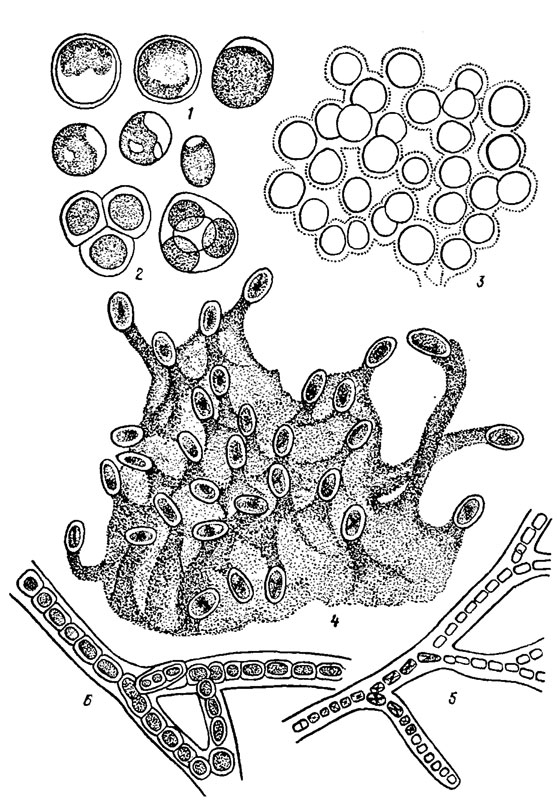
Рис. 17.1. Красные водоросли с коккоидной и пальмеллоидной структурой таллома: 1, 2 - Cyanidium caldarium (Tild.) Geitl. (1 - группа клеток, 2 - автоспоры); 3 - фрагмент таллома Porphyridium purpureum (Bory) Drew et Ross; 4 - фрагмент таллома Chroothece mobilis Pasch. et Popova; 5 - фрагмент таллома Chroodactylon wolleanum Hansg.; 6 - фрагмент таллома Goniotrichum elegans (Chauv.) Zanard
Усложненный вариант коккоидной структуры, проявляющийся в образовании большого количества слизи, объединяющей в колонии неподвижные, непосредственно не связанные между собой клетки. Пальмеллоидная структура [78] также свойственна примитивным, преимущественно пресноводным бангиевым водорослям. Если слизь выделяется всей поверхностью клетки, то возникают слизистые скопления клеток (см. рис. 17.1, 5), или образуются пленки неопределенных очертаний (виды родов Porphyridium Näg., Flintiella Ott., Rhodospora Geitl.). Если слизь выделяется лишь отдельным участком оболочки клетки (Chroothece Hansg.), возникают своеобразные слизистые ножки, на которых и размещаются клетки (см. рис. 17.1, 4). У колониальных форм пресноводных бангиевых водорослей наблюдается тенденция к нитевидному росту талломов. Клетки, заключенные в общую довольно толстую слизистую обвертку, не плотно соединены друг с другом и располагаются преимущественно в один, местами в два ряда. Нитевидные колонии Chroodactylon Hansg. и Goniotrichum Kütz. могут быть простыми или ветвиться по типу ложного ветвления (см. рис. 17.1, 5, 6).
Нитчатая (трихальная) структура встречается преимущественно как среди морских, так и пресноводных представителей класса Bangiophyсеае. В простейшем случае таллом состоит из однорядной, местами двурядной нити, все клетки которой, кроме базальной, морфологически одинаковы и способны к делению преимущественно в одной, реже в двух плоскостях (Erythrotrichia Aresch.; рис. 17.2, 1-5). Дальнейшее усложнение строения вертикальной нити заключается в приобретении клетками способности делиться в трех взаимно перпендикулярных плоскостях, в результате чего однорядная нить преобразуется в многорядную (Bangia Lyngb.).
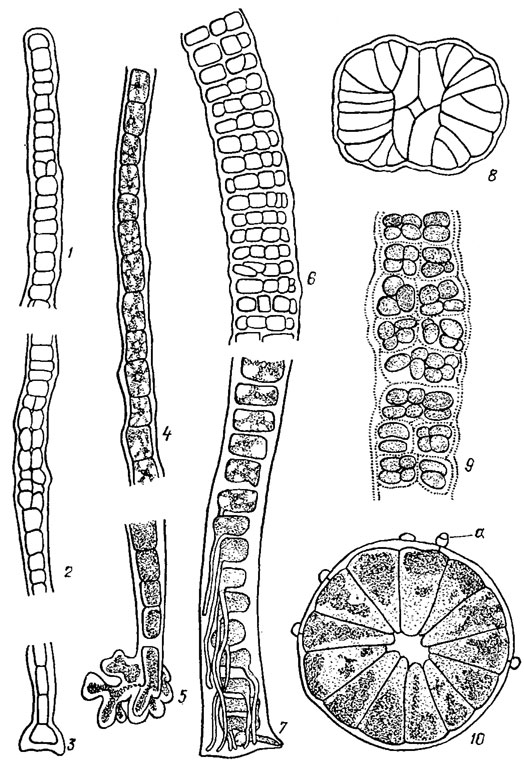
Рис. 17.2. Красные водоросли с нитчатой структурой таллома: 1-5 - Erythrotrichia carnea (Dillw.) J. Ag. (1 - верхушка нити, 2, 4 - средние части нитей, 3, 5 - базальные части нитей без ризоидов и с ризоидами); 6-10 - Bangla atropurpurea (Roth) Ag. (6 - фрагмент средней части многорядной нити; 7 - базальная часть однорядной нити с ризоидными выростами, 8-10 - многорядные части талломов в поперечном сечении, видны спермации - а)
Вместе с усложнением строения вертикальной нити, усложняется и строение органа прикрепления - у базальной клетки появляются ризоидные выросты. У более продвинутых форм ризоидные выросты отходят не только от базальной, но и от соседних с ней клеток (см. рис. 17.2, 7). Что касается представителей класса Florideophyceae, то нитчатая структура свойственна лишь немногим примитивным видам порядка Nemaliales. Это преимущественно мелкие организмы, живущие обычно, на других водорослях и животных.
Пластинчатая структура* таллома встречается преимущественно у морских водорослей класса. Bangiophyceae и, особенно, класса Florideophyceae. У наиболее примитивных бангиевых она представлена однослойными микроскопическими многоклеточными пластинками, состоящими из однорядных разветвленных в одной плоскости нитей, тесно сросшихся друг с другом боковыми поверхностями (Erythrocladia Rosenv.). У более совершенных бангиевых - видов рода Porphyra Ag. - таллом в виде довольно крупной однослойной или двуслойной пластины паренхиматозной структуры с ровными или складчатыми краями (рис. 17.3, 1-3). В нижней части пластина сужается в маленький стебелек, переходящий в подошву, образованную многочисленными ризоидами. У флоридеевых водорослей крупные пластинчатые талломы сложного анатомического строения псевдопаренхиматозной структуры свойственны преимущественно представителям семейств Phyllophoraceae, Rhodymeniaceae, Delesseriaceae (см. рис. 17.3, 4, 5).
* (Строение тела особи дано по Голлербаху (1977).)
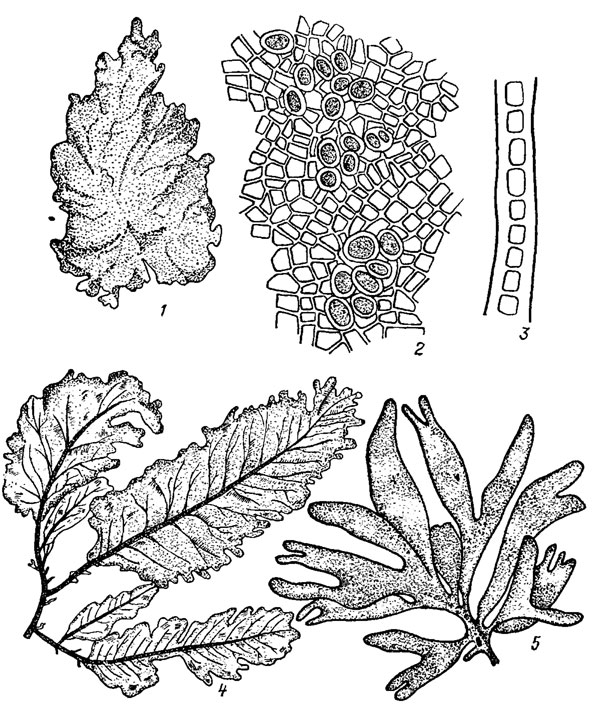
Рис. 17.3. Красные водоросли с пластинчатой структурой таллома: 1-3 - Porphyra Ieucosticta Thur. (1 - общий вид; 2 - вид сверху, заметны карпоспоры; 3 - в поперечном разрезе); 4 - фрагмент таллома Delesseria sanguinea Lamour.; 5 - фрагмент таллома Chondrus Stack
Разнонитчатая (гетеротрихальная) структура лежит в основе строения многих сложно организованных бангиевых и флоридеевых водорослей. В простейшем случае гетеротрихальный таллом у бангиевых состоит из однорядных вертикальных свободных нитей (скудно или обильно разветвленных) и горизонтальных, стелющихся по субстрату, обычно срастающихся в плотный диск или пластину неопределенных очертаний (Kylinielia Skuja; рис. 17.4, 1). У бангиевых водорослей из рода Compsopogon Mont, таллом кустиковидный, основные (главные) нити состоят из крупных бесцветных клеток, окруженных однослойной или многослойной мелкоклеточной пигментированной корой. Слоевище прикрепляется к субстрату подошвой, а также ризоидными выростами (см. рис. 17.4, 2, 3).
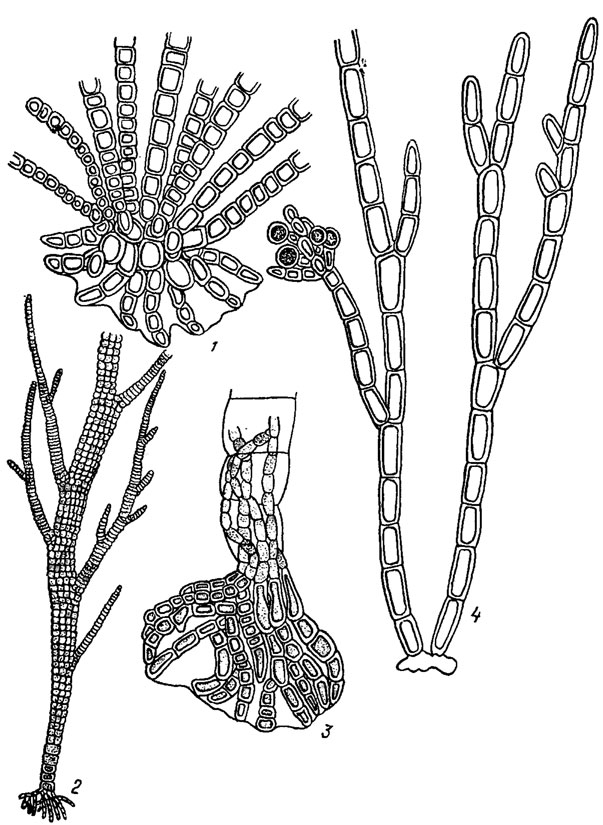
Рис. 17.4. Красные водоросли с разнонитчатой структурой таллома: 1 - общий вид таллома Kyliniella latvica Skuja; 2, 3 - Compsopogon chalybeus Kütz. (2 - фрагмент таллома, 3 - базальная часть таллома с ризоидами, образующими подошву); 4 - фрагмент таллома Chantransia pygmaea Kütz
Дальнейшим усложнением строения однорядного нитчатого таллома флоридеевых водорослей явилось возникновение большого числа обильно разветвленных веточек ограниченного роста, отходящих супротивно или мутовчато от каждой клетки основной нити и растущих лишь до определенного предела. В отличие от них основные нити (ветви), растущие в длину в течение всего вегетационного периода, называют нитями (ветвями) неограниченного роста. Талломы багрянок с большим количеством веточек ограниченного роста обильно пропитываются слизистой субстанцией, скрепляющей все разветвления, и принимают вид слизистых шнуров, простых или разветвленных.
При дальнейшем усложнении строения талломов, веточки ограниченного роста ветвятся и разрастаются так, что смыкаются друг с другом, образуя вокруг центральной части плотный периферический слой - кору. Сложная анатомическая структура многих морских багрянок создается за счет развития особых ризоидных нитей, или гиф, имеющих вид узких клеток с толстой оболочкой, которые тянутся вдоль осевых нитей. По мере усложнения организации талломов степень развития ризоидов увеличивается, нередко они развиваются настолько обильно, что заполняют всю внутреннюю часть таллома вокруг основных осевых нитей и клеток внутренней коры.
У флоридеевых водорослей по особенностям анатомического строения различают два основных типа талломов - одноосевой, или центрально-нитчатый, и многоосевой, или фонтанный. В основе строения одноосевого таллома лежит единственная однорядная клеточная нить с боковыми веточками ограниченного роста (рис. 17.5). Примером дальнейшего усложнения строения таллома этого типа может служить таллом Lemanea Bory, в котором кольца боковых ветвей срастаются, образуя пустотелую трубку, с проходящей в середине осевой нитью (рис. 17.6). Слизистое вещество развивается незначительно и утрачивает значение скрепляющего материала. Центральная ось многоосевого таллома состоит из пучка параллельно идущих нитей. Их расположение напоминает струи воды в фонтане, поэтому этот тип организации называют фонтанным (рис. 17.7). Нити пучка могут размещаться плотно в центре таллома или кольцом, тогда в центре остается полость.
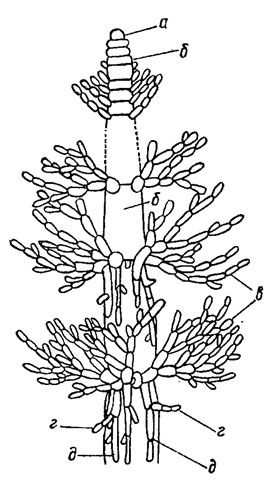
Рис. 17.5. Таллом красных водорослей с одноосевым типом строения (Sirodotia Kylin, Batrachospermum Roth): а - инициальная клетка; б - клетки центральной нити; в - кольца веточек ограниченного роста; г - образование побегов междоузлий; д - нити коры на междоузлиях
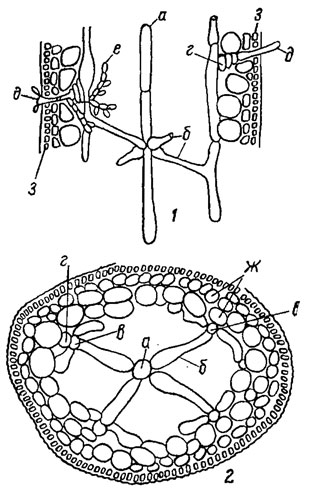
Рис. 17.6. Таллом Lemanea Bory: 1 - в продольном разрезе; 2 - в поперечном разрезе; а - клетки осевой нити; б - крестообразно расположенные клетки; в - боковые нити; г - карпогон; д - трихогина; с - гонимобласт; ж - подкоровые клетки; з - коровые клетки
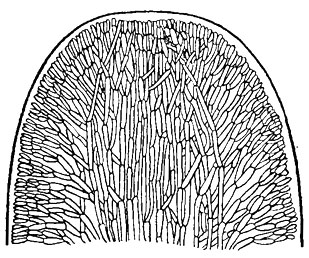
Рис. 17.7. Таллом красной водоросли многоосевого (фонтанного) типа строения в продольном разрезе (Furcellaria fastigiata (Huds.) Lamour.)
Псевдопаренхиматозное (ложнотканевое) строение тела свойственно преимущественно морским красным водорослям, у которых оно достигает значительного развития; у пресноводных багрянок умеренных широт этот тип строения можно обнаружить лишь у видов рода Thorea Bory (рис. 17.8).
Рис. 17.8. Красная водоросль с псевдопаренхиматозной структурой таллома: фрагмент таллома Thorea ramosissima Bory в продольном разрезе
Клетки красных водорослей разнообразны по форме - шаровидные, эллипсоидные, боченковидные, цилиндрические, дисковидные, многоугольные, с цельной оболочкой, в которой хорошо различаются два слоя: внутренний, состоящий из целлюлозы, и наружный - из пектиновых соединений. У примитивных багрянок класса Bangiophyceae (Chroothece, Cyanidium Geitl., Rhodospora Geitl., Goniotrichum, Chroodactylon, Phragmonema Zopf.) оболочки пектиновые, а целлюлоза отсутствует. У многих более сложно организованных багрянок внешняя часть оболочки может быть очень желатинизированной или преобразованной в пектиновую слизь.
Пектиновые вещества красных водорослей представляют собой кальциевые и магниевые соли особых пектиновых кислот. Они способны растворяться в кипящей воде с образованием коллоидных растворов. К группе пектиновых веществ относятся также коллоидные вещества, которые содержатся в клеточных оболочках и межклетниках многих морских багрянок (Phyllophora Grev.). Они представляют собой сложную смесь полисахаридов, содержащих серу, и носят общее название фикоколлоидов (агар, агароиды, каррагенин). Эти вещества отличаются друг от друга по составу и свойствам, но обладают общей желирующей способностью. В оболочках некоторых морских багрянок (Gelidium Lamour., Laurencia Lamour.) обнаружен амилоид - углевод такого же состава, что и гемицеллюлоза (запасная клетчатка).
Среди красных водорослей встречаются формы с обызвествленными оболочками. Состав инкрустирующих соединений извести неодинаковый. У водорослей семейства Corallinaceae откладывается кальцит, у некоторых Nemaliales - арагонит. Часто встречаются карбонаты кальция, магния, а также соли железа. Инкрустированным может быть весь таллом или только его часть. У некоторых групп морских багрянок инкрустация тела постоянна и является систематическим признаком (Lithothamnion Phil., Coralline L.). В пресных водах постоянная инкрустация талломов известью у багрянок отсутствует, она возникает при определенных экологических условиях и в определенном возрасте.
Оболочка клеток красных водорослей растет путем отложения новых слоев на вершине протопласта апикальной клетки. Наружные слои оболочки по мере роста клетки постепенно разрываются и раздвигаются. Так как новые слои откладываются всегда под некоторым углом, вся оболочка имеет слоистое строение (рис. 17.9). У форм с грубым жестким талломом снаружи клеточных стенок развивается тонкий слой кутикулы, отличающийся по составу и строению от кутикулы цветковых растений. На мягких слизистых талломах багрянок кутикула отсутствует.
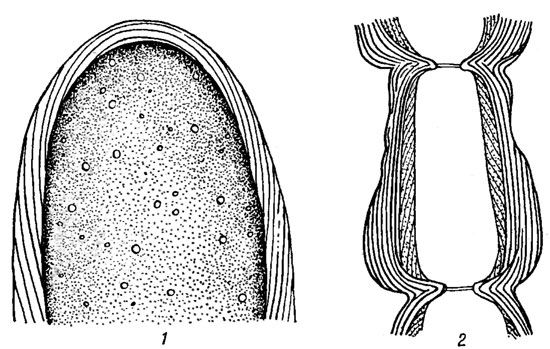
Рис. 17.9. Оболочки клеток красных водорослей: 1 - Bornetla secundiflora (J. Ag.) Thur.; 2 - Anthithamnion cruciatum (Ag.) Nag
Цитоплазма отличается большой вязкостью, плотно прилегает к клеточным стенкам, очень легко подвергается плазмолизу. Ядро одно (или ядер несколько), оно мелкое, имеет четкую оболочку и ядрышко. Одноядерные клетки свойственны преимущественно представителям класса Bangiophyceae. В классе Florideophyceae одноядерные клетки встречаются лишь у некоторых видов порядка Nemaliales и у примитивных форм порядков Cryptonemiales и Gigartinales. У морских высокоорганизованных багрянок клетки обычно многоядерные, за исключением молодых клеток таллома - апикальных и коровых. Среди них, однако, встречаются виды, у которых старые клетки центральных нитей одноядерные, а окружающие их более молодые клетки - обычно многоядерные (виды рода Callithamnion Lyngb., некоторые виды рода Polysiphonia Grev. и др.). Репродуктивные клетки - спермации, карпо- и тетраспоры - всегда с одним ядром, но яйцеклетки, как и ауксиллярные клетки, из которых развивается гонимобласт, часто окружены многоядерными клетками, даже у форм, весь таллом которых состоит из одноядерных клеток.
Хлоропласты багрянок разнообразны по форме и величине (рис. 17.10). У растений класса Bangiophyceae клетки содержат один хлоропласт, либо звездчатый с одним крупным пиреноидом в центре, либо пристенный, пластинчатый, без пиреноида. У видов класса Florideophyceae звездчатые хлоропласты встречаются лишь у примитивных представителей порядка Nemaliales. Пиреноид в хлоропластах этих водорослей находится в центре или сбоку. При боковом размещении пиреноида хлоропласт большей частью имеет вид лопастной пластины. У некоторых багрянок наблюдаются т. н. свободные пиреноиды, расположенные за пределами хлоропластов. Роль пиреноида у красных водорослей окончательно не выяснена. В одних случаях его присутствие связывают с синтезом и отложением зерен крахмала, однако пиреноиды встречаются и в клетках, не связанных с процессами ассимилляции. У высокоорганизованных флоридеевых водорослей хлоропласты не имеют пиреноида. Они располагаются в пристенном слое цитоплазмы и бывают двух основных типов - лентовидные и линзовидные (дисковидные) многочисленными переходами между ними. Лентовидные хлоропласты свойственны видам, находящимся на низкой ступени эволюции; для высокоорганизованных форм более характерны линзовидные хлоропласты; причем с усложнением организации водорослей увеличивается и число хлоропластов. Центральные клетки у высокоорганизованных форм лишены хлоропластов.
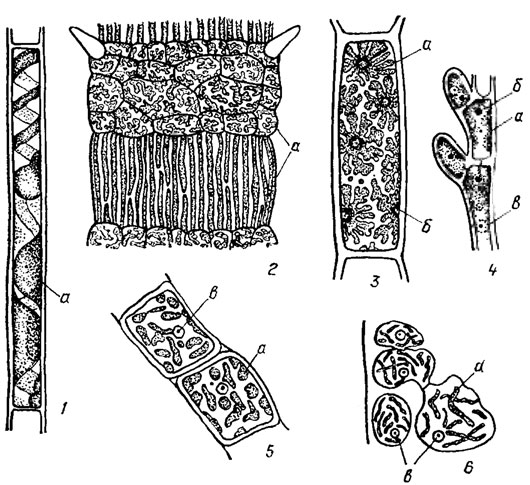
Рис. 17.10. Хлоропласты красных водорослей: 1 - Audoulnella sp.; 2 - Ceramlum sp.; 3 - Kylinia floridulum (Rosenv.) Papenf. 4 - Kylinia rhipidandra (Rosenv.) Papenf.; 5 - Asparagopsls armata Harv.; 6 - Cystoclonium purpureum (Huds.) Batt.; a - хлоропласты; б - пиреноиды; в - ядра
Форма хлоропластов багрянок не всегда постоянна; она может быть разной даже в разных клетках одного организма. Так, у видов рода Ceramium Roth в крупных клетках междоузлий хлоропласты вытянутые, лентовидные, а в мелких коровых клетках на узлах они пластинчатые, короткие, неправильно лопастные (см. рис. 17.10, 2). Форма хлоропластов может меняться также с возрастом растения, с изменением размеров клеток, при изменении условий освещения и аэрации. Размножение хлоропластов происходит путем простого деления (перешнуровыванием). По своему тонкому строению, видимому под электронным микроскопом, хлоропласты багрянок сходны с хлоропластами других водорослей.
Красные водоросли отличаются сложным набором пигментов. Наряду с хлорофиллами а и d, α- и β-каротинами и несколькими ксантофиллами, хлоропласты красных водорослей содержат специфические для данной группы растений дополнительные пигменты: красный фикоэритрин и синий фикоцианин. Эти пигменты относятся к группе фикобилипротеидов. обнаружены они кроме багрянок еще у синезеленых и криптофитовых водорослей. По химическому составу эти пигменты, однако, неидентичны в разных группах водорослей. Различное количественное соотношение фикоэритрина и фикоцианина определяет цвет красных водорослей. У морских водорослей все пигменты большей частью маскируются фикоэритрином, у пресноводных фикоэритрина мало либо он совершенно отсутствует, поэтому пресноводные багрянки окрашены обычно в серый, темно-зеленый, сине-зеленый или ярко-зеленый цвет. Фикоэритрин и фикоцианин растворяются в воде, после отмирания водорослей они вымываются из клеток.
Продуктом ассимиляции многих красных водорослей является полисахарид, называемый багрянковым крахмалом. От иода он не синеет, а сначала становится желто-бурым, затем красным. По химической природе он ближе всего к амилопектину и гликогену и, по-видимому, занимает промежуточное положение между обычным крахмалом и гликогеном. Откладывается багрянковый крахмал в виде мелких полутвердых телец, имеющих форму конусов или плоских овальных пластинок с концентрическими зонами либо с углублением на широкой поверхности (рис. 17.11). Зерна багрянкового крахмала образуются частично в цитоплазме, частично на поверхности хлоропластов, иногда скапливаются вокруг ядра или пиреноида, но они, в отличие от обычного крахмала зеленых растений, никогда не встречаются внутри пластид. Багрянковый крахмал образуется лишь у представителей класса Florideophyceae. У видов класса Bangiophyceae продуктом ассимиляции, по-видимому, является гликоген.
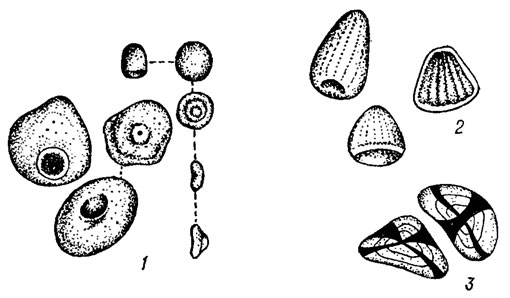
Рис. 17.11. Зерна багрянкового крахмала: 1 - Ptilota plumosa (L.) Ag.; 2 - Gracilaria dura (Ag.) J. Ag.; 3 - в поляризованном свете
В качестве запасных веществ у красных водорослей, кроме багрянкового крахмала, откладываются сахариды: галактозид, флоридозид и др. У некоторых форм встречаются многоатомные спирты. Состав жиров меняется в зависимости от условий среды.
Особенностью красных водорослей является наличие у некоторых представителей класса Florideophyceae особых клеток с бесцветным содержимым, сильно преломляющих свет (рис. 17.12). Эти клетки известны как пузырчатые, друзовые, или железистые. Их содержимое у разных водорослей отличается по химическому составу. Они встречаются, как правило, в поверхностном слое таллома и заполнены йодистыми, реже бромистыми соединениями. Железистые клетки встречаются чаще всего у представителей порядка Ceramiales. Роль пузырчатых клеток не выяснена. Возможно, они являются недоразвитыми спорангиями, хранилищем запасных веществ, "плавательными пузырями" и т. д. Железистые клетки характерны для определенных групп красных водорослей и поэтому имеют важное таксономическое значение.
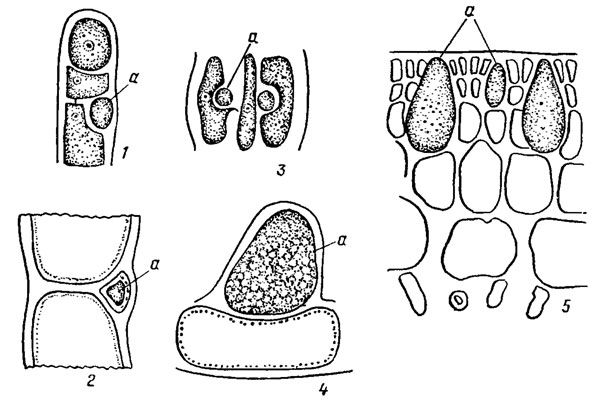
Рис. 17.12. Железистые клетки (а) красных водорослей: 1, 2 - Trailliella intricata Batt.; 3 - Falkenbergia hillebrandii (Born.) Falkenb.; 4 - Antithamnion plumula (Ell.) Thur.; 5 - Opuntiella californica (Farl.) Kyl
Для защиты от сильного света у багрянок, живущих на небольших глубинах, особенно в тропических и субтропических морях, служат особые иридирующие тельца, мутно-желтые, неправильной формы (рис. 17.13). Они образуются в вакуолях поверхностных клеток таллома, состоят из мелких зернышек протеиновой природы и обладают способностью рассеивать и отражать падающий на них свет. При очень сильном освещении иридирующие тельца располагаются под внешней стенкой клетки и служат своеобразным занавесом для хлоропластов. При рассеянном свете происходит взаимное перемещение этих органелл, и хлоропласт оказывается у верхней стенки.
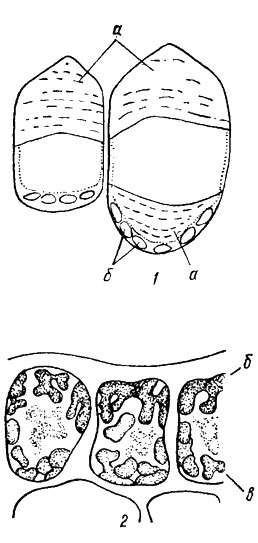
Рис. 17.13. Иридирующие тельца красных водорослей: 1 - Gasteroclonium clavatum (Roth) Ardiss.; 2 - Nitophyllum sp.; а - крупные иридирующие тельца; б - хлоропласты; в - мелкие иридирующие тельца
Для многих представителей класса Florideophyceae характерно образование волосков. При этом необходимо отличать настоящие волоски багрянок от волосовидных ветвей и ложных волосков. Ложные волоски обычны для видов с однорядным нитчатым талломом. При их образовании конечные клетки веточек удлиняются, обесцвечиваются и приобретают волосовидное строение. Настоящие волоски красных водорослей бывают двух типов одноклеточные и многоклеточные. Одноклеточные волоски не ветвятся. У однорядных нитчатых форм они образуются из апикальных клеток ветвей, у многорядных - из поверхностных клеток коры. Клетка будущего волоска отделяется от материнской клетки поперечной перегородкой, сильно вытягивается (до 1 мм и больше). Материнская клетка волоска обычно ничем не отличается от соседних вегетативных клеток, но иногда она намного крупнее и остается хорошо заметной после отпадения волоска. Клетка волоска содержит ядро и небольшое количество цитоплазмы. Хлоропласт по мере роста волоска исчезает и волосок становится бесцветным (рис. 17.14). У флоридеевых волоски встречаются довольно часто, у бангиевых они отсутствуют. Многоклеточные волоски обычно более или менее сильно разветвлены, встречаются они только у некоторых видов порядка Ceramiales (Florideophyceae). Роль волосков окончательно не выяснена. Некоторые исследователи считают, что они способствуют поглощению питательных веществ из окружающей среды, другие приписывают им защитную роль при чрезмерном освещении.
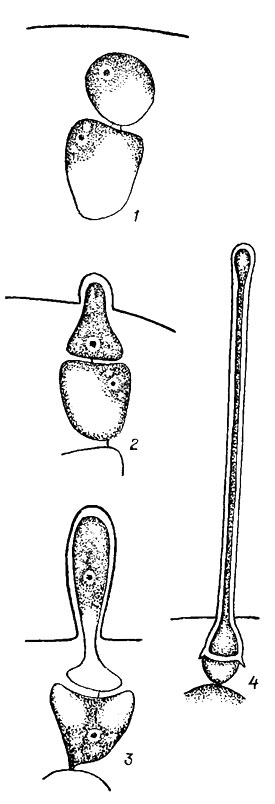
Рис. 17.14. Стадии развития (1-4) волоска у Cystoclonium purpureum (Huds.) Batt
Клетки, слагающие таллом, соединяются между собой с помощью пор, среди которых различают первичные и вторичные (рис. 17.15). Соединение между дочерними клетками, происходящими от одной материнской, осуществляется посредством первичных пор. Они формируются в результате развития неполной перегородки между двумя вновь образующимися клетками. Первичные поры расположены в середине перегородки и представляют собой тонкую пластину, через которую проходят тяжи, соединяющие цитоплазму соседних клеток. Первичные поры свойственны не только флоридеевым, как считалось ранее, но и некоторым бангиевым. Кроме первичных пор, у многих представителей класса Florideophyceae происходит и вторичное соединение соседних клеток. Образование вторичных пор начинается с деления ядра и перемещения одного из дочерних ядер к периферии клетки. В том месте, куда попадает ядро, образуется небольшой вырост, который затем отчленяется, как маленькая клетка, соединенная с первой посредством первичной поры. Вытягиваясь, эта дополнительная клетка достигает соседней и сливается с ней. Теперь первичная пора соединяет уже две соседние клетки и становится вторичной. Иногда две соседние клетки соединены несколькими вторичными порами. Роль этих соединений до конца не выяснена; возможно, они способствуют переносу питательных веществ от одной клетки к другой.
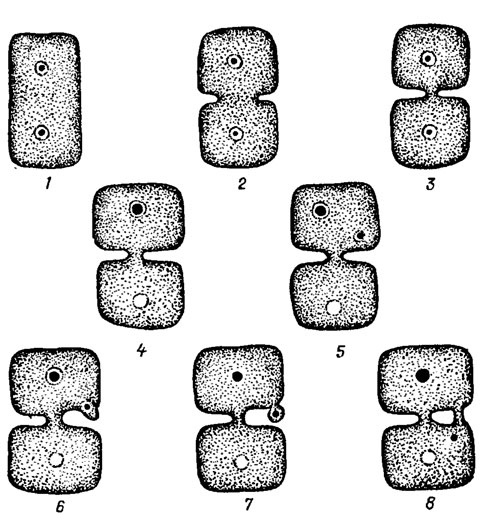
Рис. 17.15. Схема образования первичных (1-3) и вторичных (4-8) пор в клетках талломов красных водорослей
Размножаются красные водоросли бесполым и половым путем. Наиболее простой способ вегетативного размножения - деление клетки на две или более дочерних. Таким образом размножаются одноклеточные и колониальные представители класса Bangiophyceae. Вегетативное размножение водорослей класса Florideophyceae происходит посредством образования дополнительных побегов, берущих начало от подошвы или стелющихся ветвей. У ряда флоридеевых таллом способен к регенерации и пролиферации. Так, у видов родов Lemanea и Batrachospermum Roth часто прорастают коровые клетки, образуя молодую "заростковую" стадию. У Hildenbrandtia rivularis (Liebm.) Ag. образуются специальные вегетативные бородавочки (гемы). У многих флоридеевых в определенный период отмирает вертикальная часть таллома и остается только базальная (горизонтальная) часть растения, которая через некоторое время прорастает, образуя новые побеги.
Собственно бесполое размножение красных водорослей происходит с помощью разнообразных спор. У низкоорганизованных багрянок класса Bangiophyceae - это моноспоры, образующиеся по одной в клетке, называемой моноспорангием, в результате превращения всего протопласта (рис. 17.16). У бангиевых моноспоры образуются в любой клетке "таллома и до выхода из нее не отличаются от вегетативных клеток. В некоторых случаях в моноспору может превратиться лишь часть клетки, предварительно отделившаяся от нее (Erythrotrichia), или вегетативная клетка при образовании моноспоры делится на 2-4 споры (Bangia). Моноспоры характерны для всех бангиевых, тогда как среди флоридеевых они известны, за очень немногим исключением, только в порядке Nemaliales. При этом у водорослей, построенных по типу свободной нити, моноспорангии образуются на веточках ограниченного роста. В них превращаются конечные клетки веточек. Иногда моноспорангии встречаются группами по 2-3 на одной и той же клетке (см. рис. 17.16, 1-3). У водорослей, имеющих более плотное строение и дифференцированный коровой слой (Thorea), в моноспорангии превращаются внешние клетки ассимиляционных нитей (см. рис. 17.16, 4). Созревшие моноспорангии открываются на вершине, голые моноспоры выскальзывают из них, а через некоторое время одеваются оболочкой и начинают прорастать. Часто наблюдается повторное образование моноспор в стенках старого моноспорангия (см. рис. 17.16, 2).
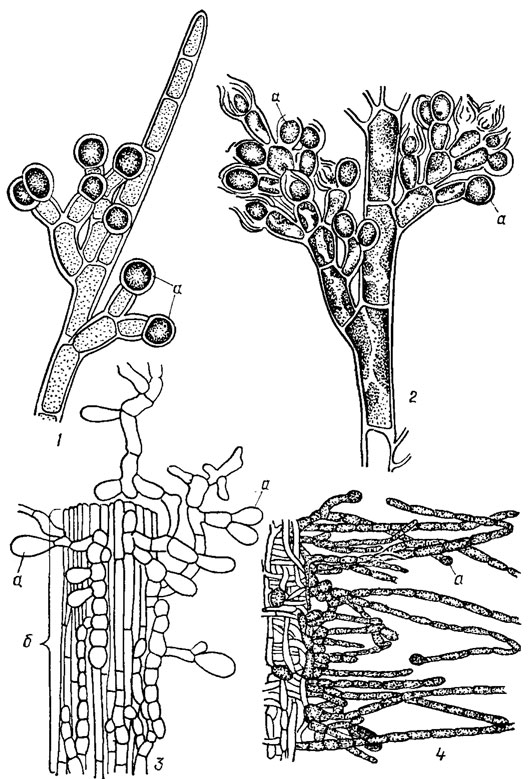
Рис. 17.16. Моноспорангии (а) флоридеевых водорослей: 1 - Chantransla pygmaea Kütz.; 2 - Chantransia sinensis Jao; 3 - Balbianla Investiens (Lenorm.) Slrod. на междоузлии (б) Batrachospermum; 4 - Thorea ramoslssima Bory
Для подавляющего большинства флоридеевых водорослей наиболее обычны тетраспоры, образующиеся в тетраспорангии (рис. 17.17). По способу деления содержимого клетки различают три типа тетраспорангиев: зонально, крестообразно и тетраэдрически разделенные. При зональном делении все три перегородки ориентированы параллельно друг другу, в результате чего тетраспоры располагаются в ряд (см. рис. 17.17, 2). В крестообразно разделенных тетраспорангиях клетка сначала делится поперечной перегородкой пополам, после чего каждая из двух образовавшихся клеток делится продольной перегородкой, при этом обе перегородки проходят в одной плоскости или под прямым углом друг к другу (см. рис. 17.17, 4, 5). В тетраэдрически разделенных тетраспорангиях клеточные перегородки ориентированы косо (см. рис. 17.17, 1).
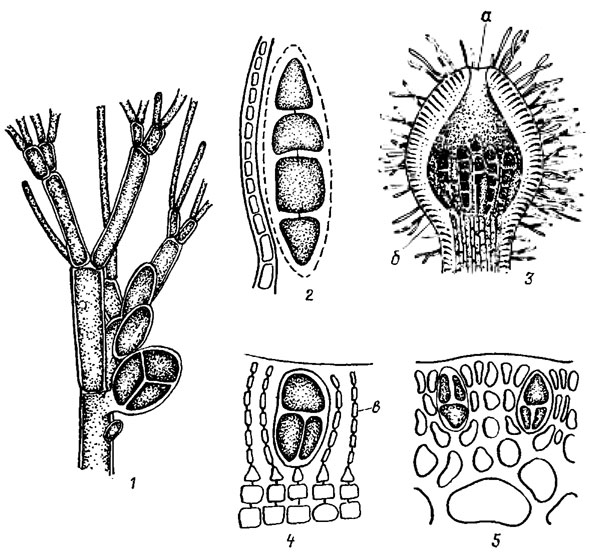
Рис. 17.17. Тетраспорангии флоридеевых водорослей: 1 - Callithamnion corymbosum (J. Sm.) Lyngb., 2 - Cruoria pellita (Lyngb.) Fries; 3 - Corallina mediterranea Aresch.; 4 - Peyssonnelia dubia Des., 5 - Gracilaria sp.; a - выводное отверстие; б - кондептакул; в - нити нематеция
Способ деления тетраспорангиев - постоянный признак, используемый в систематике. Постоянным таксономическим признаком является также место образования тетраспорангиев. Как правило, тетраспорангии занимают положение конечных клеток веточек ограниченного роста. Интеркалярное возникновение их наблюдается крайне редко. Однако в процессе роста тетраспорангиев, они оказываются погруженными в коровой слой, и их положение конечной клетки в зрелом состоянии не всегда отчетливо видно (см. рис. 17.17, 4). Располагаются тетраспорангии рассеянно или группами, часто в виде цепочки. У корковой водоросли Cruoria pellita (Lyngb.) Fries тетраспорангии представляют собой боковые выросты вертикальных нитей (см. рис. 17.17, 2). У некоторых видов тетраспорангии развиваются в нематециях - специальных образованиях на поверхности талломов, имеющих вид небольших подушечек или горбиков. Тетраспорангии развиваются среди нитей нематеция, тесно окруженные ими. Отчленяются они от поверхности клеток корового слоя (см. рис. 17.17, 4) или от самых нижних клеток нитей нематеция. У представителей семейств Hildenbrandtiaceae и Corallinaceae тетраспорангии образуются в специальных вместилищах - концептакулах (см. рис. 17.17, 3).
Половое размножение свойственно всем красным водорослям класса Florideophyceae; в классе Bangiophyceae оно встречается только у наиболее высокоорганизованных форм. Половой процесс оогамный. Мужские и женские гаметы лишены жгутиков. Однако с помощью электронного микроскопа на поперечном срезе сперматангиев у Bonnemaisonia hamifera Hariot (Florideophyceae) удалось обнаружить в спермациях структуры, предшествующие образованию двух жгутиков (т. н. скрытые жгутики) - крипты [634]. Это наблюдение чрезвычайно важно, поскольку оно, после тщательной его проверки, позволит в дальнейшем по-новому подойти к проблеме филогенетических связей Rhodophyta.
При оплодотворении женские гаметы - оогонии - остаются в гаметангии на гаметофите, мужские гаметы - спермации - выбрасываются наружу и пассивно переносятся током воды. Спермации - шаровидные или яйцевидные клетки, У бангиевых спермации голые, у флоридеевых отсутствие оболочки у спермациев окончательно не установлено. Известно только, что в момент прикрепления к женскому растению они уже окружены тонкой оболочкой. Спермации образуются в клетках, которые функционируют как мужские органы размножения и называются сперматангиями, или антеридиями (рис. 17.18, 1-7). В каждом сперматангии образуется по одному спермацию. Вегетативную клетку, дающую начало сперматангиям, называют материнской. У бангиевых водорослей любая клетка таллома, за исключением ризоидных, может стать материнской клеткой сперматангиев. Однако сперматангии возникают преимущественно в верхних частях таллома. У нитчатых флоридеевых в сперматангии преобразуются конечные клетки веточек ограниченного роста, у багрянок с псевдопаренхиматозным строением в сперматангии превращаются конечные клетки веточек коры.
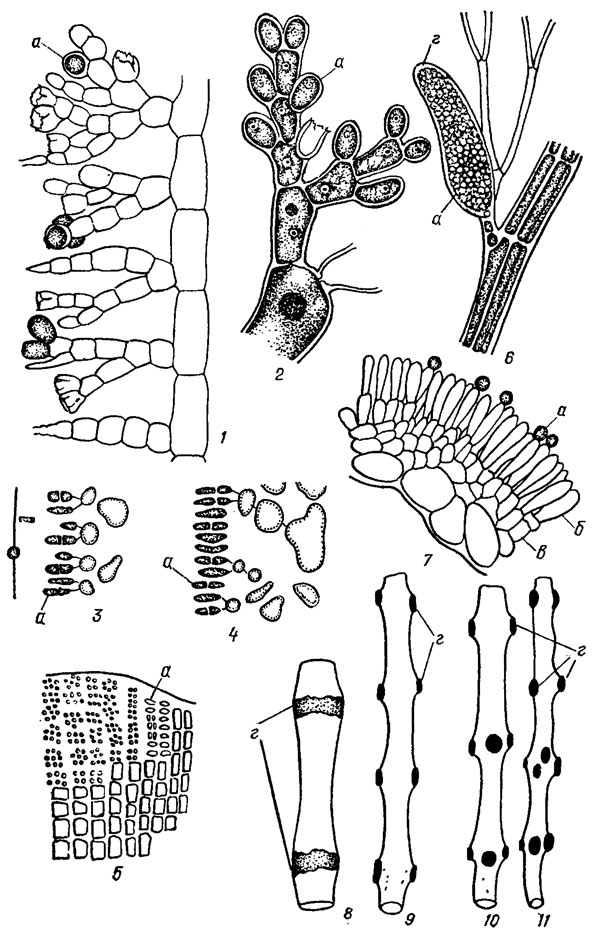
Рис. 17.18. Сперматангии (а) флоридеевых водорослей: 1 - участок таллома Batrachospermum orthostichum; 2 - участок таллома Nemalion multifidum (Web. et Mohr) Ag.; 3, 4 - сперматангии Gelidium sp. в поперечном разрезе; 5 - нематеций Peyssonnelia sp.; 6 - колосовидный сорус Poiysiahonia sp.; 7 - сорус Lemanea catenate Kütz. в поперечном разрезе; 8 - сорусы на талломе Lemanea grand is (Wolie) Atkinson; 9-11 - сорусы на талломах Lemanea mamlllosa (Sirod.) De Toni. a - сперматангии; б - антеридиальные нити; в - базальные клетки, г - сорус
Каждая материнская клетка отчленяет один или несколько сперматангиев. После созревания первых сперматангиев и выпадения спермациев, материнская клетка может образовать и отчленить еще несколько вторичных сперматангиев. У большинства багрянок сперматангии собраны в группы - сорусы и только у некоторых располагаются рассеянно. У нитчатых форм сорусы имеют вид густых пучков, расположенных у вершины ветвей, у водорослей с плотным талломом (Lemanea) сорусы располагаются на их поверхности в виде подушковидных образований (см. рис. 17.18, 8-11), иногда они оказываются погруженными в небольшие углубления на талломе, а у водорослей семейства Corallinaceae размещаются в концептакулах. У высокоорганизованных Ceramiales сперматангии образуются на специальных коротких веточках - трихобластах.
Оогоний красных водорослей называют карпогоном. Это особая клетка, заполненная цитоплазмой и лишенная хлоропласта. Только у примитивных представителей флоридеевых и у бангиевых карпогон содержит окрашенный хлоропласт. У бангиевых карпогон представляет собой слабо измененную вегетативную клетку, отличающуюся от других несколько большими размерами и более интенсивной окраской. У флоридеевых карпогон имеет своеобразную форму (рис. 17.19). Его нижняя, или брюшная, часть, конусовидная и разнообразно вздутая, содержит ядро, верхняя вытянутая узкая часть образует трубчатый вырост разной длины - трихогину, служащую для улавливания спермациев. Благодаря трихогине карпогон легко отличить от остальных клеток таллома. Трихогина может быть короткой, кеглевидной, бутылковидной, обратнояйцевидной или длинной, узкой, цилиндрической, прямой или спирально скрученной (рис. 17.20). У бангиевых водорослей трихогина отсутствует, лишь иногда карпогоны имеют короткие сосочковидные выросты. Такие карпогоны трудно отличить от обычных вегетативных клеток. У большинства флоридеевых карпогон развивается из конечной клетки короткой 3-4-клеточной карпогонной нити или ветви, иногда погруженной в окружающую ткань. Клетка, от которой отходит карпогонная ветвь, называется несущей, или базальной; в состав карпогонной ветви она не входит. Почти у всех флоридеевых карпогонная ветвь образуется как специальная боковая веточка во внутренней части корового слоя. Только у низкоорганизованных водорослей порядка Nemaliales она соответствует обычной боковой веточке таллома. У багрянок со свободнонитчатым талломом карпогон занимает открытое положение (Balbiania Sirod., Sirodotia Kylin), у багрянок с плотным талломом (Lemanea) карпогон оказывается погруженным и только трихогина достигает поверхности таллома и выступает над ней.
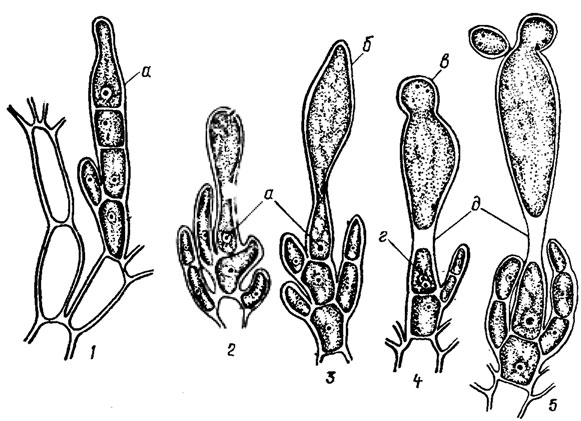
Рис. 17.19. Стадии развития (1-5) карпогона Batrachospermum Roth: а - ядро карпогона; б - трихогина; в - спермий при слиянии с трихогиной; г - слияние спермия и карпогона; д - образование перегородки между трихогиной и карпогоном
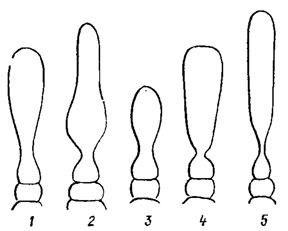
Рис. 17.20. Формы трихогины: 1 - булавовидная; 2 - бутылковидная (урновидная); 3 - яйцевидная; 4 - обратноконусовидная; 5 - цилиндрическая с ножкой
При оплодотворении спермации прикрепляются к вершине трихогины, оболочки их в месте соприкосновения растворяются и ядро спермация перетекает в полость трихогины. Передвигаясь по ней, оно попадает в брюшную часть карпогона и там сливается с его ядром. После оплодотворения карпогон отделяется от трихогины поперечной перегородкой, трихогина отмирает. Оплодотворенная яйцеклетка, или зигота, остается заключенной в оболочку карпогона, собственной же оболочки она не образует. Зигота прорастает без периода покоя, претерпевая ряд сложных преобразований, в результате которых образуются особые споры - карпоспоры, дающие начало спорофиту. Клетки, в которых образуются карпоспоры, называют карпоспорангиями. Карпоспоры представляют собой голые, лишенные жгутиков клетки, неподвижные или изредка обнаруживающие амебоидное движение. Перед началом прорастания они одеваются оболочкой. В каждом карпоспорангии формируется по одной карпоспоре.
У разных групп багрянок превращение зигот в карпоспоры происходит по-разному. У бангиевых этот процесс весьма прост: после оплодотворения карпогон делится на 4-32 карпоспоры, которые в виде голых клеток выходят в воду. У флоридеевых развитие зиготы - сложный многообразный процесс, в котором, кроме карпогона, участвуют многие другие специальные клетки. После оплодотворения карпогона развиваются особые спорогенные нити из небольшого числа клеток (нити гонимобласта). В наиболее простом случае (порядок Nemaliales род Batrachospermum) оплодотворенный карпогон отчленяет от себя несколько боковых клеток, которые, делясь, дают пучок нитей гонимобласта, конечные клетки которых превращаются в карпоспорангии (рис. 17.21). В этом случае нити гонимобласта развиваются непосредственно из карпогона. Гонимобласты обычно шаровидные, реже эллипсоидные или овалоидные. Не совсем правильно гонимобласты этой группы растений называют иногда цистокарпиями, или цистокарпами.
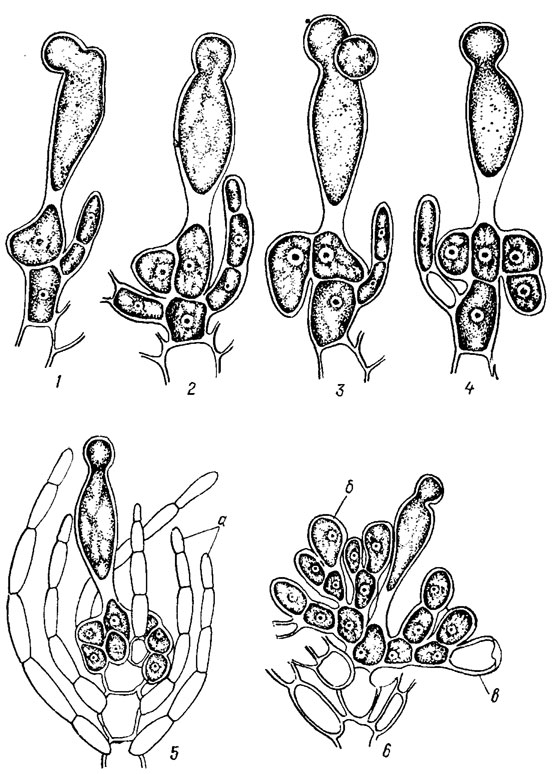
Рис. 17.21. Стадии развития гонимобласта Batrachospermum Roth.: 1-4 - первые клетки гонимобласта, образовавшиеся из оплодотворенного карпогона; 5 - молодой карпогон; 6 - карпогон с карпоспорами; а - стерильные веточки карпогона, или брактеа; б - карпоспора; в - пустой карпоспорангий
У подавляющего большинства флоридеевых водорослей образованию нитей гонимобласта предшествует слияние карпогона с особыми клетками, богатыми питательными веществами, среди которых различают клетки двух типов: просто питающие клетки, которые образуются до и после оплодотворения и служат только для питания развивающегося гонимобласта, и настоящие ауксиллярные клетки. Последние также функционируют как питающие клетки, но их главная роль состоит в том, что они представляют собой исходную клетку, от которой начинается развитие гонимобласта. Этому процессу обязательно предшествует слияние карпогона с ауксиллярной клеткой. Слияние осуществляется с помощью специальных нитей или выростов, называемых соединительными, или ообластемными, которые отходят от карпогона и тянутся, пока не соединятся с ауксиллярной клеткой. По этим нитям ядро оплодотворенного карпогона передвигается и переходит в ауксиллярную клетку, но слияния ядер не происходит, поскольку ауксиллярная клетка дает лишь дополнительное питание при образовании гонимобласта. У некоторых багрянок соединительные нити продолжают расти дальше и последовательно сливаются с другими ауксиллярными клетками, образуя гонимобласты в месте каждого слияния. Таким образом, у флоридеевых водорослей, имеющих ауксиллярные клетки, гонимобласты развиваются не от карпогона, а от ауксиллярной клетки. Наиболее примитивными считаются те багрянки, которые имеют несколько ауксиллярных клеток, удаленных от карпогона.
Цистокарпом называют зрелый гонимобласт, заключенный в обвертку (оболочку), созданную клетками коровых нитей и клетками, расположенными рядом с карпогонными ветвями. Для выхода карпоспор в обвертке цистокарпа образуются поры. Однако они возникают не всегда, и тогда карпоспоры освобождаются после разрушения оболочки цистокарпа.
Циклы развития красных водорослей описаны в разделе 3.2.4.
Современная классификация водорослей основывается на исследованиях крупного шведского альголога Г. Кюлина. В целом она принимается всеми альгологами, противоречия возникают только в отношении систематического положения некоторых мелких таксонов. В основу этой классификации положено строение женских репродуктивных органов и процесс развития гонимобласта. По этой системе все красные водоросли делятся на два класса - Bangiophyceae и Florideophyceae. Основная масса багрянок относится ко второму классу.
Вопрос о происхождении красных водорослей и их филогенетических связях с другими отделами растительного мира проблематичен и до сего времени трактуется разными авторами по-разному. Одни исследователи связывают их возникновение с зелеными водорослями. Предположение о возможных родственных связях красных и зеленых водорослей основывается, с одной стороны, на сходстве строения таллома Prasiola Ag. из улотриксовых водорослей и Porphyra из бангиевых, а с другой - на сходстве оогония видов рода Coleochaete Breb. и его прорастающей зиготы с карпогоном и цистокарпом флоридеевых водорослей.
Некоторые исследователи [518, 635 и др.] на основании сходства пигментов красных и синезеленых водорослей, близости багрянкового крахмала к крахмалу синезеленых и отсутствия подвижных стадий выводят примитивные багрянки (класс Bangiophyceae), у которых еще отсутствует половой процесс, от синезеленых водорослей, а класс Florideophyceae от эволюционно продвинутых бангиевых водорослей, которым уже свойствен половой процесс. По мнению других исследователей, оба отдела водорослей возникли в одной и той же геологической эпохе в сходных условиях освещения, имеют самостоятельное происхождение и непосредственные родственные связи между ними отсутствуют [82, 120, 433 и др.].
Что касается филогенетических связей в пределах отдела красных водорослей, то по мнению У. Прескотта [596], флоридеевые произошли от бангиевых водорослей. У. Прескотт указывает два возможных пути их происхождения - от примитивных бангиевых и от форм более продвинутых, близких к роду Compsopogon Mont.
Красные водоросли широко распространены во всех морях земного шара, однако по направлению к высоким широтам общее число видов багрянок постепенно уменьшается и резко падает в Арктическом бассейне.
Поселяются морские красные водоросли обычно на каменистом субстрате - скалах, рифах, валунах, на разнообразных искусственных сооружениях, а также на живых и отмерших организмах как животного, так и растительного происхождения. Благодаря наличию красных пигментов, способных усваивать даже незначительные количества света в спектральном диапазоне, недоступном для других водорослей, багрянки могут расти на глубине 100-200 м. Обычной границей их массового распространения является глубина 40-60 м, в зависимости от грунтов, благоприятных для их поселения. Чаще багрянки развиваются в верхних горизонтах моря, в том числе и на литорали, где периодически происходит смена среды обитания, обусловленная приливно-отливными явлениями, в связи с чем резко меняются такие экологические факторы как температура, соленость, инсоляция и т. п. На открытых скалистых участках в зоне брызг и заплесков чаще всего встречаются представители родов Bangia и Porphyra.
Среди морских красных водорослей (преимущественно Florideophyceae) имеется большое количество эпифитов и эндофитов. Эндофитизм бывает частичным или полным. В последнем случае таллом эндофита, питающегося автотрофно за счет фотосинтеза, полностью погружен в ткани водоросли-хозяина, на поверхность выносятся только нити с органами размножения. Немало багрянок ведут паразитический и полупаразитический образ жизни. Талломы эндофитных и паразитических водорослей значительно редуцированы и выступают над поверхностью водоросли-хозяина в виде шариков или лопастных образований.
В пресных и солоноватых континентальных водоемах обитают немногочисленные представители примитивных наименее специализированных багрянок, существенно отличающихся по экологии от морских. Одни из них принадлежат к холодноводным реофильным формам, растут в реках, ручьях с низкой температурой и быстрым течением, другие обитают в более теплых стоячих или слабо проточных водах. Поселяются пресноводные багрянки на раковинах живых брюхоногих и двустворчатых моллюсков, на обработанной и необработанной древесине, каменистом субстрате, а также на крупноклеточных нитчатых водорослях и водных цветковых растениях. Они могут существовать как при значительном освещении, так и в затененных условиях (например, Hildenbrandtia rivularis (Liebm.) Ag. - типичный тенелюбивый вид).
Аэрофитные формы багрянок обитают на влажной затененной почве, камнях, старых стенах оранжерей. Массовое развитие одноклеточной водоросли Porphyridium purpureum (Bory) Drew et Ross. связно обычно с присутствием в почве органических соединений азота.
Морские красные водоросли играют важную роль в природе, являясь важным источником органических веществ, а также пищей для многих морских животных, они несомненно играют определенную роль в процессах естественного самоочищения воды, усиливая окисление и конечную минерализацию органических веществ. Известковые, преимущественно водоросли семейства Corallinaceae, принимают участие в образовании коралловых рифов; благодаря цементирующим свойствам они способствуют сохранению рифовой структуры.
Хозяйственное значение морских багрянок определяется присутствием в оболочках некоторых водорослей большого количества фикоколлоидов - слизистых веществ из группы полисахаридов. Наиболее известным промышленным продуктом является агар, получаемый из видов родов Gelidium Lamour., Gracilaria Grev., Ahnfeltia E. Fries и других агарофитов. Агар широко применяют в медицине, микробиологии, пищевой, целлюлозно-бумажной и текстильной промышленности. Из других фикоколлоидов существенное значение имеют каррагенин и агароид. Каррогенин получают в основном из видов рода Chondrus Stack., агароид - из черноморской Phyllophora nervosa (D. С.) Grev. Морские красные водоросли вместе с другими водорослями используют также для изготовления водорослевой муки, идущей на корм домашним животным и на удобрение. Ряд морских багрянок человек употребляет в пищу в виде салатов, приправ, гарниров для мяса и рыбы. Первое место среди съедобных багрянок занимают виды родов Rhodymenia Grev. и Porphyra. В Японии развито промышленное культивирование порфиры в естественных условиях. Аквакультура некоторых красных водорослей широко практикуется в США, Канаде, Франции, Норвегии, Дании, Великобритании и других странах. В нашей стране морские водоросли выращивают на Дальнем Востоке, на Черном, Белом и Баренцевом морях [126, 155, 180, 293].
Некоторые пресноводные багрянки, довольно чутко реагирующие на гидрохимические изменения среды, используют как биологические индикаторы [311]. Показателями ксеносапробных условий в водоемах являются, при высокой продуктивности, Batrachospermun moniliforme Roth, В. vagum (Roth) Ag., Hildenbrandtia rivularis (Liebm.) Ag. Четко выраженным ксеносапробом следует считать Lemanea torulosa (Roth) Ag. Для олигосапробных водоемов характерны Lemanea fluviatilis С. Ag., L. nodosa Kütz., L. annulata Kütz., Chantransia chalybea (Roth) Fr. и др.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'