16. Отдел 7. Желтозеленые водоросли - Xanthophyta (Confervales, Heterocontae, Xanthophyceae, Xanthophycophyta, Tribophyceae, Botrydiophyta) (Н. П. Масюк)
(В системах многих авторов - класс Xanthophyceae [413, 415, 417, 423, 578, 581 и др.].)
Отдел объединяет около 600 видов преимущественно микро-, реже макроскопических эукариотических водорослей желто-зеленого, светло- или темно-желто-зеленого, реже зеленого цвета, иногда бесцветных. Размеры их колеблются от 0,5-1,5 мкм (Chloridella glaciales Kol) до нескольких миллиметров в диаметре (Botrydium Wallr.) или до десятков сантиметров в длину (нити Tribonema Derb. et Sol., Vaucheria D. С.). Индивиды одноклеточные, многоклеточные и неклеточные, либо колонии индивидов (в том числе ценобиальные), активно подвижные и неподвижные, прикрепленные и свободноживущие.
Наблюдаются почти все типы структуры вегетативного тела от монадного до сифонального, исключая сифонокладальный (рис. 16.1). Преобладающее большинство желтозеленых водорослей имеет коккоидный тип структуры (Mischococcales), реже наблюдаются монадный (Chloramoebales) и гемимонадный (Heterogloeales), ризоподиалъный (Rhizochloridales), сарциноидный (Chlorellidium Visch. et Pasch., Chlorellidiopsis Pasch.), нитчатый (Neonemataceae, Tribonemataceae), разнонитчатый (Heterococcus Chod.), псевдопаренхиматозный (Chaetopedia Pasch.), паренхиматозный (Chloropedia Pasch.) и сифональный (Botrydiales).
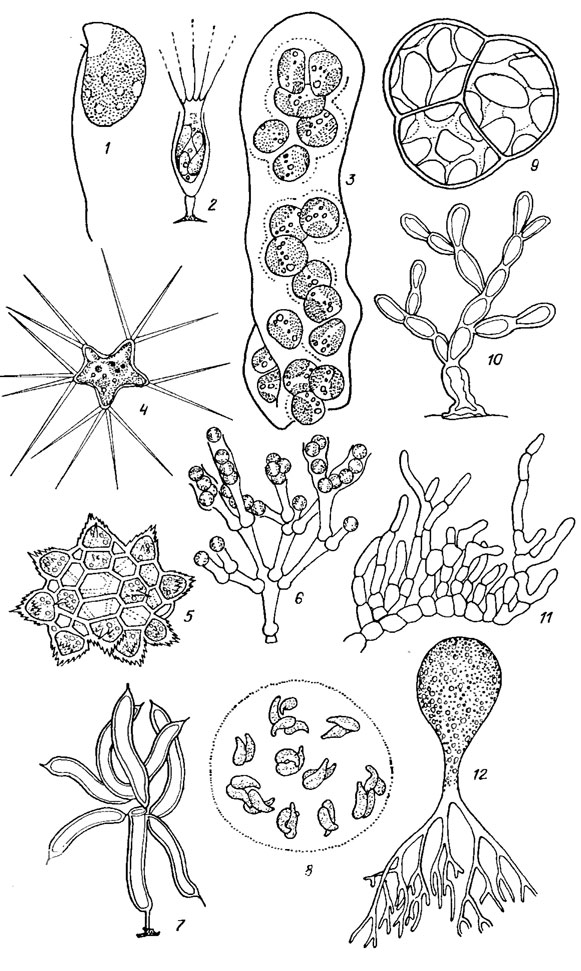
Рис. 16.1. Типы структуры вегетативного тела желтозеленых водорослей: 1 - монадный (Pedinomonadopsis minor Massjuk et Guk); 2 - ризоподиальный (Stipitococcus calix Ettl); 3 - гемимонадный (Helminthogloea ramosa Pasch., апикальная часть слизистого шнура); 4-8 - коккоидный (4 - Pseudcpolyedriopsis skujae Gollerb., 5 - Ducelliera chodatii (Ducel.) Teil., 6 - Mischococcus confervicola Näg., 7 - Ophiocytium mucronatum (A. Br.) Rabenh., 8 - Diplochloris decussata Korsch.); 9 - сарциноидный (Chlorellidium tetrabotrys Visch. et Pasch.); 10 - нитчатый (Heterodendron squarrosurn Pasch.); 11 - разнонитчатый (Heteroccoccus sp.); 12 - сифональной (Botrydium granulatum (L.) Grev.)
В цитологическом отношении желтозеленые водоросли изучены недостаточно. Отрывочные ультраструктурные данные свидетельствуют об отсутствии у них единого типа клеточной организации. Разнообразны клеточные покровы желтозеленых водорослей. У видов, обладающих амебоидной, монадной и отчасти гемимонадной структурой вегетативного тела, клетки покрыты лишь плазмалеммой; они легко изменяют свою форму, образуя лобоподии или ризоподии (Rhizochloris Pasch.). У Chlorocardion Pasch., Phacomonas Lohm. клетки мало метаболичны, что позволяет предполагать наличие у них более сложных клеточных покровов типа перипласта [207]. Это предположение нуждается в электронно-микроскопическом подтверждении. Иногда "голые" клетки находят в домиках (Stipitococcus W. et G. S. West), стенки которых могут быть окрашены в бурый цвет солями железа или марганца. У преобладающего большинства желтозеленых водорослей клетки имеют плотную клеточную оболочку, цельную или двустворчатую; в последнем случае створки могут быть равными или неравными (рис. 16.2). Иногда верхняя часть оболочки прикрывает нижнюю, подобно крышечке (см. рис. 16.2, 3). Нередко протопласт формирует наперстковидные вставочные образования, благодаря которым оболочка удлиняется по мере роста клетки. Это создает косую слоистость оболочки, заметную на одной или на обеих створках (см. рис. 16.2, 1). У нитчатых водорослей с двустворчатыми оболочками дезинтеграция нитей ведет к распаданию клеточных оболочек на Н-образные участки, представляющие собой тесно соединенные половинки оболочек двух соседних клеток. Двустворчатое строение оболочек иногда трудно различимо и может быть выявлено лишь с помощью 60%-го раствора гидроксида калия или специального окрашивания. Клеточная оболочка пектиновая, иногда с примесью целлюлозы или гемицеллюлозы (Tripanochloris clausilie Geitl., Tribonema), у Vaucheria - целлюлозная. Тонкая и нежная у молодых клеток с возрастом клеточная оболочка утолщается, становится многослойной. На ее поверхности может отлагаться известь, сначала в виде отдельных гранул, которые затем сливаются, образуя сплошную корку. Нередко оболочка насыщена соединениями железа, окрашивающими ее в желтоватый, красноватый, ржаво-красный, бурый или коричневый тона. Иногда она покрыта коричневыми пятнами (Chloridella ferruginea Pasch.) или бородавками (Centritractus brunneus Fott). Еще чаще в клеточных оболочках обнаруживается кремнезем, придающий ей особые твердость, хрупкость и блеск.
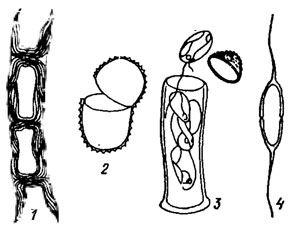
Рис. 16.2. Двустворчатые клеточные оболочки желтозеленых водорослей: 1 - Tribonema sp.; 2 - Chlorallantus oblongus Pasch.; 3 - Chlorothecium gladius Pasch.; 4 - Centritractus belonophorus Lemm
Плотная клеточная оболочка обеспечивает определенную форму клеток, нередко видоспецифичную: от шаровидной и полу шаровидной до шаровидно-ограненной, эллипсоидной, яйцевидной, грушевидной, клиновидной, веретеновидной, цилиндрической, червеобразной, тетраэдрической, трех- или многоугольной, лопастной, почковидной; чашевидной, серповидной, бубликовидной, звездчатой и др. Многообразие внешнего вида клеток умножается благодаря разнообразной скульптурированности клеточной оболочки (в виде ячеистой или сетчатой скульптуры, выпячиваний, утолщений, бородавок, шипов, шипиков, щетинок, зубчиков, палочковидных, сосочковидных или иной формы выростов). У прикрепленных форм оболочка обычно образует вырост - ножку с прикрепительным диском (подошвой).
Обычно оболочка плотно прилегает к протопласту по всей поверхности. Иногда она ослизнена и далеко отстоит от протопласта (Merismogloea Pasch.) или имеет многочисленные поры (Sklerochlamys fragilis Pasch.). Иногда клетки полностью покрыты бесструктурной или слоистой слизью, объединяющей их в колонии (Gloeobotrydaceae Pasch.), либо находятся на концах слизистых ветвящихся тяжей, образующихся в результате ослизнения оболочек или одностороннего выделения слизи протопластами клеток (Мilleodendron Pasch., Mischococcus Näg.).
Обычно клетки одноядерны. С возрастом количество ядер может увеличиваться (Perone Pasch., Exentrochloris Pasch.). Число ядер возрастает также при подготовке к размножению (Botrydiopsis Borzi, Chlorocloster Pasch.). Многоядерность свойственна плазмодиям Myxochloris Pasch. и Chlamydomyxa Arch., а также сифональным талломам Botrydium и Vaucheria.
Как и у бурых водорослей, наружная мембрана ядер ной оболочки у многих (но не у всех) видов переходит в канал эндоплазматического ретикулюма, окружающий хлоропласты. У представителей класса Eustigmatophyceae канал эндоплазматического ретикулюма, окружающий хлоропласт, не связан с ядром.
Митоз изучен у немногих видов, и неизвестно, окажутся ли описанные картины типичными для всего отдела. Во время деления ядерная оболочка остается интактной, ядрышко фрагментируется; веретено деления центрического типа, на полюсах его располагаются центр ноли. Между дочерними ядрами сохраняется межзональное веретено. Кинетохоры не обнаружены.
Хлоропласты разнообразны по форме и тонкому строению. Они бывают дисковидными, пластинчатыми, корытовидными, кольцевидными, цилиндрическими, лентовидными, звездчатыми, чашевидными. Края их ровные, лопастные или глубоко рассеченные на дольки, которые иногда отделяются, образуя несколько мелких хлоропластов. Лишь изредка хлоропласты занимают в клетке центральное положение (Heterogloea minor Pasch., Gloeoskene turfosa Fott), обычно они располагаются пристенно одним или несколькими слоями. Число их колеблется от одного до нескольких сот в одной клетке и может изменяться в онтогенезе; чаще всего их несколько.
Хлоропласты окружены каналом эндоплазматического ретикулюма, который вместе с оболочкой хлоропласта образует четырехмембранную систему. В строме хлоропласта продольно располагаются ламеллы, состоящие из трех параллельных дисков каждая. На периферии пластиды наблюдается одиночный огибающий диск, хотя иногда (у представителей класса Eustigmatophyceae) он может отсутствовать. По тонкой структуре хлоропласты желтозеленых напоминают пластиды золотистых, диатомовых и бурых водорослей.
Комплекс фотосинтетических пигментов своеобразен. Он включает хлорофиллы а и с, хлорофилл b отсутствует. Из каротиноидов отмечены β- и ξ-каротины. В качестве основного ксантофилла выступает вошериаксантин. У представителей класса Xanthophyceae ксантофилловый цикл осуществляют диадиноксантин - диатоксантин, к которым присоединяется гетероксантин, в то время как у Eustigmatophyceae в ксантофилловом цикле участвуют преимущественно виола ксантин антераксантин зеаксантин. Характерный для бурых, золотистых и диатомовых водорослей коричневый пигмент фукоксантин у желтозеленых отсутствует. Наряду с окрашенными наблюдаются бесцветные недостаточно изученные формы.
Пиреноиды наблюдаются редко. Они бывают двух типов: полу погруженными, с проходящими сквозь них многочисленными ламеллами, являющимися прямым продолжением пластидных (у представителей класса Хаnthophyceae: Miscnococcus, Bumilleriopsis Printz, Botrydium) и выступающими, сидящими на выросте внутренней поверхности хлоропласта, имеющем форму подушечки или тоненькой ножки, без ламелл (у представителей класса Eustigmatophyceae: Polyedriella helvetica Visch. et Pasch., Pseudocharaciopsis texensis Lee et Bold). В обоих случаях пиреноиды "голые", не покрытые ассимиляционной обкладкой.
Продукты ассимиляции - внепластидные масла, жиры, хризоламинарин, белковые кристаллы; наблюдаются также сильно преломляющие свет кристаллы неизвестной природы. Хризоламинарин, отлагающийся главным образом в покоящихся клетках, представляет собой β-1,3(1,6)-полиглюкан, идентичный с обнаруженным у Chrysophyta и Bacillariophyta и близкий к ламинарину бурых водорослей.
У подвижных монадных и амебоидных форм и стадий, а также у некоторых гемимонадных водорослей наблюдается стигма красного, коричневатого или оранжево-красного цвета, округлой, эллипсоидной или линейной формы, точно сориентированная с базальным утолщением на заднем рулевом жгутике, предположительно выполняющим функцию фоторецептора. У представителей класса Xanthophyceae стигма является частью хлоропласта, располагаясь на его переднем конце, под его оболочкой, и состоит из одного слоя осмиофильных капель. В зооспорах Eustigmatophyceae обнаружена крупная внепластидная стигма, состоящая из осмиофильных глобул разных размеров, не окруженных мембранами.
Монадные формы и стадии обладают двумя гетероконтными, гетероморфными, гетеродинамичными жгутиками (рис. 16.3, 1, 2). Один из них, обычно более длинный, перистый, при движении направлен вперед (главный, передний, или "плавательный", жгутик), второй, обычно более короткий, гладкий - назад (побочный, задний, или рулевой, жгутик). Длина главного жгутика в 1,5-8 раз превышает длину побочного; последний может быть полностью редуцированным (Nephrochloris incerta Geitl. et Gim.). У Pedinomonadopsis minor Massjuk et Guk соотношение длин жгутиков обратное: передний очень короткий, едва заметный, задний - длинный [200]. Оба жгутика выходят из апикальной, субапикальной или вентральной жгутиковой ямки. Передний жгутик несет два ряда трубчатых мастигонем, каждая из которых заканчивается двумя длинными и тонкими, одинаковой или неравной длины терминальными волосками. Наличие мастигонем существенно влияет на направление движения клетки. У основания заднего жгутика находится вздутие - парабазальное тело, расположенное в мелком углублении на поверхности клетки, как раз над стигмой.

Рис. 16.3. Основные типы строения жгутикового аппарата желтозеленых водорослей: 1 - Heterochloris Pasch.; 2, 3 - Vaucheria D. C. (2 - антерозоид, 3 - фрагмент синзооспоры); 4 - зооспора Eustigmatophyceae (а - парафлагеллярное тело; б - стигма)
Жгутиковая аксонема имеет типичную для всех эукариот структуру "9 + 2". Базальные тела, как обычно, содержат 9 триплетов микротрубочек. В переходной зоне выявлена характерная спиралевидная электронноплотная структура, состоящая из 3-4 витков [467, 470], напоминающая таковую у Chrysophyta [466, 467, 469]. От базальных тел отходит два типа корней: единственный поперечно-полосатый тонкий фиброзный корень (ризопласт), тесно прилегающий к внутренней поверхности переднего выроста ядра, и 3 микротрубчатых корня из 3-4 микротрубочек каждый, расходящихся в цитоплазме в разных направлениях.
Синзооспоры и антерозоиды Vaucheria D. С. обладают разными типами строения жгутикового аппарата (см. рис. 16.3, 2, 3). В синзооспоре все многочисленные жгутики почти равной длины, гладкие. Антерозоид имеет одну пару гетероморфных, гетероконтных, гетеродинамичных жгутиков и, кроме того, "хоботок", который содержит кольцо из 8-9 микротрубочек, берущих начало от базальных тел, и который напоминает аналогичное образование сперматозоидов Fucus L.
Зооспоры Eustigmatophyceae имеют один, реже два неравных жгутика с парабазальным телом, расположенным у основания главного, более длинного жгутика, имеющего план строения "9 + 1" и покрытого трубчатыми мастигонемами.
Монадные, гемимонадные и амебоидные формы и стадии имеют сократительные вакуоли; лишь у морских и солоноводных видов они отсутствуют. Число их в клетке у разных видов колеблется от 1-2 до многих. В одетых оболочками клетках коккоидных, нитчатых или псевдопаренхиматозных водорослей сократительные вакуоли могут наблюдаться лишь на ранних стадиях развития таллома. Во взрослых клетках они отсутствуют, а вместо них появляется центральная вакуоль с клеточным соком, образующаяся в результате слияния многих более мелких вакуолей. В делящихся клетках наблюдаются многочисленные мелкие выделяющие слизь тельца.
Размножение желтозеленых водорослей осуществляется преимущественно бесполым путем с помощью вегетативных или специализированных клеток. Бесполое (вегетативное) размножение одноклеточных, лишенных клеточной оболочки, происходит делением клеток надвое (Rhizochloris Pasch., Ankylonoton Pasch.), колониальных (неценобиальных) и многоклеточных - фрагментацией таллома или отчленением отдельных конечных клеток таллома (Heteropedia polychloris Pasch.). У Heterococcus gemmatus (Snow) Bourr. наблюдается образование боковых пальцевидных одно- или многоклеточных отростков, которые, отчленяясь от материнского растения, дают начало новым растениям. Вегетативное размножение Vaucheria D. С. осуществляется благодаря хорошо выраженной регенерационной способности. При отмирании части таллома оставшиеся жизнеспособными участки отделяются перегородками от отмерших или поврежденных участков, и затем регенерируют в целое растение. Кроме того, у вошерии наблюдается образование выводковых почек [604]. У нитчатых желтозеленых известны акинеты [405] (см. рис. 16.6).
Наиболее широко у желтозеленых водорослей представлено бесполое размножение с помощью специализированных клеток: амебоидов, зооспор и синзооспор, гемизооспор и гемиавтоспор, автоспор и синавтоспор (см. рис. 16.5), апланоспор (см. рис. 16.5) и синапланоспор, гипноспор. Размножение амебоидами наблюдается редко, преимущественно у водорослей, обладающих амебоидной структурой вегетативного тела (Stipitococcus vas Pasch.). Обычно, кроме амебоидов, в цикле развития известны и зооспоры (Stipitococcus vas Pasch., Perone dimorpha Pasch.). Нередко зооспоры некоторых видов (Tribonema viride Pasch., T. intermixtum Pasch., Pleurochloridella vacuolata Pasch., Botrydiopsis arhiza Borzi), сбрасывая жгутики, переходят в амебоидное состояние (см. рис. 16.5, 1, г, д).
Наиболее распространенный способ размножения связан с образованием зооспор. Обычно зооспоры имеют дорсивентральную форму, одно ядро, два гетероконтных, гетероморфных и гетеродинамичных жгутика, стигму, сократительные вакуоли, один хлоропласт (реже их несколько) (см. рис. 16.5, 2, г; рис. 16.6, 1, в, 3, в, 4, в, 7, в). Иногда форма зооспор может быть иной, например широковеретеновидной (Pleurochloris polyphem Pitsch.), один из жгутиков может быть редуцирован до полного исчезновения (Gloeochloris planctonica Pasch., Pleurochloris magna Boye-Pet.), стигма может отсутствовать (Botrydiopsis eriensis Snow) (см. рис. 16.6, 2, в).
Многоядерные зооспоры Vaucheria, со многими парами изоморфных гладких жгутиков почти равной длины и многочисленными хлоропластами получили название "синзооспоры" (см. рис. 16.7, 3-5). Случайное образование синзооспор отмечено также у видов родов Botrydiopsis, Tribonema, Bumilleria Borzi, Heterococcus и др. У некоторых желтозеленых водорослей (Botrydiopsis) наблюдается образование недоразвитых зооспор, лишенных плотных клеточных покровов и жгутиков, с сократительными вакуолями и стигмой, получивших название "гемиавтоспорность" [403, 405]. В цикле развития многих видов, особенно с коккоидной структурой тела, наблюдается образование автоспор (Botrydiopsis, MonodusChod., Chlorocloster, Tetraplektron Fott и др.). Размножение апланоспорами свойственно многоклеточным - нитчатым, разнонитчатым и псевдопаренхиматозным водорослям; у представителей с сифональной структурой (Botrydium, Vaucheria) наблюдается образование синапланоспор.
Ультраструктурное изучение апланоспорогенеза у Vaucheria longicaulis Норр. var. macounii Blum показало, что апланоспоры у этого рода не являются потенциальными зооспорами, отличаясь от последних рядом существенных признаков [410]. У ряда видов (Botrydiopsis antarctica Kol, Chlorocloster pachychlamys Pasch., Monodus pyreniger Pasch. и др.) образуются покоящиеся споры, или гипноспоры, с толстой, нередко слоистой, насыщенной кремнеземом, иногда шиповатой, двустворчатой и (или) окрашенной в темный цвет оболочкой и с богатым запасом питательных веществ в протопласте. Разнообразные типы спор образуются в одноклеточных спорангиях, не отличающихся от обычных вегетативных клеток или превышающих их по своим размерам.
Наряду с акинетами и гипноспорами, одновременно выполняющими репродуктивную и защитную функции, наблюдается образование других стадий покоя, например, эндогенных цист (рис. 16.4) с плотной блестящей насыщенной кремнеземом двустворчатой оболочкой, прорастающих зооспорами или амебоидами (Chloramoeba heteromorpha Bohl., Heterogloea endochloris Pasch., Chloromeson agile Pasch. и др.). Наряду с типичными одноядерными наблюдаются многоядерные цисты (Myxochloris sphagnicola Pasch.). У видов рода Botrydium Wallr. весь протопласт может превращаться в крупную многоядерную макроцисту или распадаться в подземной части слоевища на ряд более мелких ризоцист. Иногда желтозеленые водоросли переживают неблагоприятные условия в пальмеллевидном состоянии (Neonema quadratum Pasch.). Если слизь, объединяющая водоросли, имеет концентрическую слоистость и состоит из системы вложенных друг в друга пузырей, такие образования называют глеоцистами [403].
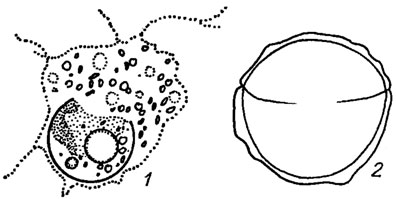
Рис. 16.4. Эндогенные цисты желтозеленых водорослей: 1 - Myxochloris sp.; 2 - Chloromeson agile Pasch
Половой процесс (оогамного типа) с достоверностью известен лишь у видов рода Vaucheria*. Половые органы - антеридии и оогонии - образуются либо на одном и том же растении (однодомные; см. рис. 16.7, 10), либо на разных (двудомные). Образующиеся в антеридии двужгутиковые антерозоиды, проникая в оогоний, оплодотворяют яйцеклетку. Зигота, или ооспора, покрывается толстой многослойной оболочкой и после периода покоя прорастает в новое растение. Хотя зиготический мейоз с точностью не доказан, считают, что цикл развития вошерии гаплофазный [604].
* (Сведения об изогамном половом процессе у видов родов Botrydium, Tribonema и Monodus нуждаются в подтверждении [207].)
Цикл развития остальных Xanthophyta, у которых отсутствует половой процесс, проходит без смены ядерных фаз. Иногда он простой и состоит из вегетативной и репродуктивной фазы, представленной одним из известных способов бесполого размножения (Rhizochloris Pasch., Stipitoporos Ettl, Chloridella Pasch. и др.). Однако нередко цикл развития усложнен наличием у одного и того же вида различных форм бесполого размножения, сочетание и последовательность которых может варьировать в широких пределах. Поливариантность циклов развития свойственна видам Tribonema Derb. et Sol., Botrydium Wall., Myxochloris Pasch. и др.
На основании большого морфологического сходства с зелеными водорослями, желтозеленые до 1899 г. относили к Chlorophyta в качестве класса Heterocontae наряду с Isocontae, Acontae, Stephanocontae. В 1899 г. они были выделены А. Лютером в самостоятельную группу под тем же названием Heterocontae [120]. Однако название "разножгутиковые", обоснованное как противопоставление "равножгутиковым" пока их включали в состав зеленых водорослей, потеряло свой смысл, когда их отделили от зеленых, так как разные жгутики имеют также золотистые, бурые и другие водоросли. В 1930 г. им было присвоено новое название Xanthophyceae. А. Пашер [576, 579] впервые обосновал родство желтозеленых с золотистыми и диатомовыми водорослями, объединив их в одну филу (ствол) под названием Chrysophyta. Особое значение он придавал наличию у этих групп эндогенных цист, оболочки которых пропитаны кремнеземом. Впоследствии эта точка зрения была подтверждена сравнительными биохимическими и цитологическими исследованиями, установившими ряд других общих для них признаков: отсутствие хлорофилла b и наличие хлорофилла с, отсутствие крахмала и наличие хризоламинарина, особенности тонкой структуры клетки, в первую очередь хлоропластов и др. [348, 392]. Поэтому точка зрения А. Пашера принимается многими авторами [54, 356, 371, 374, 403, 405, 413, 414, 417, 483, 484, 653 и др.], объединяющими Xanthophyceae, Chrysophyceae и Bacillariophyceae (иногда к ним присоединяют также Phaeophyceae) в отделе Chromophycophyta [371], либо Chrysophyta [356, 405, 413, 414], Chromophyta [417, 484], Heteroconthophyta [584] или в подцарстве Chrysobionta [54]. В отечественной литературе [99, 207] желтозеленые рассматриваются как самостоятельный отдел Xanthophyta. Д. К. Зеров, хотя и подчеркивал родство желтозеленых с золотистыми, диатомовыми и более отдаленное - с бурыми водорослями, оставляет за всеми этими группами статус отдела.
Данные биохимических [378, 451, 497, 664, 694-696, 697 и др.] и электронно-микроскопических [352, 384, 465-468, 474-478] исследований привели к выводу о гетерогенности желтозеленых водорослей [470-474, 476, 478]. Предложено выделить из их состава ряд видов с коккоидной организацией вегетативного тела, отличающихся своеобразными ультраструктурными и биохимическими признаками, в отдельный класс Eustigmatophyceae [472, 474] или отдел Eustigmatophyta [472]. Уникальное сочетание химических и ультраструктурных признаков у Chlorarachnion reptans Geitl. послужило основанием для выделения его в самостоятельный класс Chlorarachniophyceae и отдел Chlorarachniophyta [478].
Классификация желтозеленых водорослей на уровне порядков со времен А. Пашера [577, 581] базируется на морфологических признаках (тип структуры вегетативного тела). Эти же признаки другими авторами [207, 585] используются для разграничения классов. Использование одних и тех же признаков на различных таксономических уровнях привело к тому, что таксономический ранг одних и тех же таксонов желтозеленых водорослей (включая наивысшие) в системах разных авторов неоднозначен. Еще одним источником нестабильности системы желтозеленых является дискуссионность вопроса об исходных первичных организмах и в связи с этим - о месте в системе монадных и ризоподиальных организмов. В настоящем издании принято предложение о выделении класса Eustigmatophyceae [472]; система Xanthophyceae дана по X. Эттлу [403].
Желтозеленые распространены в воде, почве, в наземных местообитаниях (включая толщу воздуха), на разных географических широтах, на всех континентах земного шара. Большинство видов - пресноводные организмы, и лишь немногие принадлежат к солоноводным (Rhizolekane sessilis Pasch.) и морским (Rhizochloris mirabilis Pasch., Nephrochloris salina Cart.). Некоторые обитают в водоемах с необычным солевым составом, например, Chlorocardion sal inarum Komaromy - в выщелоченном натриевом растворе урановых руд. Они встречаются в водоемах с различной степенью трофности, загрязненности органическими остатками, при различных концентрациях водородных ионов, однако предпочитают чистые воды стоячих водоемов с кислой реакцией среды. Чаще всего это сфагновые и торфовые болота, канавы, ямы и лужи на торфяниках, заболоченные лесные и луговые лужи и канавы, пруды, речные старицы и запруды, водохранилища и озера, реже - водоемы с проточной водой. Некоторые виды развиваются в сточных и загрязненных водах водоемов системы биологической очистки (Centritractus belonophorus Lemm., Ophiocytium capitatum Wolle, Goniochloris mutica (A. Br.) Fott и др.).
Желтозеленые - преимущественно эвритермные организмы, предпочитающие теплые сезоны года, но есть виды, развивающиеся при низких температурах (Ellipsoidion simplex Pasch., Monodus subglobosa Pasch., Gloeochloris planctonica Pasch., Mischococcus confervicola Näg., Ophiocytium arbuscula (A. Br.) Rabenh. и др.), а также типичные крисфилы (Chloridella glacialis Kol, Ellipsoidion perminimum Pasch. var. cryophila Kol). Термофилы среди желтозеленых не выявлены.
В водных биотопах желтозеленые обитают преимущественно в толще воды, или входят в состав перифитона. Бентосных и нейстонных форм известно немного. Разнообразие, численность и биомасса их в водных ценозах обычно незначительны. Лишь изредка в связи с эвтрофикацией водоемов численность желтозеленых возрастает, и они выходят на доминирующее положение в биоценозе, вызывая "цветение" воды (Botrydiopsis arhiza Borzi).
Значительно разнообразнее и обильнее представлены желтозеленые в почвах разных типов, где они могут составлять до 50% общего числа видов и нередко при массовом развитии вызывают "цветение" поверхности почвы. Наиболее распространенными во всех растительно-климатических зонах и типах почв СССР являются Botydiopsis arhiza Borzi, В. eriensis Snow, Heterothrix bristoliana Pasch., H. exilis (Klebs) Pasch., Bumilleria sicula Borzi, виды родов Pleurochloris Pasch., Polyedriella Pasch., Monallantus Pasch., Monodus Chod., Ellipsoidion Pasch., Botrydium Wallr., Vaucheria D. C. [85, 327]. Аэрофитные желтозеленые водоросли поселяются на стволах деревьев, скалах, стенах домов, вызывая иногда их позеленение [200].
Разнообразны формы взаимоотношений желтозеленых с другими организмами. Эпифиты поселяются на других водорослях (Peroniella eudorinae Skv., P. hyalothecae Gobi, Characiopsis anabaenae Pasch., Ch. microcysticola Skuja и др.), высших водных растениях (Chadefaudiothrix minouchetii Bourr., Peroniella curviceps Pasch.) и сфагновых мхах (Perone dimorpha Pasch.), эпизоиты - на корненожках, ракообразных и коловратках (Characiopsis diffugicola Hub.-Pest., Ch. rossica (Elenk.) Deduss., Ch. saccata Cart.). Эндофиты произрастают в водоносных клетках сфагновых мхов (Myxochloris sphagnicola Pasch.) или интеркалярно в листьях и черешках ароидных (Phyllosiphon Kuhn), иногда вызывая галообразование на вегетативных органах у представителей Utricaceae и Solanaceae (Phytophysa Boss., Chlorocloster solani George). Известны случаи внутриклеточного симбиоза ("зооксантеллы" в клетках Protozoa). В целом сравнительно небольшой по числу видов отдел Xanthophyta отличается широкой экологической амплитудой.
Желтозеленые водоросли играют определенную роль в природе как продуценты кислорода и органических веществ, агенты самоочищения загрязненных вод и почв; они входят в состав первичного звена трофических цепей, принимают участие в почвообразовательных процессах, в создании почвенного плодородия. Хозяйственное значение желто-зеленых определяется их использованием в качестве показательных организмов в системе экологического мониторинга, в составе комплекса микроорганизмов, осуществляющих очистку сточных вод.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'