14. Отдел 5. Золотистые водоросли - Chrysophyta (Chrysophycophyta, Chrysophyceae) (З. И. Ветрова)
Золотистые, или хризофитовые, водоросли представлены одноклеточными, колониальными или многоклеточными организмами золотисто-желтого, буровато-зеленого, реже зеленовато-желтого цвета. Имеются бесцветные формы. В пределах отдела насчитывается около 800 видов.
Большинство индивидов имеет монадный тип структуры тела, только незначительное число форм имеют амебоидную, плазмодиальную*, пальмеллоидную, коккоидную, нитчатую, разнонитчатую и пластинчатую структуру. Монадные формы и стадии имеют 1-2, реже 4 жгутика, различных по строению и длине, реже последние одинаковые.
* (Плазмодиальная структура соответствует амебоидной в разделе 1.3, пластинчатая - паренхиматозной либо псевдопаренхиматозной.)
Форма клеток различная, преобладает яйцевидная, эллипсоидная, веретеновидная, цилиндрическая, реже встречаются шаровидные, грушевидные, булавовидные, кубические и другие клетки (рис. 14.1; 14.2).
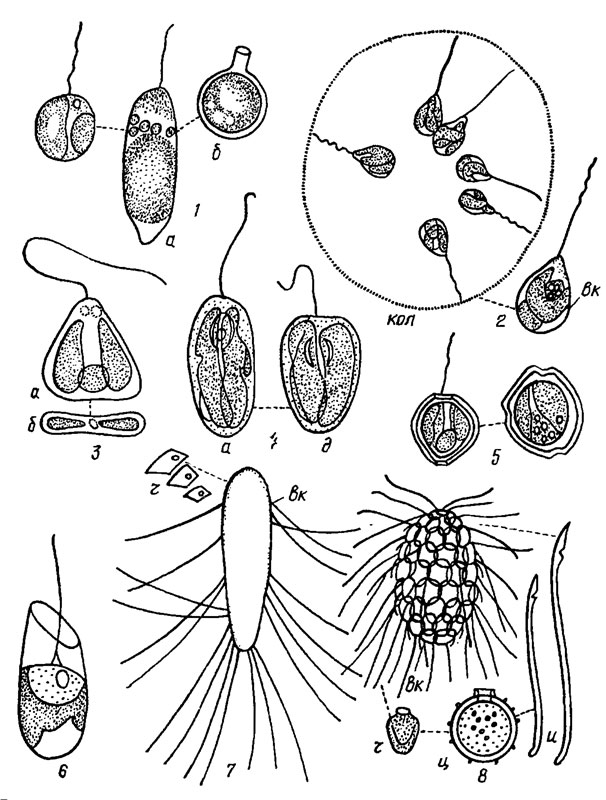
Рис. 14.1. Золотистые водоросли порядка Chromulinales (семейство Chromulinaceae): 1 - Chromulina rosanoffii (Woronin) Butschli (a - вид сбоку, б - циста); 2 - Saccochrysis pyriformis Korsch.; 3 - Sphaleromantis ochracea (Butschli) Pasch. (a - вид спереди, б - поперечный срез); 4 - Amphichrysis compressa Korsch. (a - вид сбоку, б - вид спереди); 5 - Chrysococcus trlporus Matv. (клетка в разных положениях); 6 - Kephyrion ovum Pasch.. 7 - Mallomonas producta Iwan.; 8 - M. acaroides Perty; вк - вегетативная клетка, ц - циста, кол - колония, ч - чешуйки, и - иглы
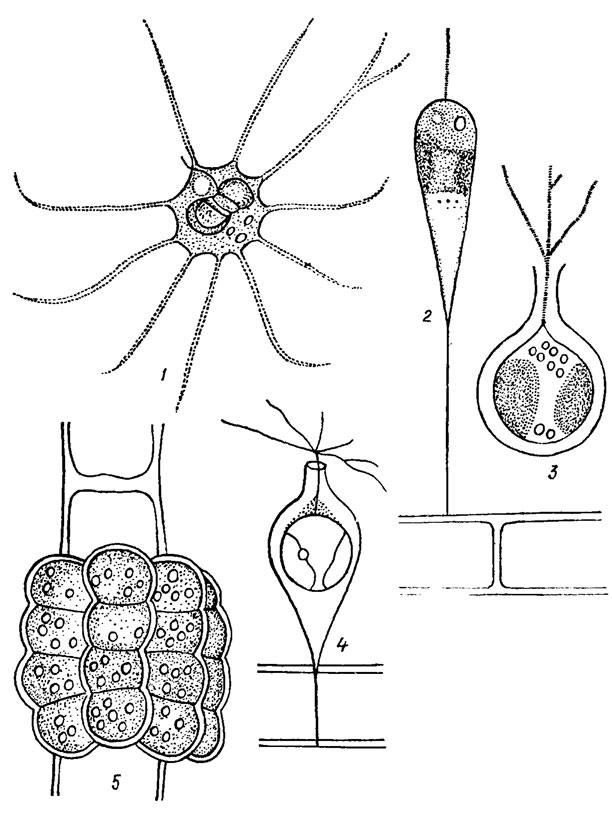
Рис. 14.2. Золотистые водоросли порядка Chrysamoebales (семейства Chrysamoebaceae и Chrysocapsaceae): 1 - Chrysamoeba tenera Matv.; 2 - Stipitochrysis monorhiza Korsch.; 3 - Laginion ampullaceum (Stokes) Pasch.; 4 - Chrysopyxis urna Korsch.; 5 - Epichrysis piludosa (Korsch.) Pasch
Размеры клеток колеблются в пределах 2-120 мкм. Наиболее мелкие клетки имеют представители родов Chromulina Cienk. (Ch. minima Dofl. - 2-3 мкм, Ch. elegans Dofl. - 3-3,5 мкм в длину) и Stenokalyx Schill (S. parvula Schmidt. - 4-5 мкм в длину). Наиболее крупные клетки наблюдаются у родов Mallomonas Perty (М. insignis Penard - 85-100 мкм, М. cylindracea Pasch. - 50-120 мкм в длину и Microglena Ehr. (-М. punctifera Mull.) Ehr. - до 50 мкм в длину). Имеются значительные колебания размеров клеток и в пределах одного и того же вида как одноклеточных (Mallomonas akrokomos Ruttn. - 8-62 мкм в длину), так и колониальных форм (Uroglena proxima Korsch. et Matv. - 9-19,5 мкм в длину).
Клетки в большинстве случаев лишены клеточной оболочки, иногда покрыты лишь плазмалеммой, часто также мелкими кремнеземными чешуйками, нередко срастающимися в панцирь, реже выделяют домики (Chrysococcus Klebs, Hyalobryon Laut.). Некоторые представители имеют целлюлозную или целлюлозно-пектиновую, бесцветную или окрашенную оболочку, иногда покрытую многочисленными известковыми тельцами - кокколитами (Hymenomonas Stein), или кремнеземными чешуйками, а также инкрустированную солями железа (Chrysotilos ferrea Pasch.). Некоторые формы обладают особыми стрекательными структурами - дискоболоцистами, организованными иначе, чем трихоцисты динофлагеллат.
Домики бывают шаровидные или яйцевидные, реже полушаровидные, эллипсоидные, колбовидные, вазовидные, цилиндрические и др. Размеры их разные и колеблются в пределах от 2 мкм (Chrysococcus punctiformis Pasch.) до 70 мкм (Hyalobryon ramosum Laut.). Стенки домиков состоят из клетчатки, пропитанной углекислой известью и солями железа; последние окрашивают домики в коричневый цвет (Eleutheropyxis fulva Scherff.). У некоторых видов домики окружены слизью (Chrysocrinus hydra Pasch.) или зернистыми образованиями (Heliochrysis eradians Pasch.). Стенки домиков могут быть пропитаны и кремнием.
Домики могут иметь одно крупное отверстие или таких отверстий 2-11 и даже больше. Они расположены или на концах или по всей поверхности домика, а также по экватору. У некоторых видов вокруг отверстия образуется короткая или длинная шейка, трубочка, воротничок или другой формы выросты стенок домиков. Поверхность домиков бывает гладкая или орнаментированная единичными или несколькими выростами или утолщениями. У представителей рода Hyalobryon Laut. наблюдаются воронкообразные образования, т. н. кольца нарастания. Домики прикрепляются к субстрату подошвой, окрашенной часто солями железа в бурый цвет и поэтому хорошо заметной (Stephanoporos scherffelii Pasch.). Нередко прикрепление осуществляется с помощью хорошо заметной толстой слизистой ножки (Lepochromulina calyx Scherff.) или тонкого стебелька (Rhizaster crinoides Pasch.), пропитанных солями железа. Клетки в домиках к стенкам либо не прикрепляются (Dinobryon Ehr , Hyalobryon), либо прикрепляются к внутренней стенке длинными стебельками.
Панцирь состоит из значительного числа кокколитов или кремнистых скорлупок, реже кремнистых кольцевидных ободков. Строение панциря имеет важное систематическое значение. Скорлупки панциря бывают яйцевидные, эллиптические, округлые, ромбические, лопатовидные, в виде часового стекла и подковы. Как правило они симметричные, гетеропольные, или изопольные, реже асимметричные (Mallomonas robusta Matv.). У многих из них имеется особый придаток в виде подковы, служащий для соединения скорлупок в сплошной панцирь. Этот придаток становится заметным только после специальной обработки панциря. У некоторых видов скорлупки панциря имеют подковообразные (Mallomonas acaroides Perty), пальцевидные (Mallomonas clavata Conr.), или кольцевидные (Synura sphagnicola Korsch.) утолщения. Иногда скорлупки снабжены выростами в виде шипов, иголок или щетинок различной длины, тупыми или острыми, прямыми или косо направленными, тонкими или толстыми. Размеры скорлупок до 10 мкм в длину и 7,5 мкм в ширину; иногда они очень мелкие (0,8 мкм в длину, 0,5 мкм в ширину) и обнаружить их можно только при электронно-микроскопическом исследовании (Chrysochromulina).
Скульптура скорлупок очень разнообразна. Их форма, размеры, количество и характер расположения используются в систематике для идентификации видов. На скорлупках очень часто встречаются различных размеров отверстия и поры, а также углубления, служащие для прикрепления шипов и иголок, соединения скорлупок в пределах панциря. Размещение скорлупок неодинаково. Как правило многочисленные скорлупки образуют сплошной панцирь (Mallomonas), реже скорлупки покрывают только переднюю часть клетки (Synura splendida Korsch.). Наиболее часто скорлупки панциря расположены косыми рядами в виде черепицы, реже прямыми рядами в виде кирпичной кладки (Mallomonas caudata Iwan.).
Кокколиты известны у нескольких родов, выделенных даже в отдельный подкласс [204]. К ним относятся преимущественно морские формы. Кокколиты также имеют разнообразную форму и мелкие размеры. У одних видов они расположены равномерно по всей клетке на определенном расстоянии друг от друга, у других тесно соединяются своими краями (Acanthoica ornata).
Некоторые представители образуют ризоподии, псевдоподии и аксоподии. Ризоподии - длинные тонкие образования на поверхности клеток в виде простых или разветвленных выпячиваний, достигающих значительной длины (до 140 мкм). Расположены они или равномерно по всей поверхности (Chrysamoeba Klebs) или локализуются в определенных участках клетки (Rhizaster crinoides Pasch.). У представителей, живущих в домиках, ризоподии выходят наружу через одну, несколько или много пор в стенках домика (Derepyxis Stokes, Porochrysis Pasch., Chrysocrinus). У некоторых видов ризоподии служат для прикрепления клеток к субстрату (Amphirhiza epizootica Skuja). Кроме участия в передвижении клеток, ризоподии принимают участие в голозойном питании путем захвата твердых органических частиц, бактерий и даже водорослей. Псевдоподии - короткие широкоокруглые выпячивания клеток, хорошо выраженные у Brehmiella chrysohydra Pasch. К псевдоподиям, по мнению А. М. Матвиенко [204], следует также отнести и губовидные выпячивания, наблюдающиеся у некоторых хромулин (Chromulina pascheri Holer.) и многих охромонасов (Ochromonas charkowiensis Matv., О. coronifera Matv. и др. Назначение псевдоподий такое же, как и ризоподий. Аксоподии - выпячивания с уплотненной осевой нитью, окруженной тонким слоем цитоплазмы. Внешне они напоминают щупальца животных и встречаются у видов семейства Pedinellaceae. Расположены они кругом. Число, длина и толщина их всегда постоянны для одного и того же вида. В определенных условиях аксоподии могут быть втянутыми и незаметными.
Преимущественное большинство золотистых водорослей имеют жгутики. Наиболее часто встречаются одножгутиковые или двужгутиковые формы с жгутиками разной длины. Известны роды с тремя жгутиками, причем все они разной длины, как у Prymnesium Mass., Platychrysis Geitl., или два из них одинаковы, а третий значительно длиннее (Chrysochromulina). Четырехжгутиковые формы имеются у представителей рода Didymochrysis Pasch. (Didymochrysis paradoxa), причем жгутики образуют две "разножгутиковые" пары. У представителей с одним или двумя неравными жгутиками длинный называется главным, более короткий - боковым. У видов с тремя разными жгутиками один из них, наиболее короткий, служит для приклеивания к субстрату (гаптонема), или он длинный и принимает участие в захвате твердых органических частиц. Если жгутик один, как, например у хромулины, он имеет перистое строение, если два, то один из них обычно перистый, другой бичевидный (Ochromonas Wyssotz., Synura Ehr.). У некоторых родов (Chrysochromulina) оба жгутика бичевидные и равной длины [120]. Во время деления клеток или перед образованием цист жгутики могут или втягиваться, или сбрасываться. Втягивание характерно для представителей, не имеющих плотных покровов.
Ядро обычно одно, шаровидное или слегка эллипсоидное, расположено в передней части клетки или в центре, реже в задней части. Размеры его незначительны. Часто оно маскируется капельками масла. Наиболее крупное ядро у представителей рода Mallomonas; оно может составлять 1/3 клетки (Mallomonas denticulata Matv.), достигая 10 мкм в диаметре (М. mesolepis Skuja var. spinosa Matv.). Ядрышко одно. Особенностью митоза является то, что ядерная оболочка сначала фрагментируется, а затем исчезает. Веретено деления имеет типичный вид, но центриоли отсутствуют, вместо них на полюсах расположен ризопласт.
Хлоропласты обычно пластинчатые, корытовидные, реже лентовидные с неровным волнистым или ровным краем, дисковидные, сетчатые, звездчатые и другой формы. Обычно в клетке находится 1-2, реже 4 (Amphichrysis compressa), 8 (Epichrysis nitellae Geitl.) и более (Pseudosyncrypta volvox I. Kiss.). Пигменты представлены хлорофиллами а и с, β-каротином и ксантофиллами (антераксантином, лютеином, зеаксантином, неоксантином, виолаксантином, фукоксантином, диадиноксантином, диатоксантином, диноксантином и криптоксантином). В хлоропластах находятся пиреноиды (обычно голые). У Ochrosphaera neapolitana обнаружен стебельчатый пиреноид, расположенный вне хлоропласта и соединенный с ним особым стебельком, т. н. пиреноидоносителем.
Стигма является частью хлоропласта, расположена на его переднем крае, непосредственно под боковым жгутиком, несущим фоторецептор. Обычно она округлая, округло-овальная или в виде палочки, часового стеклышка (Chromulina urophora Skuja) или червячка (Microglena ovum Schill.). Размеры стигмы обычно незначительны, но встречаются виды с крупной и хорошо заметной стигмой (Chrysoglena verrucosa Wisl.). Форма, размеры и расположение стигмы используются в систематике этих водорослей. Ультраструктура этой органеллы изучена недостаточно.
Вакуолярный аппарат состоит из пульсирующих (сократительных) или непульсирующих вакуолей-пузул. Сократительные вакуоли небольшие, округлые по 2-5, реже по 1 в клетке, с тонкой оболочкой. У видов с более плотной оболочкой система вакуолей образует сложный аппарат, состоящий из 5-7 (Hymenomonas) или 4-8 (Microglena Ehr.) мелких и одинаковых по форме и размерам сократительных вакуолей, расположенных вокруг одной пузулы. Размещены вакуоли у основания жгутиков, нередко в средней или даже в задней части клетки. Однако сложный аппарат вакуолей всегда расположен в передней части клетки.
Продукты ассимиляции золотистых водорослей - внепластидные масла и хризоламинарин. Откладываются они в задней части клетки, реже заполняют всю клетку. Имеются сведения о наличии у некоторых золотистых волютина и гликогена (Monochrysis aphanaster Skuja, Ochrosphaera neapolitana). У некоторых видов рода Synura в передней части клетки накапливается гематохром [204].
У золотистых известно бесполое размножение (вегетативное или с помощью специализированных клеток - амебоидов, зооспор и апланоспор) и половое. Вегетативное размножение широко представлено у одноклеточных форм. Осуществляется оно путем простого деления клетки надвое. У амебоидных форм деление происходит в подвижном состоянии, без предварительного втягивания ризоподий (Chrysamoeba tenera Matv., Brehmiella chrysohydra Pasch., Chrysarachnion insidians Pasch. и др.). Вегетативное размножение с плазмодиальной морфологической структурой (Myxochrysis paradoxa Pasch.) заключается в расщеплении плазмодия на много одноядерных и реже многоядерных амеб, которые затем вырастают во взрослый плазмодий. У монадных форм размножение происходит тоже в подвижном состоянии, но значительно медленнее. У многоклеточных нитчатых, разнонитчатых и пластинчатых форм разрастание слоевища может происходить за счет деления клеток в одном (Nematochrysis Pasch.), двух (Chrysomeris Cart.) и трех (Phaeodermatium Hansg.) направлениях. Слоевище многоклеточных форм может размножаться путем фрагментации на отдельные участки, способные к дальнейшему развитию. К вегетативному размножению относится также почкование, имеющее место у видов рода Palatinella Laut.
Бесполое размножение с помощью амебоидов происходит, например, у представителей порядка Rhizochrysidales, особенно имеющих домики. Амебоиды очень мелкие, с короткими тупыми псевдоподиями, с одним хлоропластом или без него. Образуются они по 4 и более (Chrysocrinus, Stephanoporos), выползают из отверстия домика и образуют затем свой собственный. Реже амебоиды возникают в значительном количестве в специальных образованиях, напоминающих зооспорангий (Myxophrysis paradoxa).
Зооспоры могут возникать в обычных вегетативных клетках или в особых органах - зооспорангиях (Apistonema commutatum Pasch.). Обычно образуется по 2 зооспоры, реже по 4-8 (Phaeothamnion confervicola Lager h.) и только у отдельных видов число зооспор может составлять 16 (Моnochrysis aphanaster Skuja). Нередко в клетке возникает лишь одна зооспора (Celloniella palensis Pasch.). У большинства видов зооспоры яйцевидные, реже шаровидные (Chrysostephanosphaera globulifera Scherff.), тетраэдрические (Hydrurus foetidus (Vill.) Kirchn.) и др. В поперечном сечении зооспоры округлые, у некоторых видов несколько сжатые (Chrysotylos ferrea). Преобладают одножгутиковые - хромулиноподобные зооспоры, реже встречаются двужгутиковые с неравными жгутиками - охромонасоподобные (Phaeothamnion borzianum Pasch.).
У некоторых хризокапсовых и хризосферовых кроме зооспор образуются автоспоры. Развиваются они непосредственно в вегетативных клетках по 4 и 7, реже по 16 (Chrysapion rhigophylos Pasch. et Vlk), и по форме напоминают материнские клетки.
Половой процесс редок, представлен в виде гологамии, изогамии и автогамии. При гологамии две одинаковые исходные с вегетативными клетки сливаются в двуядерные зиготы, превращающиеся в кремнистые цисты, которые после периода покоя прорастают в одну или несколько клеток (Dinobryon borgei Lemm.). При изогамии сливаются гаметы с образованием зиготы с диплоидным ядром, которое редукционно делится; из зиготы образуются 4 гаплоидные особи (Ochrosphaera neapolitana). Автогамный половой процесс отличается от предыдущих тем, что слияния протопластов не происходит. Разрастающиеся двуядерные клетки превращаются в кремнистые цисты, как и при гологамии (Stenokalyx, Kephyrion Pasch. и другие роды). Нередко у одного и того же вида одновременно образуются цисты и происходит гологамный и автогамный половой процесс (Dinobryon borgei, Stenokalyx inconstans Schmid. и др.). Вероятно, половой процесс у золотистых водорослей не такое уж редкое явление [513], однако изучен он еще недостаточно.
Характерной особенностью цикла развития золотистых является образование цист. Они бывают эллипсоидной, яйцевидной, цилиндрической или иной формы, с целлюлозной оболочкой, позже пропитывающейся оксидом кремния. Стенки цист обычно толстые, бесцветные или окрашенные в бурый цвет, гладкие или покрытые различными скульптурными образованиями в виде бородавочек, шипов, реже морщин (Chromulina echinocystis Conr.), крыловидных образований (Celloniella palensis Pasch.) или экваториальных ободков (Hydrurus foetidus). Особенно разнообразны шипы, короткие или длинные, заостренные или тупые и раздвоенные (Ochromonas fragilis). Скульптурные образования размещены преимущественно равномерно на поверхности цист, однако у некоторых видов они локализуются в задней части (Uroglena soniaca Conr., Ochromonas eehinulata). У большинства видов диаметр цист не превышает 20 мкм и только в пределах таких родов, как Mallomonas или Microglena, цисты больших размеров - 34 мкм в длину и 23 мкм в ширину (Mallomonas allorgei (Dofl.) Conr., M. clavata Conr.). Реже размеры цист гораздо меньше - до 3 мкм (Chromulina minima Dofl.).
Цисты снабжены отверстием или порой, которая иногда вытянута в воротничок различной формы, шейку (Uroglena volvox Ehr.), крыловидный вырост (Uroglena soniaca) и т. д. Циста сверху закрыта пробочкой различной формы. Образуются цисты в разное время года, обычно летом и осенью (май, октябрь). Прорастание цист изучалось недостаточно и на нескольких видах. Известно, что при прорастании цист могут возникать от 1 до 4 и даже до 8 клеток. Цисты бывают одноядерные бесполые, двух ядерные бесполые (потенциально аутогамные), двухядерные половые (зиготические). Все цисты морфологически идентичны. Отдельные клоны в пределах видов способны к образованию цист всех типов или только одного.
Издавна золотистые водоросли родственно связывали с желтозелеными и диатомовыми. Так, А. Пашер [576, 579] объединил эти группы водорослей в одну филу под названием Chrysophyta. Особое значение он придавал образованию у представителей этих групп эндогенных цист, оболочки которых пропитаны кремнеземом, а также тому, что оболочки цист желтозеленых состоят из двух частей (створок), а цисты золотистых закрыты пробочкой, которую А. Пашер рассматривает как гомолог второй створки. Кроме того, золотистые, желтозеленые и диатомовые не образуют крахмала и для всех этих групп характерно отсутствие хлорофилла с. Все это говорит в пользу предположения А. Пашера о родстве золотистых, желтозеленых и диатомовых водорослей. Однако, как отмечал Д. К. Зеров [120], нельзя не отметить, что у этих групп имеются и существенные различия. Так, у золотистых обнаружены ксантофиллы, которых нет у представителей двух других групп, и только у желтозеленых обнаружен хлорофилл е. Диатомовые резко отличаются от золотистых и желтозеленых циклом развития и своеобразным строением ядра. Несмотря на черты сходства между золотистыми, желтозелеными и диатомовыми водорослями, А. Пашер все же отмечал, что они представляют собой самостоятельные стволы развития, обнаруживающие больше родственных связей друг с другом, чем с другими отделами [576]. Поэтому Д. К. Зеров, не отрицая общности происхождения по крайней мере золотистых и желтозеленых водорослей, рассматривал их как самостоятельные филы, а в систематическом отношении - как самостоятельные отделы автотрофных эукариотических организмов. Этой точки зрения придерживается большинство исследователей.
Что касается классификации золотистых, то, например, А. Пашер делит отдел на 5 классов: Chrysomonadineae с 3 подклассами - Chrysomonadeae, SUicoflagellatae, Coccolithophoreae, Rhizochrysidineae, Chrysocapsineae, Chrysosphaerineae, Chrysotrichineae. Систему А. Пашера с незначительными и несущественными изменениями принимают многие современные авторы. Основные таксоны золотистых, как и других отделов водорослей, А. Пашер выделил на основании особенностей их морфологии. Исходной группой он считал Chrysomonadineae, из которой выводил амебоидные Rhizochrysidineae, пальмеллоидные Chrysocapsineae, коккоидные - Chrysosphaerineae и нитчатые Chrysotrichineae. А. М. Матвиенко [203, 204], принимая классификацию А. Пашера, в основании системы золотистых ставит Rhizochrysidineae (Chrysopodinophvceae). Были и другие предложения, например, выделять золотистые с двумя бичевидными жгутиками и часто с гаптонемой между ними в особый класс [573], который Христенсен [375] назвал Haptophyceae. П. Буррелли [358] дал несколько иную классификацию золотистых водорослей, также основанную на строении жгутиков. По Д. К. Зерову [120], отдел Chrysophyta делится на 2 класса: Heterochrysophyceae и Isochrysophyceae. В пределах класса Heterochrysophycеае выделяется 7 порядков: Chrysomona dales, Coccol it hales, Dictyochales (Silicoflagellatae), Rhizochrvsidales, Chrysocapsales, Chrysosphaerales, Phaeothamniales (Chrysotrichales). Выделение их основано на различиях в типе структуры, числе и строении жгутиков, на способности образовывать кок колиты, особенностях строения скелета и панциря, на наличии или отсутствии слизистой обвертки и т. д. В основу разделения порядков на семейства положены такие признаки: число жгутиков, наличие или отсутствие псевдоподий или аксоподий, тип структуры и др. В пределах класса Isochrysophyceae различают два порядка: Isochrysidales и Prymnesiales.
Д. К. Зеров предполагает, что среди золотистых водорослей намечается две филогенетические ветви: изохризофитная и гетерохризофитная [120]. В гетерохризофитной ветви основной исходной группой следует признать хризомонадовые (Chrysomonadales). От них, как боковые ветви, могли развиваться, с одной стороны Coccolithales и Dictyochales (Silicophlagellatae), а с другой - амебоидные Rhyzochrysidales, пальмеллоидные Chrysocapsales и коккоидные Chrysosphaerales, которые в свою очередь могли дать начало нитчатым и пластинчатым Phaeothamniales. Классы Isochrysophyceae и Heterochrysophyceae, очевидно, обособились на ранних стадиях становления отдела Chrysophyta.
Золотистые водоросли распространены по всему Земному шару, в водоемах разных типов. Преимущественное большинство хризофитовых обитает в пресных водах и только незначительное число их обитает в море. Пресноводные виды встречаются в стоячих водоемах: болотах, озерах, прудах, старицах рек, водохранилищах, реже - в реках и в водоемах с текущей водой. Типичным местообитанием золотистых являются сфаговые болота с кислой реакцией воды. Известны виды, обитающие в водоемах, богатых известью (Heliaktis conradiana Pasch.), а также при pH воды 8,2 (Chrysapion rhigophylos, Koinopodium aggregatum Pasch. и др.). Золотистые водоросли обычно обитают в чистой воде, немногие из них встречаются в водоемах, загрязненных различными органическими веществами, среди гнилых листьев и водорослей, в загрязненных водоплавающей птицей прудах (Synochromonas pallida Korsch., Pseudokephyrion spirale Schmid.
Экология золотистых водорослей довольно хорошо изучена. Преимущественно это холодолюбивые организмы, встречающиеся в холодное время года, ранней весной, поздней осенью, а также зимой, подо льдом. Исключение составляют виды, обитающие в теплых, сильно прогревающихся водоемах (Eleutheropyxis oblonga Pasch.). Некоторые виды вызывают "цветение" воды, придавая ей желтовато-коричневую окраску (виды родов Mallomonas, Chromulina, Synura, Dinobryon, Uroglena, Uroglenopsis Lemm.).
Золотистые водоросли встречаются в планктоне, бентосе и нейстоне. Преимущественное большинство из них типичные представители планктона. К типичным бентосным формам относятся немногие виды родов Phaeodermatium Hansg., Celloniella, Hydrurus. Как правило это прикрепленные формы, живущие в холодных текущих чистых водах - на дне рек, каменистой почве каналов, деревянных сваях и камнях. Здесь встречаются и ползающие по дну с помощью ризоподий виды родов Chrysamoeba, Chrysarachnion, Heliapsis Pasch. Среди золотистых есть и эпифиты, обитающие на различных водорослях, а также на других золотистых водорослях (Lagynion scherfelii Pasch., Lepochromulina calyx Scherff.) или на цистах динофитовых (Еpicystis peridinearum Pasch.). Наиболее типичным представителем нейстона является Chromulina rosanoffii (Woronin) Biitschli, весь цикл развития которой проходит в поверхностной пленке воды стоячих водоемов. Нередко этот вид встречается в массовом количестве, образуя пленку почти монокультуры [204].
Золотистые водоросли принимают активное участие в накоплении органических веществ, ила (сапропеля), который имеет большое практическое значение для народного хозяйства. Определенное значение они имеют в цепи питания рыб. Как обитатели чистых вод, некоторые виды служат индикаторами на отсутствие загрязнения воды органическими веществами, т. е. могут быть использованы при санитарно-биологическом анализе воды. При массовом развитии, приводящем к "цветению" воды, некоторые виды золотистых водорослей играют отрицательную роль в деле водоснабжения (вода становится непригодной для употребления, приобретает неприятный привкус и запах). Ископаемые остатки золотистых водорослей представлены главным образом кокколитофоридовыми и кремнистыми жгутиковыми, они играют определенную роль в образовании залежей извести и мела, в формировании осадочных пород [204].
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'