Эукариотические водоросли
10. Отдел 1. Эвгленовые водоросли - Euglenophyta (З. И. Ветрова)
Эвгленовые водоросли представляют собой высокоорганизованную своеобразную группу низших растений, стоящую на границе растительного и животного мира и поэтому названную еще в XIX ст. порубежниками. Отдел объединяет свыше 900 видов микроскопических одноклеточных, преимущественно жгутиковых, эукаритических водорослей зеленого, редко красного цвета. Имеется много бесцветных форм. Представители этого отдела характеризуются следующими признаками.
1. Одноклеточные организмы с монадным типом структуры, имеющие одни или несколько жгутиков, за исключением небольшой группы безжгутиковых эвгленовых, а также форм, ведущих преимущественно неподвижный образ жизни и образующих колонии.
2. Клетки лишены целлюлозных оболочек. Под плазмалеммой расположен плотный, эластичный, белковой природы слой протопласта, называемый пелликулой, от плотности которой зависит постоянство формы клеток. Виды с тонкой и гибкой пелликулой не имеют постоянной формы, им свойственна метаболия, т. е. изменчивость формы тела. Под пелликулой у многих эвгленовых в наружных слоях протоплазмы имеются слизеносные тельца. У представителей родов Trachelomonas Ehr., Strombomonas Defl. кроме пелликулы, вокруг клетки образуется твердый минеральный домик, состоящий из солей железа и марганца.
3. На переднем конце клетки имеется мешковидное углубление называемое глоткой, в расширенную часть которой - резервуар - изливается содержимое сократительных вакуолей.
4. Форма клеток довольно разнообразна, но наиболее распространенной считается веретеновидная или, как ее часто называют, эвгленообразная.
5. Основным запасным веществом является особый тип полисахарида - парамилон - который концентрируется вокруг пиреноида или образуется непосредственно в протоплазме.
6. Стигма, или глазок, расположена на переднем конце клетки вблизи жгутикового аппарата, образуется вне хлоропласта и состоит из разных по величине глобул, расположенных рядами по 3-5 в каждом; глобулы обособлены друг от друга тонкой мембраной.
7. Ядро крупное, хромосомного типа, окружено двойной мембраной с порами. Нуклеоплазма содержит хроматин и одно или несколько ядрышек. Они фельген-отрицательны и содержат РНК.
8. Митоз особого типа - эвгленомитоз.
9. Каждый хлоропласт имеет трехслойную мембрану, ламеллярная система группируется в пачки, по 3 плотно прилегающих диска в каждой [440, 528].
10. Типы питания: автотрофное, мезотрофное, гетеротрофное, что обусловливает наличие значительного числа бесцветных форм.
11. Размножение путем продольного деления. Цитокинез осуществляется перетяжкой при участии плазмаллемы и относится к самому простому типу.
12. Преимущественно пресноводные формы, довольно широко распространенные особенно в водоемах замедленного стока
Индивиды эвгленовых водорослей имеют разнообразную форму тела: шаровидную, эллипсоидную, яйцевидную, веретеновидную, ланцетовидную, игловидную, цилиндрическую, трапециевидную, ромбическую, фляжковидную, мешковидную и т. д. Многие клетки скручены или серповидно изогнутые, иногда с разными выростами (килями, гребнями, ребрами). На поперечном срезе клетки округлые, четырехугольные, 3-4-6-килевые, нередко в той или иной степени уплощенные. Для большинства клеток характерна билатеральная симметрия. Жгутиковое отверстие находится или точно посередине переднего конца или сдвинуто в сторону от продольной оси. Поэтому та сторона, на которой находится жгутиковое отверстие, называется брюшной, противоположная ей - спинной.
Размеры клеток сильно варьируют у видов разных родов. Среди бесцветных эвгленовых наиболее крупными размерами отличаются виды родов Cyclidiopsis Korsch., Peranema Duj., Heteronema Duj. em. Stein, наиболее мелкие клетки наблюдаются у родов Petalomonas Stein, Notosolenus Stokes em. Skuja, Scytomonas Stein, Entosiphon Stein, Urceolus Mereschk., Rhabdomonas Fres., Astasia Ehr. em. Duj. [15, 16, 39, 258, 259, 277].
Пелликула обычно штриховатая, толщиной до.70 нм. Штриховка бывает самая разнообразная - от мелких точек до ребер (рис. 10.1). Штрихи вместе с пелликулой выполняют защитную и опорную функции. Наклон штрихов отражает направление и степень скрученности клеток. Этот признак некоторые авторы используют в качестве таксономического.
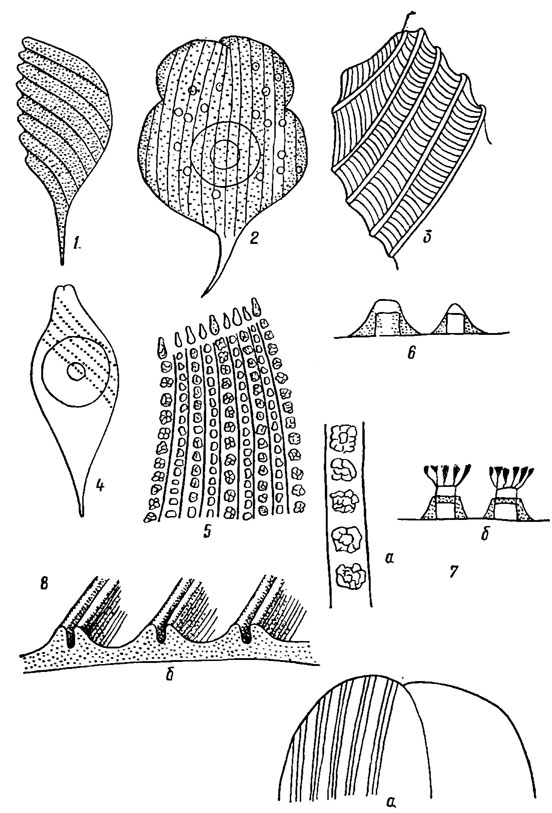
Рис. 10.1. Структура пелликулы эвгленовых водорослей: 1 - спиральные штрихи; 2 - продольные штрихи; 3 - ребра с поперечными полосами между ними; 4 - ряды бородавочек; 5-7 - скульптура Euglena spirogyra Ehr. var. fusca Klebs (6 - простые бугорки, продольный срез; 7 - сложные бугорки, вид сверху (а) и сбоку (б)); 8 - строение штрихов Phacus curvicauda Swir. (а - вид сверху, б - в проекции)
Под пелликулой у многих видов в наружных слоях протоплазмы располагаются слизеносные тельца - мелкие, наполненные слизью вакуоли, открывающиеся наружу порами в области штрихов. По форме они шаровидные, веретеновидные, иногда палочковидные, легко обнаруживаются с помощью прижизненной окраски (нейтральным красным, крезил-блау).
У некоторых бесцветных форм эвгленовых в субпелликулярном слое обнаружены трихоцисты (своеобразные стрекательные структуры). Они бывают трубчатые, веретеновидные или напоминающие по форме песочные часы. Сведений о механизме выстреливания трихоцист в литературе нет.
Образование минеральных домиков, характерных для родов Trachelomonas, Strombomonas и Ascoglena Stein, связано с размножением монад. Их формирование начинается вскоре после выхода монады из отверстия старого домика. Выскользнувшая монада, вначале вытянутая в длину, некоторое время свободно плавает, затем округляется и выделяет вокруг себя нежную бесцветную оболочку, которая постепенно утолщается, буреет и приобретает соответствующую орнаментацию. Домики бывают самой разнообразной формы. Как правило, они имеют радиальную симметрию, на поперечном срезе округлые, иногда сжатые. Жгутиковое отверстие представляет собой крупную пору, снабженную валикообразным утолщением или выростами в форме зубцов или сплошной трубки, называемой горлышком, или воротничком (рис. 10.2). Наличие или отсутствие воротничка, его форма, величина, изогнутость имеет таксономическое значение. Домики редко бывают гладкие. Обычно они в той или иной мере орнаментированы.
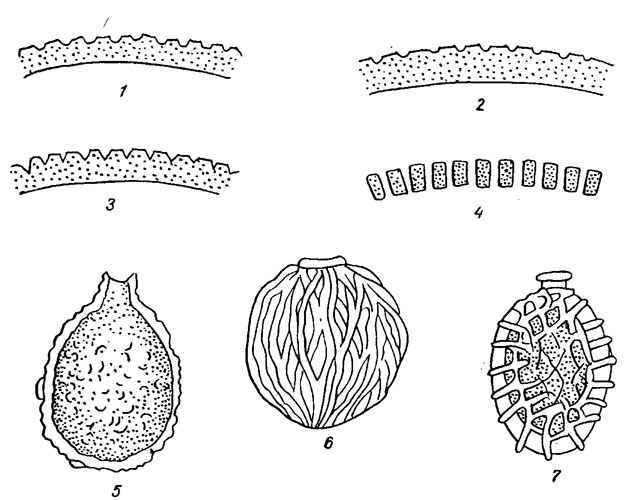
Рис. 10.2. Структура домиков эвгленовых водорослей: 1-4 - стенки домиков в поперечном разрезе (1, 2 - с точечными утолщениями и мелкими углублениями, 3 - со скробикулированной поверхностью, 4 - со сквозными порами); 5-7 - поверхность домиков (5 - морщинистая; 6 - полосатая, 7 - сетчатая)
Количество и длина жгутиков у представителей разных родов эвгленовых сильно варьируют. Виды рода Eutreptia имеют 2 почти одинаковых по длине жгутика, родов Eutreptiella Da Cunha и Distigma Ehr. - 2 неравных жгутика, из которых один длинный (основной) и второй короткий (дополнительный). У родов Euglena, Phacus, Trachelomonas, Lepocinclis и других короткий жгутик более тонкий [525], не выходит из глотки и соединяется с длинным жгутиком на уровне стигмы. Наружная часть длинного жгутика часто сбрасывается. Жгутик покрыт очень тонкими и длинными волосками - мастигонемой - разной длины (рис. 10.3). Есть виды вообще не образующие жгутик (безжгутиковые эвглены). Подробно строение жгутиков и других органелл клетки изложено в разделе 1.2,2. Следует указать, что корешковая система жгутиков эвгленовых водорослей отличается однородностью, поскольку представлена тремя микротрубочновыми корешками, отходящими от двух параллельно расположенных кинетосом. Они выстилают стенки жгутикового резервуара и направлены в переднюю часть клетки. Корешки, идущие в глубь клетки или к ядру, у эвгленовых не обнаружены.
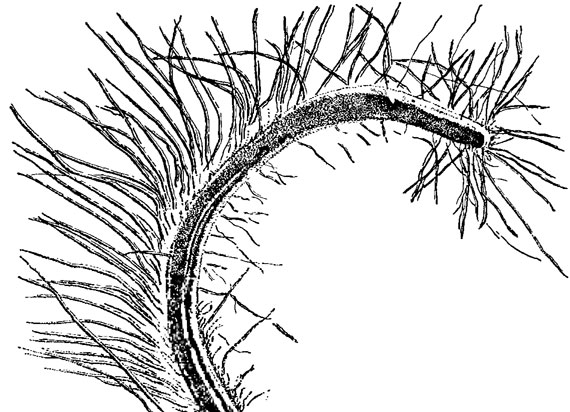
Рис. 10.3. Дистальный конец жгутика Euglena spirogyra Ehr. с мастигонемой
Особенностью митоза является то, что при делении ядра ядрышко делится как отдельная структура, ядерная оболочка при этом сохраняется. Ядрышко хорошо красится гематоксилином, но не дает реакции по Фельгену.
Аппарат Гольджи у эвгленовых состоит из плоских цистерн, плотно прилегающих друг к другу, расположенных группами. На одном или обоих концах цистерн имеются отростки в виде пузырьков (рис. 10.4) [439, 593, 605]. Обычно он расположен в непосредственной близости от глотки. Его функции связаны с секреторной деятельностью клетки.
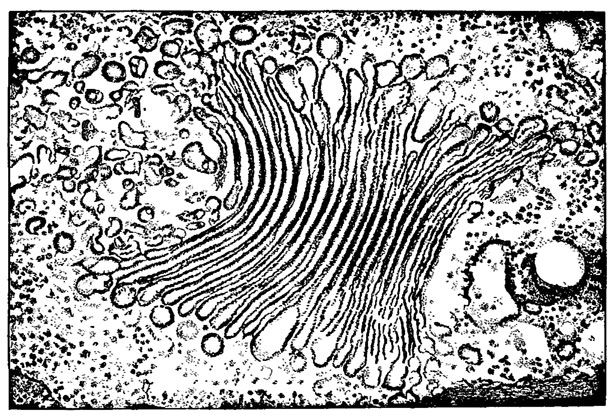
Рис. 10.4. Аппарат Гольджи Euglena spirogyra Ehr.
Митохондрии сравнительно крупные, часто овальной формы, размеры их колеблются в широком диапазоне (рис. 10.5). Их число варьирует в зависимости от возраста клетки и ее функционального состояния. Располагаются они обычно по периферии клетки. Эти органеллы окружены двойной мембраной: наружная гладкая, сплошная, внутренняя образует выросты или кристы, обращенные в гомогенный матрикс. Кристы обычно пластинчатые и расположены перпендикулярно продольной оси митохондрий.
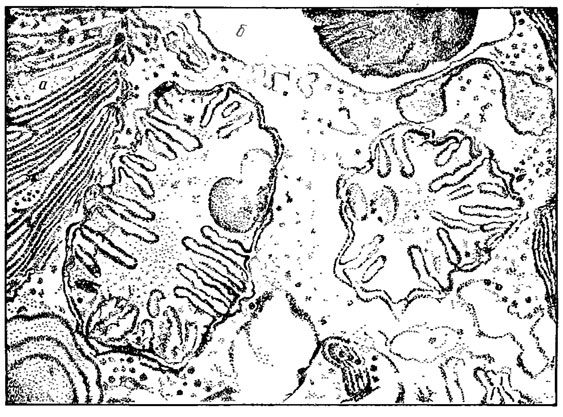
Рис. 10.5. Митохондрии Euglena spirogyra Ehr. в поперечном разрезе: а - хлоропласт, б - фосфолипидный пузырек
Эндоплазматический ретикулюм представляет собой систему канальцев, которые местами расширяются, образуя цистерны или пузырьки, окруженные мембранами (гладкой и шероховатой), несущей рибосомы. Он принимает участие в перемещении и распределении различных веществ в клетке.
Микротрубочки представляют собой очень динамичные клеточные структуры, то появляющиеся, то исчезающие, перемещающиеся, увеличивающиеся или уменьшающиеся в числе. Чаще всего они сосредоточены на периферии клетки, вокруг стигмы [688], в области глотки, у основания жгутиков [524 , 609, 640].
Зеленая окраска хлоропласта обусловлена присутствием хлорофиллов a и b. Снаружи хлоропласт окружен каналом эндоплазматического ретикулюма. По форме хлоропласты эвгленовых довольно разнообразны (рис. 10.6): дисковидные, чечевицеобразные, пластинчатые, лентовидные, звездчатые, лопастно-рассеченные [15, 16,39]. Число их колеблется от 1-2 до многих. Некоторые виды рода Euglena (Е. sanguinea Ehr., Е. haematodes (Ehr.) Lemm.) вырабатывают большое количество внепластидного пигмента красного цвета, не принимающего участия в фотосинтезе и являющегося, видимо, лишь ширмой, изменяющей интенсивность и качество света, падающего на хлоропласты.
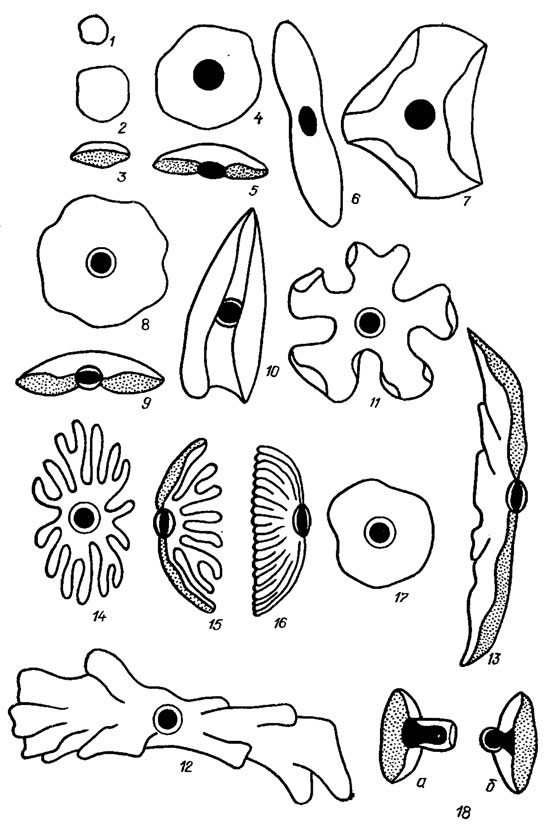
Рис. 10.6. Хлоропласты эвгленовых водорослей: 1, 2 - дисковидные; 3 - чечевицеобразные; 4-7 - с голыми пиреноидами; 8-10 - с двускорлупчатыми пиреноидами; 11-16 - лопастно-рассеченные с двускорлупчатыми пиреноидами; 17, 18 - с внутренними пиреноидами (17 - вид сверху, 18 - чечевицеобразный пиреноид с цилиндрическим (а) и сферическим (б) выступами)
В хлоропластах у большинства эвгленовых обнаружены более плотные образования - пиреноиды, в которых помимо ферментов обнаружены следы ДНК, РНК и даже хлорофилла. Пиреноиды обычно окружены сильно преломляющей свет обкладкой, в образовании которой принимает участие парамилон. Он окружает пиреноид в виде двух скорлупок, отчего такие пиреноиды получили название двускорлупчатых. Иногда встречаются односкорлупчатые пиреноиды. Изредка пиреноиды не имеют обкладки и в таком случае называются голыми. Имеются еще внутренние пиреноиды (см. рис. 10.6). Они сильно выступают со стороны хлоропласта, обращенной внутрь клетки, а парамилон на них откладывается в виде узких колпачков или наперстков.
Полисахарид парамилон откладывается в клетке в виде зерен различной формы и величины. Он состоит из остатков глюкозы, соединенных β-1,3-связью [377, 440, 442, 515]. При гидролизе парамилона образуется глюкоза. С иодом он не дает цветной реакции. Зерна его разбухают при действии слабого раствора гидроксида калия и других щелочей.
Парамилон образуется на наружной стороне пиреноида в виде вогнутых скорлупок. Если хлоропласты лишены пиреноидов, парамилон откладывается в протоплазме. Непосредственно в протоплазме образуется парамилон и у бесцветных форм. Число зерен парамилона и их размеры в клетках эвгленовых может быть разным. Они бывают мелкими и многочисленными, или крупными от одного до нескольких (крупные зерна парамилона М. Шадефо [370] назвал парамилиями). Мелкие зерна разбросаны в протоплазме беспорядочно, положение парамилий строго локализовано. У одних видов они лежат посередине клетки, вдоль ее продольной оси, у других расположены по периферии. Считают, что парамилон, кроме энергетической функции, имеет определенное отношение к передвижению клеток.
Стигма присутствует у всех зеленых и отчасти у бесцветных эвгленовых. Она представляет собой густое скопление оранжево-красных липидных капелек, расположенных в одной плоскости [439, 701]. У эвгленовых стигма расположена на переднем конце клетки, вне хлоропласта, прилегая к спинной стороне глотки, обычно в месте перехода ее в глоточный канал (рис. 10.7). Ультраструктура стигмы и ее функции описаны во многих публикациях [397, 398, 506, 525, 688 , 690, 692], а также в разделах 1.2.2 и 1.3.1. В состав стигмы входит да нескольких десятков разных по размеру пигментных глобул, которые располагаются рядами по 3-5 в каждом и обособляются друг от друга тонкой мембраной (см. рис. 10.7). Стигма такой организации свойственна только эвгленовым водорослям. В стигме обнаружены флавины, липиды, каротиноиды, β-каротин, эхиненон, криптоксантин и др. [344, 457, 479, 568, 569, 689].
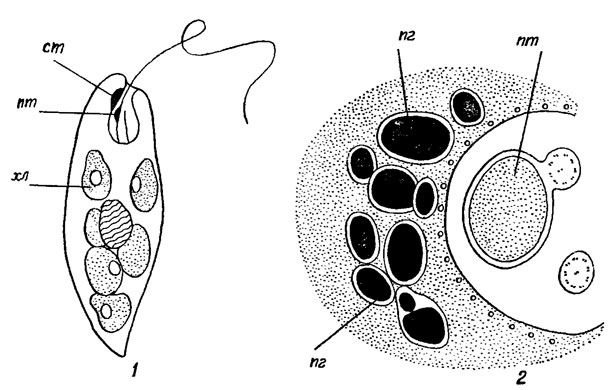
Рис. 10.7. Расположение стигмы в клетке эвгленовой водоросли (1) и ее строение (2): ст - стигма; пт - парабазальное тело; хл - хлоропласт; пг - пигментные глобулы
Фоторецептор, или парафлагеллярное тело [489] - особая светочувствительная органелла, напоминающая висячую слезу или пластинку с выростами и расположенная на жгутике на уровне стигмы (рис. 10.8). При сбрасывании наружной части жгутика фоторецептор остается. Обычно он наблюдается у тех эвгленид, которые имеют стигму. По своим функциям фоторецептор тесно связан со стигмой и жгутиками и играет определенную роль в движении эвгленовых.
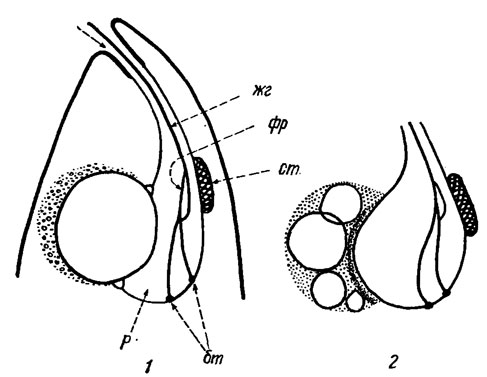
Рис. 10.8. Строение глотки эвгленовых водорослей: 1 - вакуоль перед опорожнением в полость резервуара (жг - жгутик, фр - фоторецептор, ст - стигма, бт - базальные тела, р - резервуар); 2 - вакуоли перед слиянием
На переднем конце клетки находится постоянная полость - глотка, с выводным отверстием наружу (см. рис. 10.8). Ее расширенная часть - резервуар, или главная (сборная) вакуоль, переходит в узкий выводной канал - глоточный канал. Та часть протоплазмы, которая прилегает к брюшной стороне резервуара, отличается зернистостью и способностью образовывать вакуоли. Первоначально возникают мелкие многочисленные вакуоли, которые затем последовательно сливаются в одну (иногда в две и больше) вакуоль, содержимое которой поступает в полость резервуара. В старых клетках нередко наблюдается одна крупная центральная вакуоль.
Размножение эвгленовых происходит путем деления надвое в подвижном или пальмеллевидном состоянии. Перед делением клетки обычно становятся неподвижными, сбрасывая наружную часть жгутика, и часто выделяют вокруг себя слой слизи. Разделению клетки предшествует деление ядра, образование нового жгутикового аппарата, деление хлоропластов, пиреноидов, стигмы. Деление подвижных особей происходит всегда продольно от переднего конца клетки, за исключением представителей рода Colacium Ehr., ведущих прикрепленный образ жизни, у которых этот процесс начинается со свободного морфологически заднего конца тела. У некоторых видов рода Trachelomonas плоскость деления монады часто не совпадает с продольной осью домика, а у видов рода Phacus деление клеток происходит в плоскости, параллельной сжатию. Поведение ядра при делении клеток у многих видов изучал Г. Лидейл [522, 526].
Имеются указания о наличии полового процесса у эвгленовых. Описан гологамный половой процесс у одного из представителей рода Scytomonas (= Copromonas) - S. subtil is Dobell [390] и у Euglena sp. Аутогамный половой процесс и последующий мейоз описан у Phacus pyrum (Ehr.) Stein [516, 523]. Однако эти сведения явно недостаточны для того, чтобы считать данные, приведенные в упомянутых выше работах вполне достоверными. Пока что можно утверждать, что половой процесс, известный у эвгленовых, не является характерной чертой их цикла развития.
Цикл развития у эвгленовых водорослей имеет две основные формы. Большинство представителей обычно вегетирует в двух состояниях - подвижном и временно неподвижном, пальмеллевидном. В состоянии пальмеллы клетки округлой формы лишены жгутиков, но стигма при этом сохраняется. Покоящиеся стадии у таких эвгленовых представлены округлыми клетками с толстой, иногда слоистой оболочкой, из которой при благоприятных условиях возникает одна вегетативная клетка. У других эвгленовых, особенно у всех неметаболичных видов имеется лишь подвижная стадия. В состоянии покоя они сохраняют форму вегетативных клеток, хотя часто сбрасывают жгутик, утолщают оболочки и переполняются парамилоном.
В цикле развития некоторых представителей рода Colacium наблюдается сезонная смена морфологических форм развития - цикломорфоз. Так, С. libellae Ros. et Wil. в весенне-осенний период представлен свободно живущими одноклеточными жгутиковыми или колониями из нескольких клеток, сидящих на стебельке. С наступлением зимы клетки С. libellae проникают в прямую кишку личинок некоторых стрекоз и существуют там в пальмеллевидном состоянии в виде скоплений неподвижных клеток, окруженных слоем слизи. Весной когда личинки поднимаются в верхние теплые слои воды, клетки С. libel lae выходят наружу и начинают самостоятельно вегетировать.
Происхождение эвгленовых и их филогенетические связи трактуются по-разному. Еще со времен Ф. Штейна высказывалось предположение о возможности общего происхождения эвгленовых и зеленых водорослей. В качестве основного доказательства родства этих групп организмов принимали наличие пигментации (хлорофиллов a и b). На этом основании эти две группы водорослей связывают и даже объединяют в один отдел зеленых водорослей [32, 375, 509].
Некоторые ученые, основываясь на таких признаках, как отсутствие полового процесса, наличие большого числа гетеротрофных форм, отложение запасных продуктов вне пластид и наличие глотки, относят эвгленовые водоросли вместе с Cryptophycineae, Dinophycineae и Raphidophycineae к отделу Chromophycophyta [370, 371]. Существуют представления о родственных связях эвгленовых с рафидофитовыми [65, 257, 529, 626]. Однако эта точка зрения имеет много противников (А. Пашер, Ф. Фрич, Г. Прескот, А. Холланд, X. Скуя, Б. Фотт, Я. Миньо, Д. К. Зеров).
По нашему мнению наличие ряда специфических признаков у эвгленовых не позволяет тесно связывать или объединять их с другими отделами водорослей. Так, от зеленых водорослей эвгленовые резко отличаются протеинсодержащей пелликулой, наличием глотки, слизистых телец, внепластидного запасного продукта парамилона, составом каротиноидов, особенностями строения и митотического деления ядра, уникальным строением стигмы, особой структурой хлоропласта. С криптофитовыми водорослями эвгленовые действительно близки по некоторым признакам, но и резко отличаются от них составом пигментов и химической природой запасных продуктов, строением стигмы, хлоропластов, организацией клеточного покрова [39]. О нецелесообразности объединения эвгленовых с дин офитовыми свидетельствуют различия в пигментном составе, отличия в строении ядерного аппарата, а также наличие у динофитовых особого типа вакуолей (пузул), стигмы, клеточного покрова в виде теки, сложно организованных стрекательных структур.
Что касается объединения эвгленовых с рафидофитовыми, то правильнее было бы рассматривать рафидофитовые как небольшую самостоятельную филу [8, 17, 120, 595]. Сравнительное изучение ультраструктуры клеток эвгленовых и рафидофитовых убедительно показало, что рафидофитовые имеют ряд своеобразных признаков, не позволяющих объединять их в один отдел с эвгленовыми [553]. Это в первую очередь касается строения ядерного и вакуолярного аппаратов, структуры клеточного покрова, состава пигментов и продуктов ассимиляции.
Современные эвгленовые, хотя и представлены одноклеточными формами, являются довольно высокоорганизованными и сложными организмами по сравнению с теми древними примитивными формами, от которых они ведут свое происхождение. Предполагают, что гипотетическими предками эвгленовых были сильно метаболичные эвгленообразные гетеротрофы, эволюция которых шла в двух основных направлениях [12, 15]. Одно из них вследствие дальнейшего развития и усовершенствования организации клеток в результате симбиогенного приобретения хлоропластов привело к становлению современных представителей (фототрофных и сапрофитных) порядка Euglenales. Вторая линия развивалась по пути специализации примитивных гетеротрофов, связанному с реорганизацией воспринимающего аппарата, что привело к возникновению первичных голозойных форм с узкоспециализированным типом питания, из которых образовались высокоорганизованные современные представители порядка Peranematales.
В основу современной систематики эвгленовых положены следующие основные критерии: тип питания, число жгутиков, степень морфологической дифференциации тела, наличие или отсутствие палочковой органеллы, степень метаболии. На основании этих признаков в отделе Euglenophyta выделен один класс Euglenophyceae, содержащий три порядка Euglenales, Peranematales, Euglenomorp hales [8, 16, 39].
Роль в природе эвгленовых водорослей определяется их довольно широким распространением. Они встречаются в самых разнообразных местообитаниях - от небольших водоемов с различной степенью загрязнения до крупных рек и водохранилищ. Эвгленовые - обитатели преимущественно пресных вод. Большинство из них развивается в поверхностных или придонных слоях воды. Некоторые виды приспособились к ползанию по дну. Среди эвгленовых встречаются и такие, которые образуют на поверхности воды нейстонные пленки зеленого, желто-зеленого, бурого, кирпично-красного цвета [18]. Некоторые бесцветные формы обнаружены в почвах на глубине 8-25 см в довольно значительном количестве (почти 100 штук в 1 г почвы). Некоторые виды (род Colacium) ведут прикрепленный образ жизни на различных зоопланктерах. Среди эвгленовых обнаружены и паразитические формы, живущие в кишечниках олигохет, нематод, копепод, амфибий, на жабрах рыб. Эвгленовые нередко поражаются различными паразитами (Sphaerita, Pseudosphaerita, Polyphagus).
Эти водоросли интенсивно развиваются в водоемах замедленного стока с умеренной минерализацией (100-700 мг/л) и повышенным содержанием органических веществ и биогенных элементов [6, 9-11, 13, 14, 18, 38]. Они могут развиваться в условиях значительных колебаний температурного режима (0-27°С) и pH среды (3,0-8,9), однако максимум вегетации обычно наблюдается при температуре воды 20-27°С, pH 6,0-7,0. При уменьшении уровня воды в пересыхающих в летний период эфемерных водоемах обеднение видового состава эвгленовых происходит в первую очередь за счет бесцветных форм. Установлено, что значительное число фототрофных эвгленовых отличается широкой экологической амплитудой и повсеместным распространением.
Хозяйственное значение эвгленовых определяется способностью некоторых из них к миксотрофному питанию, что обеспечивает их участие в самоочищении пресных вод и в доочистке сточных вод различных предприятий. Изучение этих организмов интересно в теоретическом и практическом отношении. Их используют как удобный модельный объект при разработке вопросов физиологии, биохимии, генетики и цитологии клетки, а также при решении некоторых общебиологических проблем. Ряд видов используется как биологические индикаторы загрязнения водоемов бытовыми органическими веществами в системе экологического мониторинга. Отдельных представителей эвгленовых применяют в медицинской практике как модельные объекты для выяснения действия различных антибиотиков, гербицидов, гибберелинов, для количественного определения в среде витамина В12 и т. д. Довольно заметно участие эвгленовых водорослей в формировании пищевых звеньев в водоемах. В общем, в настоящее время эвгленовые изучаются как существенный компонент альгоценозов. Исследование химического состава эвгленовых показало довольно высокое содержание в них белков, витаминов и других биологически активных веществ, что небезынтересно с точки зрения их изучения и использования в практике народного хозяйства.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'