8.5. Обзор важнейших современных систем водорослей (Н. П. Масюк)
Для современной альгологической систематики характерно множество разнообразных систем. Даже на самых высоких таксономических уровнях царств (надцарств, подцарств), отделов и классов систематики не могут прийти к единому мнению (табл. 8.2, см. вклейку). Некоторые ученые вообще отказываются от таких таксономических категорий как надцарство, царство, подцарство [77, 356, 417, 426, 483, 484, 685] и отдел [426, 632, 633]. В последнем случае в качестве наивысшей таксономической категории принимается класс. От объединения классов водорослей в таксоны более высоких рангов отказались также редакторы известного издания "Suβwasserflora von Mitteleuropa" (1978-1985)*.
* (Исключение сделано лишь для классов зеленых водорослей, объединенных в отдел Chlorophyta.)
В современной альгологической систематике наблюдаются две противоположные тенденции: с одной стороны, тенденция к сокращению количества отделов и умножения количества классов, с другой, - к увеличению числа отделов и уменьшению классов. М. Шадефо [371] впервые выдвинул концепцию четырех отделов водорослей, относящихся к двум царствам (см. табл. 8.2). Эту концепцию принял Т. Христенсен [374, 376]. Однако системы этих двух авторов неидентичны, они различаются, как номенклатурно, так и по систематическому статусу отдельных таксонов, например, по положению Euglenophyceae: у М. Шадефо этот класс отнесен к отделу Chromophyta, у Т. Христенсена - к Chlorophyll. Концепцию четырех отделов (но без выделения царств) принимают также Б. Фотт [417] и Ф. Гиндак с соавторами [484]. Однако объем отделов Chromophyta и Chlorophyta у этих авторов не одинаков, так как Б. Фотт выделяет из состава первого отдела классы Cryptophyceae и Chloromonadophyceae, а из второго - Euglenophyceae, создавая искусственную группу "жгутиковых с неясным систематическим положением" [417, с. 413].
Тенденция к объединению многочисленных классов водорослей в немногие крупные группы еще ярче выражена в одной из более поздних работ М. Шадефо [372]. Автор рассматривает три большие группы водорослей, не придавая им статус отдела или таксона иного ранга: 1) архаическую цианородофициевую, объединяющую два класса прокариотических (Cyanoschizophyceae, Chloroschizophyceae) и два класса эукариотических водорослей (Glaucophyceae и Rhodophyceae), 2) хромофициевый цикл (Xanthophyceae, Raphidophyceae и Eustigmatophyceae; Chrysophyceae, Haptophyceae, Diatomophyceae и Phaeophyceae; Cryptophyceae и Dinophyceae; Euglenophyceae) и 3) хлорофициевую ветвь (Pocillophyceae, Prasinophyceae, Phragmophyceae, Zygophyceae и Euchlorophyceae).
С другой стороны, Ф. Гиндак и др. [483] рассматривают пять отделов: наряду с Cyanophyta, Rhodophyta, Chromophyta и Chlorophyta, в самостоятельный отдел выделены Euglenophyta. В системе Т. Калины [685] - восемь отделов: наряду с Cyanophyta, Rhodophyta, Chromophyta, Chlorophyta и Euglenophyta, в качестве самостоятельных отделов рассматриваются также Dinophyta, Cryptophyta и Chloromonadophyta. Х. Эттл [405] распределяет эукариотические водоросли между девятью отделами: кроме перечисленных выше, выделены также отделы Phaeophyta и Charophyta.
В системе Д. К. Зерова [120], в несколько модифицированном виде положенной в основу настоящего издания, 11 отделов, отнесенных к двум царствам - Procaryota и Eucaryota (см. табл. 8.2; рис. 8.1). В последнее время описан или выделен из состава других отделов ряд новых отделов: Prochlorophyta [533], Haptophyta, или Prymnesiophyta [469], Eustigmatophyta [584], Chlorarachniophyta [478], Zygnematophyta [54]. Некоторые из них признаны многими альгологами и приводятся в учебниках [54, 584].
Таким образом, для современной систематики водорослей характерно наличие множества систем, различающихся между собой в большей или меньшей степени даже на самых высоких таксономических уровнях. Еще больше разногласий существует на более низких уровнях. Нередко один и тот же таксон в одном и том же объеме рассматривается то в качестве отдела (Charophyta - М. Голлербах, Х. Эттл [79, 405]), то в качестве класса (Charophyceae - Д. Зеров, Б. Фотт [120, 417], то как порядок (Charales - Т. Христенсен [376]). Различный таксономический ранг придается диатомовым, желтозеленым, золотистым, рафидофитовым и многим другим систематическим группам водорослей. Иногда в работах одного и того же автора, опубликованных в разные годы, принимаются различные системы (сравни, например, работы Б. Фотта [414] и [417], Ф. Гиндака и др. [483] и [484]). Наблюдается переоценка систематических признаков и критериев. Все большее значение, особенно на уровне высших таксонов, приобретают биохимические и ультраструктурные признаки. Одновременно наблюдается снижение таксономического уровня значимости морфологического критерия. Продолжается интенсивный процесс описания новых таксонов и переведения некоторых ранее описанных в синонимы. Существенные изменения предлагают внести в классификацию зеленых, диатомовых, динофитовых и других групп водорослей [368, 612, 625 и др.]. Так, зеленые растения (Chlorophyta в широком понимании [см. 120]) предлагают выделить в самостоятельное царство Viridiplantae, в пределах которого рассматривают шесть отделов; четыре из них (Charophyta, Prasinophyta, Ulvophyta, Chlorophyta) объединяют водоросли, а два (Bryophyta и Tracheophyta) - высшие растения. Филогенетические системы, основанные на новых принципах, подчас кажутся парадоксальными или преждевременными, так как филогенетическая значимость используемых при их построении признаков нередко остается неясной (сравни [555]) или недостаточно большим оказывается объем проведенных исследований. Многообразие существующих систем водорослей допускает определенную степень свободы их выбора при проведении исследований несистематического профиля.
Причины нестабильности альгологической систематики кроются, прежде всего, в неразработанности общей теории систематики как науки [151], в частности, - в отсутствии общепринятого понимания сущности основных таксономических категорий. Известно, что "Международный кодекс ботанической номенклатуры" (см. раздел 8.4) утверждает лишь названия таксономических единиц и их последовательность в иерархической системе таксономических категорий, но не дает толкования их сущности, их оперативных определений. Обычно в систематических работах содержание основных таксономических категорий не обсуждается, а описание новых или ревизия уже существующих таксонов проводится без достаточно четкого определения позиций авторов относительно принципов выделения и сущности основных таксономических единиц. Однако без такого обсуждения работа систематика приобретает стихийный, интуитивный характер, а предлагаемые системы являются недостаточно обоснованными. Все еще недостаточно разработаны вопросы о специфике вида в альгологии как основной таксономической единицы, о принципах внутривидовой классификации, особенностях процессов макро- и микроэволюции.
Нестабильность современной альгологической систематики обусловлена также информационным взрывом, наблюдающимся в XX ст. в связи с внедрением новых методов исследования: электронно-микроскопических, биохимических, физиологических, генетических и других, со стремлением к использованию в филогенетических и систематических целях всей суммы информации о водорослях, полученной с помощью разнообразных методов исследования [301].
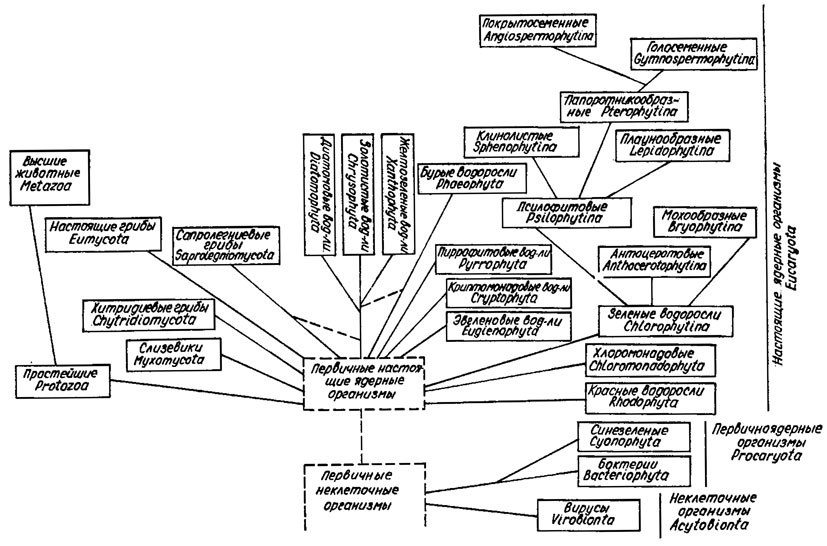
Рис. 8.1. Схема филогенетических связей между отделами (филами) и подотделами растительного мира (по Д. К. Зерову)
Одним из важнейших событий в эволюционной биологии XX ст. было открытие двух типов строения клетки: прокариотического и эукариотического, а также осознание глубоких, принципиальных различий между прокариотическими и эукариотическими организмами, проявляющихся не только в ультраструктурном, но и в генетическом, химическом, физиологическом и биологическом отношениях (сравни [91, 197]).
К числу важнейших цитологических признаков прокариот принадлежат отсутствие в их клетках окруженных мембранами ядер, пластид, митохондрий, аппарата Гольджи и других органелл, отсутствие тонопласта, эндоплазматической сети и микротрубочек, своеобразное строение клеточных покровов, более мелкие, чем у эукариот, размеры рибосом (70S). ДНК прокариот не имеет хромосомной организации, не связана с гистонами и обычно сосредоточена в нуклеоиде, не отделенном от цитоплазмы мембранами. Митоз и мейоз не наблюдаются. Цитоплазматическая ДНК находится в плазмидах и эписомах. Жгутики имеют субмикроскопические размеры и уникальное строение (наличие одной или нескольких микрофибрилл, не организованных в систему "9 + 2"). Дыхательная система сосредоточена в мембранах или мезосомах. Фотосинтетический аппарат представлен одиночными тилакоидами, расположенными непосредственно в цитоплазме, преимущественно в ее периферической области. В цитоплазме нередко наблюдаются окруженные мембранами газовые вакуоли; вакуоли с клеточным соком отсутствуют. К числу химических особенностей прокариот следует отнести отсутствие у них гистонов, своеобразный состав клеточных оболочек (наличие муреина), продуктов ассимиляции и комплексов фотосинтетических пигментов, отсутствие в мембранах стеролов. Из физиологических особенностей заслуживают внимания уникальные способности некоторых прокариот к облигатному анаэробиозу, к усвоению молекулярного азота, совмещению оксигенного и аноксигенного фотосинтеза, их исключительная устойчивость к экстремальным значениям различных факторов среды (длительному высушиванию, высоким температурам, высоким дозам ультрафиолетового и γ-излучений и пр.). Половой процесс фрагментарный (обычно передается лишь часть наследственной информации) (см. раздел 9).
Эукариотам свойственна компартментализация клетки на окруженные мембранами органеллы - ядро, митохондрии, аппарат Гольджи и другие, наличие эндоплазматической сети и микротрубочек, более крупные, чем у прокариот, размеры рибосом (805), хромосомная организация ДНК (молекула ДНК в хромосоме обычно связана с гистонами). Как правило наблюдается митоз и во многих случаях мейоз. Цитоплазматическая ДНК обнаружена в митохондриях, пластидах, по данным некоторых авторов, также в центр нолях, базальных телах, аппарате Гольджи. Жгутики микроскопических размеров построены из 20 микротрубочек по типу "9 + 2"*. Газовые вакуоли отсутствуют, в клетках нередко обнаруживаются вакуоли с клеточным соком или сократительные вакуоли. Цитоплазматические мембраны содержат стеролы. Дыхательная система сосредоточена в митохондриях, фотосинтетическая - в пластидах. Эукариоты неспособны к облигатному анаэробиозу, азотфиксации, аноксигенному фотосинтезу; верхние пределы устойчивости к экстремальным значениям факторов внешней среды ниже, чем у прокариот. При половом процессе передается весь набор хромосом**.
* (Отклонения от типа "9 + 2" редки, немногообразны и, по-видимому, являются его вариантами.)
** (Сравнительная характеристика прокариот и эукариот содержится в ряде работ [96, 227 и др.].)
Выявление этих кардинальных различий между прокариотами и эукариотами при полном отсутствии каких-либо промежуточных организмов выдвинуло проблему происхождения эукариотической клетки. Существующие разнообразные точки зрения можно свести к двум основным гипотезам: эндогенной, или автогенной, согласно которой эукариотическая клетка возникла путем постепенных преобразований, постепенного усложнения прокариотической [671], и экзогенной, или эндосимбиотической, согласно которой эукариотическая клетка с ее органеллами возникла путем симбиоза между разнородными микроорганизмами.
Выдвинутая в конце XIX - начале XX ст. А. Е. Шимпером (цит. по: [91]), К. С. Мережковским [218] и А. С. Фаминциным [312] гипотеза симбиогенеза, получившая поддержку в работах известных советских ботаников А. А. Еленкина и Б. М. Козо-Полянского, снова возродилась на основе достижений генетики, биохимии и молекулярной биологии. Наличие в клеточных органеллах эукариот собственной ДНК, отличающейся по размеру молекулы, нуклеотидному составу, способности к ренатурации и степени молекулярной гибридизации от ядер ной и более напоминающей по этим характеристикам ДНК прокариот, чем ДНК, заключенную в ядре той же клетки, отличия хлоропластных рибосом (по размеру и константе седиментации) от цитоплазматических рассматриваются как доказательства в пользу теории симбиогенеза.
Открытие разных типов тонкой структуры хлоропластов у красных, криптофитовых, зеленых водорослей и представителей хромофитной группы (Dinophyta, Chrysophyta, Xanthophyta, Bacillariophyta, Phaeophyta), коррелирующих с составом фотосинтетических пигментов, послужило основанием для предположения о нескольких независимых актах симбиогенеза между гетеротрофными организмами и фотосинтезирующими прокариотами - предшественниками хлоропластов эукариотических растений. Описание нового отдела прокариотических водорослей (Prochlorophyta) (см. раздел 9.2) послужило новым подтверждением предположения о независимом возникновении симбиогенным путем различно окрашенных фотосинтезирующих эукариот [686]. В связи с открытием нуклеморфа возникло предположение, что предшественниками хлоропластов эукариот могли быть не только прокариотические, но и эукариотические внутриклеточные симбионты типа одноклеточных красных водорослей [562]. Предполагают, что предшественниками хлоропластов красных водорослей, обладающих фикобилиновыми пигментами, были синезеленые, зеленых водорослей и всех зеленых растений, для которых характерно наличие хлорофиллов а и b - прохлорофитовые, криптофитовых - одноклеточные красные водоросли, а представителей желтой пигментной группы - неизвестные в настоящее время прокариотические фотосинтетики с хлорофиллами а и с.
Все существующие в настоящее время системы водорослей несмотря на их огромное разнообразие имеют одно общее свойство. Все они являются иерархическими, имеют древовидный (или вееровидный) характер и основаны на принципе монофилии. Э. Майр [185] утверждает, что монофилия является одним из постулатов эволюционной систематики. Однако признание синтезогенеза как одного из путей эволюции органического мира [67, 199], возможности недивергентного происхождения таксонов в результате гибридизации, горизонтального переноса генов или симбиогенеза [90, 158, 192, 625] приведет к необходимости пересмотра формальной структуры системы на принципиально новой основе [197, 199].
Легко найти девушку по вкусу на https://zhigulevsksm.date благодаря удобному поиску. | Пользуйтесь удобным поиском на https://kamenkasm.date для лучшего выбора.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'