4.1.1. Абиотические факторы
К абиотическим факторам относятся: температура, свет, физические и химические свойства воды и субстрата, состояние и состав воздушных масс (что особенно важно для аэрофитных водорослей, живущих вне водных условий) и некоторые другие. Всю совокупность абиотических факторов можно, с известной долей условности, разделить на химические и физические.
Химические факторы
Вода, как лимитирующий фактор. Большую часть клетки водорослей составляет вода. Цитоплазма в среднем содержит 85-90% воды, и даже такие богатые липидами клеточные органеллы, как хлоропласты и митохондрии, содержат не менее 50% воды. Вода в растительной клетке существует в двух формах: конституционная вода, связанная водородными связями со структурами макромолекул, и резервная вода, не связанная, как правило, содержащаяся в вакуолях. В резервной воде обычно растворены сахара, различные органические кислоты и т. п., вследствие чего она может участвовать в стабилизации внутриклеточного осмотического давления. При полимеризации высокоактивных мелких молекул в макромолекулы (например, при превращении сахаров в крахмал) и при обратном процессе - гидролизе высокомолекулярных соединений, осмотическое давление в клетке способно быстро изменяться. Этот механизм обеспечивает устойчивость отдельных видов водорослей к высыханию и к резким колебаниям солености воды.
Для большинства водорослей вода - постоянная среда обитания, однако многие водоросли могут жить и вне воды. По устойчивости к высыханию, среди обитающих на суше растений выделяют (по Вальтеру) пойкилогидрические, - не способные поддерживать постоянное содержание воды в тканях, и гомойогидрические - способные поддерживать постоянную гидратацию тканей. У пойкилогидрических растений (синезеленые и некоторые зеленые водоросли) клетки при высыхании сжимаются без необратимого изменения ультраструктуры и, следовательно, не теряют жизнеспособности. При увлажнении они возобновляют нормальный метаболизм. Минимальная влажность, при которой возможна нормальная жизнедеятельность таких растений, различна. Ее значение предопределяет, в частности, распространение аэрофитов. Для гомойогидрических растений обязательно наличие крупной нейтральной вакуоли, с помощью которой стабилизируется водный запас клетки. Однако клетки с крупными вакуолями в значительной степени утрачивают способность к высыханию. К гомойогидрическим водорослям относятся, например, некоторые аэрофиты из зеленых и желтозеленых водорослей (Ulotrichales, Vaucheriales), обычно поселяющиеся в условиях постоянной избыточной увлажненности.
Соленость и минеральный состав воды. Это важнейшие лимитирующие факторы, влияющие на распределение водорослей.
Согласно международной классификации* основную массу природных водоемов составляют морские - эвгалинные, со средней соленостью 35‰. Среди континентальных водоемов преобладают пресноводные - агалинные, минерализация которых не превышает обычно 0,5%) (среди них встречаются и более минерализованные). Континентальные водоемы, объединяемые под названием минерализованные, очень разнообразны по степени минерализации: это и солоноватые, или миксогалинные, среди которых выделяют олигогалинные (с соленостью 0,5-5‰), мезогалинные (5-18‰) и полигалинные (18-30‰), а также эвгалинные (30-40‰), и ультрагалинные (не менее 40‰). Среди ультрагалинных нередко выделяют крайне засоленные - гипергалинные водоемы, концентрация солей в которых близка к предельной. Различны континентальные водоемы и по характеру минерализации. По классификации Алекина среди них выделяют гидрокарбонатные, сульфатные и хлоридные водоемы, которые в зависимости от степени и характера минерализации подразделяют на группы и типы.
* (Существует две основные классификации системы минерализации водоемов: 1) международная система классификации природных вод по общей солености, принятая в 1959 г. на Симпозиуме по классификации солоноватых вод в Венеции, и 2) классификация природных вод по ионному составу солей, разработанная О. А. Алекиным (1946) и широко применяемая в СССР [115]. )
В соответствии с упомянутыми классификациями водоемов и в зависимости от солеустойчивости водорослей, среди них выделяют олигогалинные, мезогалинные, эвгалинные, ультрагалинные, пресноводные и другие виды. Видовое богатство (численность видов) тесно связано с соленостью воды (табл. 4.1).
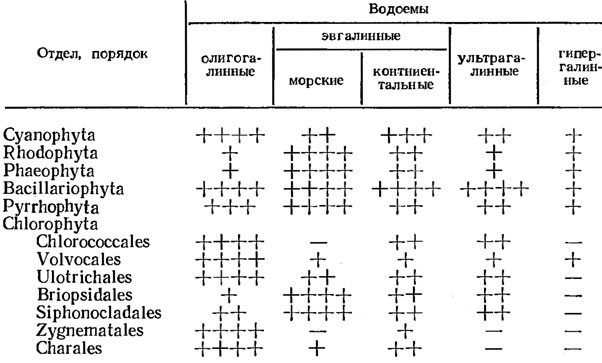
Таблица 4.1. Видовое богатство водорослей разных отделов в водоемах с различной соленостью
Примечание: ++++ - виды многочисленны; +++ - значительное число видов; ++ - ограниченное число видов; + - отдельные виды; - - виды не обнаружены (использована шкала Энглера).
Таблица 4.1 дает лишь самое приблизительное представление о видовом богатстве водорослей в водоемах различной солености. Практически в каждом из отделов можно найти виды, способные обитать в условиях крайнего засоления, и виды, живущие в водоемах с очень низкой минерализацией. Так, синезеленые водоросли - в подавляющем большинстве пресноводные организмы, однако среди них есть виды, способные развиваться в ультрагалинных водоемах (см. также раздел 4.2). Среди типично морских обитателей - золотистых водорослей порядка Coccolithophoridales - встречаются виды, распространенные и в континентальных водоемах с крайне низкой минерализацией. Диатомовые водоросли в целом в равной степени распространены и в морских и в континентальных водоемах; они встречаются в условиях с различной соленостью. Однако конкретные виды диатомовых нередко развиваются только при определенной солености и столь чувствительны к ее изменениям, что могут быть использованы как индикаторные организмы.
Очень чувствительны к изменениям солености и бурые водоросли. Многие из них не могут расти даже при незначительном опреснении. Поэтому они бедно представлены в водах Балтийского моря со сравнительно низкой соленостью. Сходную зависимость от степени солености водоема обнаруживают и красные водоросли: в Средиземном море (соленость 37-39 ‰) обнаружено более 300 видов красных водорослей, в Черном (17-18 ‰) - 129, в Каспийском (10 ‰) - 22 [122]. Зеленые водоросли преимущественно пресноводные организмы, лишь 10% из них встречаются в морях. Однако среди них имеются виды, способные выдерживать значительное засоление и даже вызывать "цветение" ультрагалинных водоемов (например, Dunaliella salina Teod., Asteromonas gracilis Artari).
Таким образом, водорослям в целом свойственна очень широкая амплитуда солеустойчивости. Что касается конкретных видов, то лишь немногие из них способны существовать в водоемах с разной соленостью, т. е. большинство водорослей - стеногалинные виды. Эвригалинных видов, способных существовать при разной солености, сравнительно немного (например, Bangia, Enteromorpha, Dunaliella).
Кислотность воды. Этот фактор также имеет большое значение для жизнедеятельности водорослей. Устойчивость разных таксонов водорослей к изменениям кислотности (pH) столь же различна, как и к изменениям солености. По отношению к кислотности среды выделяют виды, живущие в щелочных водах - алкалифит и, живущие в кислых водах, при низких значениях pH - ацидофилы. Ацидофилами, например, является большинство Desmidiales. Наибольшее видовое богатство десмидиевых водорослей наблюдается в эвтрофных и мезотрофных болотах, в условиях пониженной кислотности, однако некоторые десмидиевые могут встречаться и в щелочных водах с высокой минерализацией (например, Closterum acerosum (Schrank) Ehr., C. leibleinii Kütz.). Харовые, наоборот, преимущественно алкалифилы. Их наибольшее видовое разнообразие наблюдается в слабощелочных водах, однако некоторые из них (Chara vulgaris L. em. Wallr.) развиваются и в кислых водах, при pH 5,0.
Биогенные вещества. Наличие в среде макро- и микроэлементов, являющихся необходимыми компонентами тела водорослей, имеет решающее значение для интенсивности их развития.
Элементы и их соединения, относящиеся к макроэлементам (часто их называют макротрофными биогенными веществами), требуются организмам в сравнительно больших количествах. Особая роль среди них принадлежит азоту и фосфору. Азот входит в состав всех белковых молекул, а фосфор - обязательный компонент ядерного вещества, играющий значительную роль и в окислительно-восстановительных реакциях. Калий, кальций, сера и магний почти столь же необходимы, как азот и фосфор. Кальций в больших количествах используется морскими и пресноводными водорослями, отлагающими вокруг слоевищ "чехлы" из солей кальция (некоторые красные и харовые водоросли). Магний входит в состав хлорофилла, который является основным фотосинтезирующим пигментом водорослей большинства отделов.
Микроэлементы необходимы растениям в крайне малых количествах, но имеют огромное значение для их жизни, поскольку входят в состав многих жизненно важных ферментов. Причем, при небольшой потребности растений в микроэлементах их содержание в окружающей среде также незначительно. Микроэлементы нередко выступают как лимитирующие факторы. К ним относятся 10 элементов: железо, марганец, цинк, медь, бор, кремний, молибден, хлор, ванадий и кобальт. С физиологической точки зрения их можно разделить на три группы: 1) вещества, необходимые для фотосинтеза: марганец, железо, хлор, цинк и ванадий; 2) вещества, необходимые для азотного обмена: молибден, бор, кобальт, железо; 3) вещества, необходимые для других метаболических функций: марганец, бор, кобальт, медь и кремний.
Водоросли разных отделов имеют неодинаковые потребности в макро- и микроэлементах. Так, для нормального развития диатомовых водорослей необходимы довольно значительные количества кремния, который используется для постройки их панциря. При отсутствии или недостатке кремния панцири диатомовых истончаются, иногда до крайней степени.
Почти во всех пресноводных экосистемах к лимитирующим факторам относятся нитраты и фосфаты. В озерах и реках с мягкой водой к ним могут быть причислены также соли кальция и некоторые другие. В морских водоемах концентрация таких растворенных биогенных веществ, как нитраты, фосфаты и некоторые другие также низка, и они представляют собой лимитирующие факторы, в отличие от хлористого натрия и некоторых других солей. Низкие концентрации ряда биогенных веществ в морской воде, несмотря на то что они постоянно смываются в море, обусловлены тем, что время их жизни в растворенном состоянии довольно непродолжительно.
Физические факторы
Свет. Солнечное излучение имеет в жизни растений не меньшее значение, чем вода. Свет необходим растению как источник энергии фотохимических реакций и как регулятор развития. Его избыток, равно как и недостаток, может быть причиной серьезных нарушений развития водорослей. Следовательно, свет также является лимитирующим фактором при максимальной и минимальной освещенности. Каждый зависимый от солнечного излучения процесс осуществляется при участии определенных воспринимающих структур - акцепторов, в роли которых обычно выступают пигменты хлоропластов водорослей.
Распределение водорослей в толще воды в значительной степени определяется наличием света, необходимого для нормального фотосинтеза. Вода поглощает солнечное излучение гораздо сильнее, чем атмосфера. Длинноволновые тепловые лучи поглощаются уже у самой поверхности воды, инфракрасные - проникают в глубину на несколько сантиметров, ультрафиолетовые - на несколько дециметров (до метра), фотосинтетически активное излучение (длина световой волны около 500 нм) проникает до глубины 200 м.
Световой режим водоема зависит: 1) от условий освещения над поверхностью воды; 2) от степени отражения света ее поверхностью (при высоком стоянии солнца гладкая водная поверхность отражает в среднем 6% падающего света, при сильном волнении - около 10%, при низком стоянии солнца отражение столь значительно увеличивается, что большая часть света уже не проникает в воду: под водой день короче, чем на суше); 3) от степени поглощения и рассеивания лучей при прохождении через воду. С увеличением глубины освещенность резко убывает. Свет поглощается и рассеивается самой водой, растворенными веществами, взвешенными минеральными частицами, детритом и планктонными организмами. В мутных проточных водах уже на глубине 50 см освещенность такая же, как под пологом елового леса, где могут развиваться только самые теневыносливые виды высших растений, однако водоросли активно фотосинтезируют и на такой глубине. В прозрачных водах прикрепленные ко дну (бентосные) водоросли встречаются до глубины 30 м, а взвешенные в толще воды (планктонные) - до 140 м.
Слой воды выше границы обитания фотоавтотрофных организмов называется эвфотической зоной. В море граница эвфотической зоны обычно находится на глубине 60 м, изредка опускается до глубины 100-120 м, а в прозрачных водах океана - приблизительно до 140 м. В озерных, значительно менее прозрачных водах, граница этой зоны проходит на глубине 10-15 м, в наиболее прозрачных гляциальных и карстовых озерах - на глубине 20-30 м.
Оптимальные значения освещенности для разных видов водорослей варьируют в широких пределах. По отношению к свету выделяют гелиофильные и гелиофобные водоросли. Гелиофильные (светолюбивые) водоросли нуждаются в значительном количестве света для нормальной жизнедеятельности и фотосинтеза. К ним относится большинство синезеленых и значительное количество зеленых водорослей, обильно развивающихся в летнее время в поверхностных слоях воды. Гелиофобные (боящиеся, избегающие яркого света) водоросли приспособлены к условиям низкой освещенности. Например, большинство диатомовых водорослей избегает ярко освещенного поверхностного слоя воды и в малопрозрачных водах озер интенсивно развивается на глубине 2-3 м, а в прозрачных водах морей - на глубине 10-15 м. Однако далеко не все водоросли, живущие в условиях избыточной освещенности, нуждаются в больших количествах света, т. е. являются подлинно гелиофильными. Так, Dunaliella salina Teod. - обитатель открытых соленых водоемов и Trentepohlia jolitus (L.) Wallr., живущая на открытых скалах в горах, способные накапливать масла с избытком каротина, очевидно, играющие защитную роль, по сути являются не светолюбивыми, а светоустойчивыми организмами.
У водорослей разных отделов в зависимости от состава пигментов - фоторецепторов, максимальная интенсивность фотосинтеза наблюдается при разной длине световых волн. В наземных условиях качественные характеристики света довольно постоянны, так же как интенсивность фотосинтеза. При прохождении через воду свет красной и синей области спектра поглощается и на глубину проникает зеленоватый свет, слабо воспринимаемый хлорофиллом. Поэтому там выживают в основном красные и бурые водоросли, имеющие дополнительные фотосинтезирующие пигменты (фикоцианы, фикоэригрины и пр.), способные использовать энергию зеленого света. Отсюда становится ясным огромное влияние света на вертикальное распределение водорослей в морях и океанах: в приповерхностных слоях, как правило, преобладают зеленые водоросли, глубже - бурые, и на наиболее глубоководных участках - красные. Однако подобная закономерность не является абсолютной. Многие водоросли способны существовать в условиях крайне низкой, не свойственной им, освещенности, а иногда и в полной темноте. При этом у них могут происходить определенные изменения в пигментном составе или в способе питания. Так, у синезеленых водорослей в условиях низкой освещенности пигментный состав может изменяться в сторону преобладания фикобилинов (фикониана, фикоэритрина), цвет трихомов при этом меняется от синезеленого к пурпурному. Представители многих отделов водорослей (например, Euglenophyta, Chrysophyta) способны при отсутствии света и избытке органических веществ переходить к сапротрофному способу питания.
Движение воды. Огромную роль в жизни водорослей, обитателей водных биотопов, играет движение воды. Абсолютно стоячей, неподвижной воды не существует, и следовательно, практически все водоросли - обитатели текучих вод. В любых континентальных и морских водоемах наблюдается относительное движение водорослей и водных масс, обеспечивающее приток питательных веществ и удаление продуктов жизнедеятельности водорослей. Только в особых крайних условиях водоросли окружены постоянным слоем воды - в толще льда, на поверхности почвы, в пустотах скал, на других растениях и пр. Движение воды в результате ветрового перемешивания наблюдается даже в небольших лужах. В крупных озерах существуют постоянные приливно-отливные течения, а также вертикальное перемешивание. В морях и океанах, образующих по сути единую водную систему, кроме приливно-отливных явлений и вертикального перемешивания, наблюдаются постоянные течения, имеющие огромное значение в жизни водорослей.
Температура. Диапазон температур, в котором может сохраняться жизнь, сравнительно узок: -200 - +100°С. Водоросли являются организмами, которым свойственны, пожалуй, наиболее широкие диапазоны температурной устойчивости. Они способны существовать в крайних температурных условиях - в горячих источниках, температура которых близка к точке кипения воды, и на поверхности льда и снега, где температуры колеблются около 0°С.
По отношению к температурному фактору среди водорослей выделяют: эвритермные виды, существующие в широком температурном диапазоне (например, зеленые водоросли из порядка Oedogoniales, стерильные нити которых можно найти в мелких водоемах с ранней весны до поздней осени), и стенотермные, приспособленные к очень узким, иногда экстремальным температурным зонам. К стенотермным относятся, например, криофильные (холодолюбивые) водоросли, растущие только при температурах, близких к точке замерзания воды. На поверхности льда и снега можно встретить представителей различных таксонов водорослей: Desmidiales (например, Mesotaenium endlicherianum Näg., М. nivale Beck-Mann., Ancylonema nordenkoldii Bergr), Ulotrichales (Raphydonema nivale (Lagerh.) Chod., виды рода Koliella), Volvocales (Chlamydomonas nivalis) и др. В окрашенных снегах на Кавказе было обнаружено 55 видов водорослей, из которых 18 видов относились к зеленым, 10 - к синезеленым, 26 - к диатомовым и 1 вид - к красным [113]. В водах Арктики и Антарктики найдено 80 видов криофильных диатомовых водорослей. Всего известно около 100 видов водорослей, способных активно вегетировать на поверхности льда и снега. Эти виды объединяет способность выдерживать замерзание без нарушений тонких клеточных структур, и затем, при оттаивании, быстро возобновлять вегетацию, используя минимальное количество теплоты.
Водоросли, как уже указывалось выше, нередко выдерживают и высокие температуры, поселяясь в горячих источниках, гейзерах, вулканических озерах, в водоемах-охладителях промышленных предприятий и пр. Такие виды называются термофильными. Предельные температуры, при которых удавалось находить термофильные водоросли, судя по разным источникам, колеблются от 35-52 до 84°С и выше. Среди термофильных водорослей можно обнаружить представителей различных отделов, но подавляющее большинство их относится к синезеленым. Всего в горячих источниках обнаружено более 200 видов водорослей, однако облигатно термофильных видов среди них сравнительно немного. Большинство водорослей, обнаруженных в горячих источниках, способны выдерживать высокие температуры, но обильнее развиваются в условиях обычных температур, т. е. по сути являются мезотермными видами. Действительно термофильными можно считать только два вида: Mastigocladus laminosus Cohn, и Phormidium laminosum (C. Ag.) Gom., массовое развитие которых происходит при температуре 45-50°С. Основная масса водорослей в целом мезотермные организмы, но среди них всегда можно выделить более или менее термофильные, развивающиеся в определенных температурных диапазонах.
Отношение водорослей к температурному фактору влияет на их вертикальное распределение в водоемах. В различных водоемах и водотоках вследствие поглощения солнечного излучения верхними слоями воды прогреваются только эти слои. Теплая вода обладает меньшей плотностью, чем холодная, а вызываемые ветром течения выравнивают ее плотность только до определенной глубины. С началом вегетационного сезона, сезона интенсивного солнечного излучения, в достаточно глубоких континентальных непроточных водоемах возникает очень устойчивая температурная стратификация водных толщ. В этих водоемах образуются ограниченные друг от друга массы воды: теплый и легкий поверхностный слой - эпилимнион и лежащая под ним масса более холодной и плотной воды - гиполимнион. Осенью вода в водоеме охлаждается и температурная стратификация исчезает. В морях и океанах также имеется постоянный слой температурного скачка. Водоросли могут развиваться только в эпилимнионе (а именно в эвфотической зоне), причем наиболее теплолюбивые и светолюбивые организмы поселяются в поверхностных хорошо прогреваемых слоях воды.
Влияние температуры на водоросли, развивающиеся в водной среде, необычайно велико. Именно температура определяет их географическое распространение. Так, виды бурой водоросли рода Lessonia встречаются только в пределах летней изотермы 10°С, виды родов Laminaria, Agarum, Alaria не пересекают летней изотермы 20°С, некоторые виды Sargassum обитают только при температуре 22-23°С (Саргассово море). Даже в Балтийском море среди сообществ красных водорослей можно выделить менее теплолюбивые (Furcellaria, Delesseria, Dumontiа), обитающие при температурах ниже 4°С, и более теплолюбивые (Nemalion), обитающие при температурах выше 4°С. В целом, за исключением широко распространенных эвритермных видов (например, некоторые Fucales), в распространении водорослей наблюдается географическая зональность; конкретные токсоны морских планктонных и бентосных водорослей приурочены к определенным географическим поясам. Так, крупные бурые водоросли (Macrocystis) доминируют в северных морях. По мере продвижения к югу все более заметную роль начинают играть красные водоросли, а бурые отходят на второй план. Отношение количества видов красных и бурых водорослей в арктических морях составляет 1,5, в проливе Ла-Манш - 2, в Средиземном море - 3, а у Атлантического побережья Центральной Америки - 4,6. Данное отношение является важной характеристикой зональной принадлежности бентической флоры.
Лабораторные исследования показали, что температурная устойчивость конкретных видов красных и бурых водорослей довольно велика, но их интенсивный рост происходит в довольно узком температурном диапазоне.
Среди зеленых водорослей также известны более и менее теплолюбивые виды. Например, Caulerpa prolifera (Forsk.) Lamotif. и Cladophoropsis fasciculatus (Kjellm) Bθrg приурочены к экваториальной зоне мирового океана, a Codium ritteri S. et G., C. petrocelidus Kuck. - к северным широтам.
Хорошо выражена географическая зональность и у морских планктонных водорослей. Для морского тропического фитопланктона характерно значительное видовое богатство при очень низкой продуктивности. В планктоне тропических вод чрезвычайно богато представлены динофитовые и золотистые водоросли. Воды тропиков бедны диатомовыми водорослями, господствующими в северных морях.
Температурный фактор оказывает влияние и на вертикальное распределение морских планктонных и бентосных организмов.
Вертикальный оптимум произрастания морских водорослей как правило определяется комплексным воздействием теплового и светового режимов. Известно, что с понижением температуры интенсивность дыхания растений ослабевает быстрее, чем интенсивность фотосинтеза. Момент, когда процессы дыхания и фотосинтеза уравновешивают друг друга называется компенсационной точкой. Условия, при которых устанавливается компенсационная точка являются оптимальными для развития конкретных видов водорослей. В северных широтах в связи с низкой температурой компенсационная точка устанавливается на больших глубинах, чем в южных. Таким образом, нередки случаи, когда одни и те же виды водорослей встречаются в северных широтах на больших глубинах, чем в южных.
Очевидно, что температура влияет на географическое распределение этих (и других) водорослей в первую очередь косвенным образом - ускоряя или замедляя темпы роста отдельных видов, что приводит к их вытеснению другими, растущими более интенсивно в данном температурном режиме [113].
Все перечисленные абиотические факторы действуют на развитие и распределение водорослей в комплексе, компенсируя или дополняя друг друга.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'