2.2.2. Половой процесс, осуществляемый с помощью специализированных клеток - гамет
Половое размножение у водорослей, в том числе и у одноклеточных жгутиковых, гораздо чаще происходит путем деления содержимого клеток и образования в них специализированных половых клеток - гамет. У всех водорослей, кроме Conjugatophyceae из зеленых и красных водорослей, мужские гаметы обычно имеют жгутики, а женские гаметы имеют их не всегда. Образуются гаметы так же, как споры и зооспоры. У примитивных водорослей гаметы формируются в вегетативных клетках. У более высокоорганизованных они часто находятся в особых клетках - гаметангиях. В клетке или гаметангии может быть от одной до нескольких сотен гамет.
Размеры гамет, участвующих в слиянии, бывают разными. В зависимости от этого различают несколько типов полового процесса. Если сливающиеся гаметы имеют одинаковые размеры и форму, то такой половой процесс называют изогамным, или изогамией. Типичная изогамия встречается у одноклеточных (Pyrobotris gracilis Korsch., Stephanosphaera pluvialis Cohn) и колониальных (Gonium pectorale O. Müll.) представителей порядка Volvocales, а также у Chlamydomonadales. При половом размножении в клетках некоторых видов Chlamydomonadales образуется 32 или 64 изогаметы, похожие на зооспоры. Интересны изогаметы у Chlamydomonas proboscigera Korsch. Они имеют популяционные хоботки, выступающие через оболочку между основаниями жгутиков, которыми соприкасаются при слиянии.
Если сливающиеся гаметы имеют одинаковую форму, но разные размеры (женская гамета, крупнее мужской), то говорят о гетерогамном половом процессе, или гетерогамии (например, Chlamydomonas braunii Gorosch. и Pandorina morum Bory из зеленых). Половой процесс, при котором женская гамета, именуемая яйцеклеткой, лишена жгутиков, неподвижна и значительно крупнее мужской - сперматозоида или антерозоида, - называют оогамией (например, Chlamydomonas coccifera Gorosch. из зеленых). Гаметангии с яйцеклетками называют оогониями, а со сперматозоидами - сперматангиями, или чаще антеридиями.
У одноклеточных водорослей (например Ch. coccifera) оогамия протекает следующим образом: крупная женская гамета, превращаясь в яйцеклетку, становится неподвижной и оплодотворяется маленькой подвижной мужской гаметой. Образовавшаяся в результате слияния гамет зигота превращается в зигоспору с запасами питательных веществ и одевается многослойной целлюлозной оболочкой, в которой содержатся красный пигмент гематохром. При прорастании, сопровождаемом редукционным делением, образуется 4 зооспоры, развивающиеся в новые особи.
У колониальных видов (например, Volvox globator (L.) Ehr.) при оогамии яйцеклетки и сперматозоиды развиваются в оогониях и антеридиях. В грушевидных с темно-зеленым содержимым оогониях возникает только одна яйцеклетка. В антеридиях образуется по 64 желтоватых антерозоида, сложенных в виде пакетика. После оплодотворения яйцеклетка превращается в ооспору с многослойной шиповатой снаружи оболочкой и большими запасами питательных веществ. После периода покоя ооспора дает начало новой колонии V. globator.
Мужские и женские гаметы могут развиваться на одной особи или колонии (однодомные, или обоеполые виды) или на разных (двудомные, или раздельнополые виды). У водорослей с изогамным половым процессом существуют гомоталличные и гетероталличные виды. В первом случае сливаются гаметы из одного слоевища или колонии, во втором - из разных. При этом гамегы не различаются морфологически. Их обозначают знаками "плюс" и "минус" и соответственно называют плюс-гаметы (+ гаметы) и минус-гаметы (-гаметы).
Образующаяся в результате слияния гамет зигота после некоторых изменений превращается в зигоспору. Последняя обычно одета плотной одно- или многослойной оболочкой. В ней происходит слияние ядер гамет. У одних водорослей зигоспоры могут находиться в покое длительное время (до нескольких месяцев), у других - прорастают без периода покоя. Зигоспоры могут непосредственно давать начало новым слоевищам, либо в них происходит образование гаплоидных зооспор, которому предшествует мейотическое деление. Такие зооспоры предварительно растут, и из них, в зависимости от размеров, выходят 4 или 32 зооспоры.
У водорослей наблюдаются случаи партеногенетического развития (без оплодотворения) женских гамет. Последние морфологически напоминают зигоспоры. Обычно их называют азиготами, или партеноспорами.
Существует еще особый тип редуцированного полового процесса - автогамия. Он заключается в том, что ядро клетки предварительно претерпевает мейотическое деление, в результате которого образуются 4 ядра, 2 из которых разрушаются, а оставшиеся 2 ядра сливаются в диплоидное ядро, образуя зиготу, которая без периода покоя резко увеличивается в размерах и превращается в ауксоспору. Некоторое время ауксоспора связана с материнской клеткой. В зависимости от положения, которое занимает ауксоспора по отношению к последней различают свободную ауксоспору, конечную, боковую, интеркалярную и полуинтеркалярную. После созревания ауксоспоры в ней развивается новая клетка, которую называют инициальной. По размерам она значительно превосходит материнскую.
Таким образом, автогамия не сопровождается попарным слиянием двух клеток и увеличением числа особей, происходит лишь их омоложение; ауксоспора образуется из одной клетки. Ауксоспорообразование присуще только диатомовым водорослям (Centrophyceae).
У некоторых красных водорослей наблюдаются специфическая оогамия, не встречающая у других групп водорослей. Мужские и женские гаметы у них лишены жгутиков. При оплодотворении женские гаметы не покидают материнское растение-гаметофит, мужские же гаметы - спермации - выходят из гаметангия, называемого сперматангием, в воду и пассивно переносятся ее течением. Оогоний имеет очень сложное строение. Он представляет собой особую заполненную протоплазмой клетку - карпогон, лишенную хлоропласта. У некоторых примитивных красных водорослей карпогон снабжен окрашенными пластидами. Карпогон у представителей Florideophyceae имеет специфическую форму. Нижняя часть клетки, в которой находится ядро, имеет вид конуса, а верхняя - вытянута в длинный трубчатый вырост - трихогину, который служит для улавливания спермациев. Трихогина имеет длинную узкую спиралевидную или коротко-кеглевидную форму. У некоторых представителей Bangiophyceae трихогина еще не развита, а имеет вид коротких сосочковидных выростов - зачатков трихогины. У многих представителей Florideophyceae карпогон развивается как конечная клетка специальной карпогонной ветви, которая представляет собой короткую нить, состоящую из бесцветных клеток, заполненных гомогенной протоплазмой. Клетку, от которой отходит карпогонная ветвь, называют несущей, или базальной.
Карпогонная ветвь может образоваться как специальная боковая веточка во внутренней части корового слоя (Florideophyceae), либо как обычная боковая веточка слоевища (низкоорганизованные Nemaliales). У красных водорослей с плотным слоевищем карпогон погружен в слоевище, и только трихогина достигает его поверхности и выступает над ним, у нитчатых красных водорослей карпогон занимает открытое положение.
Карпогонные ветви играют важную роль в процессе размножения, так как принимают участие в развитии карпогона после оплодотворения. Число клеток в карпогонных ветвях у разных таксономических групп красных водорослей отражает степень их эволюционного развития. Строение карпогонной ветви и характер ее образования сильно варьирует у красных водорослей и является одним из основных систематических признаков.
Спермации представляют собой шаровидные или яйцевидные одноядерные клетки. У представителей Bangiophyceae - это голые клетки, а у Florideophyceae к моменту оседания на карпогон они покрываются тонкой оболочкой. У большинства красных водорослей спермации способны к амебоидному движению, чему способствует отсутствие оболочки.
Интересно отметить постепенную редукцию хлоропласта в спермациях у разных представителей красных водорослей. У просто организованных таксонов (Bangiophyceae), имеющих половой процесс, в спермациях развивается отчетливо видимый хлоропласт, но у высокоорганизованных представителей этого класса (например, у видов рода Porphyra Ag.), он уже сильно редуцирован. У низших представителей класса Florideophyceae остатки хлоропласта в мужских гаметах еще присутствуют, хотя не играют никакой роли в оплодотворении, спермации высокоорганизованных представителей этого класса хлоропластов, как правило, не имеют - они бесцветны.
Очень своеобразным половым процессом и специфическим строением половых органов характеризуются харовые водоросли. Женский половой орган - оогоний и мужской - антеридий многоклеточны и развиваются у большинства видов на одном растении, но известны и двудомные виды. Оогоний овальные, длиной до 1 мм, состоят из яйцеклетки и наружного ее покрова, стенки которого образованы пятью узкими клетками. Снизу оогоний снабжен одноклеточной ножкой, а сверху коронкой из 5 или 10 коротких клеток. Антеридии шаровидные, диаметром до 0,5 мм, образованные 8 плоскими, смыкающимися зазубренными краями клетками с отходящими внутрь отростками, на которых возникает множество мужских половых клеток - антерозоидов.
Антеридии и оогоний образуются из особых клеток, отчленяемых некоторыми периферическими клетками листовых узлов. Образование оогониев и антеридиев хорошо изучено. Формирование антеридия легко проследить у видов рода Nitella Ag. Начальная клетка, отчлененная от узловой, делится на 2, из которых наружная служит для образования собственно антеридия и быстро делится в трех взаимно перпендикулярных плоскостях на 8 клеток, слагающихся в шар, а из прилегающей к листовому узлу исходной клетки формируется основание антеридия (рис. 2.1). При дальнейшем делении клеток в верхней шаровидной части происходит обособление 8 наружных клеток, которые далее уже не делятся, и 8 внутренних, разделяющихся еще раз продольными перегородками. В итоге будущий антеридий оказывается состоящим из 8 внутренних клеток, плотно соприкасающихся друг с другом. Одновременно исходная клетка основания делится горизонтальной перегородкой на наружную базальную клетку, остающуюся без изменений, и внутреннюю базальную бутылковидную клетку, сильно разрастающуюся впоследствии и вдающуюся внутрь антеридия, образуя его ножку. Далее формируются 8 плоских покровных клеток трехугольной и четыхугольной формы с характерными лучистыми утолщениями оболочки - щитками, смыкающимися зазубренными краями. Примыкающие к ним изнутри клетки среднего ряда постепенно вытягиваются и превращаются в рукоятки, несущие на конце 8 округлых головок, образованных 8 внутренними клетками. При созревании антеридия эти основные головки образуют по 6 вторичных головок каждая, и на них развиваются длинные антеридиальные нити, по 4 на головке, состоящие из 200 расположенных в один ряд клеток. В каждой клетке образуется по 1 длинному спирально изогнутому антерозоиду, с 2 жгутиками на переднем конце. Каждый антеридий дает до 40000 антерозоидов.
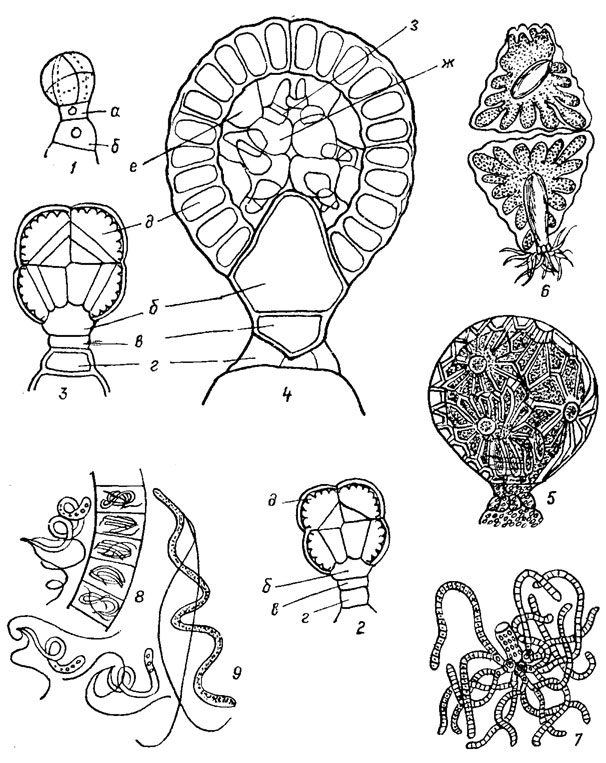
Рис. 2.1. Строение антеридия харовых водорослей: 1 - начальная восьмиклеточная стадия развития антеридия; 2, 3 - последующие стадии обособления наружных щитовых клеток и внутренней части (продольный срез) 4 - поздняя стадия развития с дифференцированной внутренней частью (продольный срез); 5 - зрелый антеридий; 6 - щитки с рукоятками (вид изнутри); 7 - рукоятки с развитыми антеридиальными нитями; 8 - участок антеридиальной нити с антерозоидами в клетках и снаружи; 9 - антерозоид при большом увеличении: а - исходная клетка основания; б - внутренняя базальная (бутылко-видная) клетка; в - наружная базальная клетка; г - клетка листового узла; д - стенка антеридия; е - рукоятка; ж - головки; з - антеридиальные нити
Зрелые антеридии у всех харовых имеют вид шарика на короткой ножке, диаметром в среднем около 0,5 мм, характерно пересеченного крест-накрест зигзагообразной линией смыкания щитков, в каждом из которых хорошо заметны по краям радиальные утолщения оболочки и в центре округлое основание внутренней рукоятки. Антеридии сначала окрашены в зеленый цвет от содержащихся в щитках хлоропластов, затем они становятся кирпич но-красными.
Формирование оогониев происходит проще. Одна из клеток листового узла, которая потом становится ножкой оогония, отчленяет 1 центральную клетку и в стороны вокруг себя - 5 будущих обволакивающих клеток (рис. 2.2). Центральная клетка делится один или два раза, в результате возникает крупная клетка, развивающаяся далее в яйцеклетку, и 1-3 мелкие стерильные клетки, получившие название поворотных, так как они, возникнув сбоку или сверху от яйцеклетки, постепенно перемещаются к ее основанию. Их считают сильно редуцированным остатком стенки собственно гаметангия. Одновременно с этим 5 боковых клеток разрастаются вокруг яйцеклетки вверх по спирали справа налево и полностью обволакивают ее. На самой вершине каждая из них отчленяет 1 или 2 небольшие клеточки, слагающие коронку. Зрелые оогонии имеют вид эллипсоидных или яйцевидных образований длиной до 1 мм, спирально исчерченных обволакивающими клетками и увенчанных небольшой или довольно крупной коронкой. Коронка из 10 мелких клеток, расположенных в два яруса характерна для Nitellaceae, одноярусная коронка из 5 крупных клеток - для Characeae.
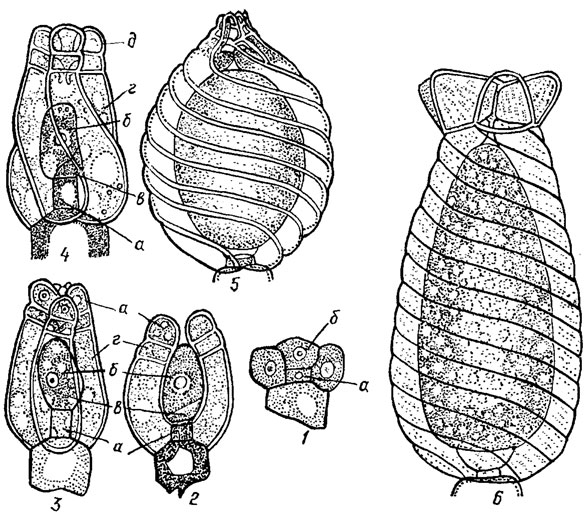
Рис. 2.2. Строение оогония харовых водорослей: 1 - начальная стадия развития оогония; 2-4 - последующие стадии развития оогония и обволакивающих спиральных клеток; 5, 6 - зрелые оогонии (5 - Nitella, 6 - Chara); а - клетка листового узла; 6 - оогонии; в - поворотные клетки; г - обволакивающие клетки; д - клетки коронки
Ооспоры развиваются внутри оогониев в результате оплодотворения яйцеклетки. Они имеют у разных видов разную форму и размеры - от почти шаровидных до узкоэллипсоидных, от очень мелких (около 0,3 мм) до крупных (почти 1 мм). По мере созревания наружная стенка ооспор затвердевает, пропитывается суберином и кремнеземом и окрашивается. Зрелые ооспоры бывают коричневато-желтыми, коричневыми или темно-коричневыми до черных, снаружи они всегда блестящие. У многих видов ооспоры покрываются еще слоем извести, а внутри заполняются питательными веществами (зернами крахмала и каплями жира).
После периода покоя ооспоры прорастают. Прорастание начинается с того, что в верхней части ооспоры грубые крахмальные зерна и капли жира постепенно исчезают, а диплоидное ядро делится дважды (первое деление сопровождается мейозом), в результате чего образуется 4 гаплоидных ядра. Далее верхний участок протоплазмы с одним ядром отделяется поперечной перегородкой, образуя линзообразную центральную клетку, от которой берет начало новое растение. Остальная часть ооспоры выполняет функцию склада запасного питательного материала. Первое деление центральной клетки происходит поперек и приводит к образованию двух функционально различных клеток. Из более крупной образуется стеблевой побег - предросток, из меньшей - первый ризоид. Предросток растет вверх и быстро зеленеет, заполняясь хлоропластами, а ризоид растет вниз и остается бесцветным.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'