1.2.2. Строение и деление клетки эукариотических водорослей (Е. Л. Кордюм)
(Форма, размеры и типы клеток эукариотических водорослей приведены при описании конкретных таксонов.)
Строение клетки
Клеточные покровы эукариотических водорослей, обеспечивающие резистентность содержимого клеток - протопластов к внешним воздействиям и придающие клеткам определенную форму, чрезвычайно разнообразны го структуре (рис. 1.15) и химическому составу. У большинства эукариотических водорослей независимо от морфологического типа структуры слоевища (монадный, гемимонадный, коккоидный, нитчатый, разнонитчатый, сифональный, паренхиматозный и др.) клетки окружены прочной полисахаридной оболочкой, проницаемой для воды и растворенных в ней низкомолекулярных веществ и легко пропускающей солнечный свет. Кроме полисахаридов в состав оболочки входят белки, гликопротеиды, минеральные соли, пигменты, липиды, вода. Повышение содержания воды в клеточной оболочке увеличивает ее проницаемость. В отличие от высших растений клеточные оболочки водорослей не содержат лигнина. Основным скелетным веществом оболочек клеток водорослей, как и высших растений, является целлюлоза, в химическом отношении представляющая собой β-1,4-D-глюкан. У некоторых водорослей микрофибриллы состоят из β-1,4-маннанов (семейства Codiaceae, Dasycladaceae, Derbesiaceae отдела Chlorophyta) или β-1,3-ксиланов (семейства Bryopsidaceae, Caulerpaceae, Udotaceae, Dichotomosiphonaceae отдела Chlorophyta; виды родов Porphyra Ag. и Bangia Lingb. отдела Rhodophyta).
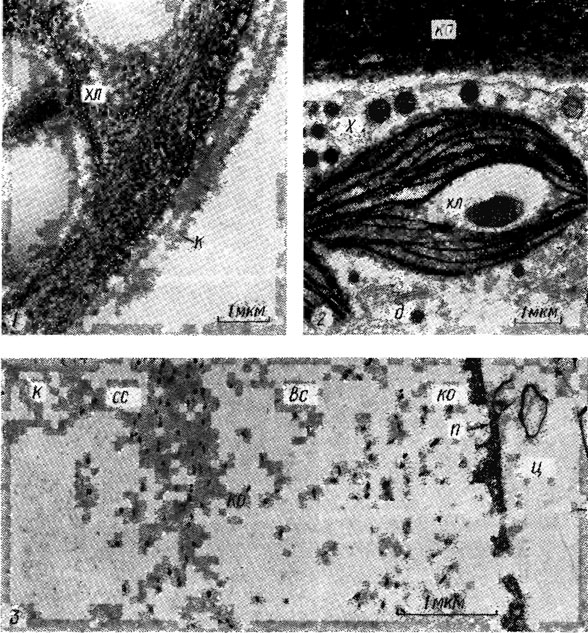
Рис. 1.15. Электронная микрофотография фрагментов клеток Chlorella vulgaris Beijer. (a), Chara vulgaris Vaill. (2) и клеточной оболочки Gracilaria verrucosa (Huds.) Papenf. (3): хл - хлоропласт; к - кутикула; ко - клеточная оболочка; х - харасома; сс - средний слой оболочки; вс - внутренний слой оболочки; п - плазмалемма, ц - цитоплазма; д - диктиосома
Нитчатые молекулы целлюлозы собраны в оболочках в структурные единицы, которые получили название микрофибрилл, составляющих каркас оболочки. Микрофибрилла состоит из нескольких десятков или сотен молекул целлюлозы, располагающихся параллельно друг другу по ее длине. Различают центральную кристаллическую часть микрофибриллы, в которой молекулы целлюлозы расположены в трехмерном пространстве так упорядочение что образуют кристаллическую решетку, и наружную - паракристаллическую. Длина микрофибрилл может достигать нескольких микрон. В поперечном разрезе микрофибрилла обычно имеет овальную форму. У отдельных видов Cladophorales и Siphonales длина большой и малой осей поперечного среза микрофибрилл составляет 18,5 и 11,5 нм, а их кристаллической части - 17 и 11 нм соответственно. На поперечном срезе кристаллической части насчитывается 500 молекул целлюлозы.
Микрофибриллы погружены в не имеющий кристаллической структуры матрикс - сложную смесь полимеров, среди которых преобладают полисахариды с различной молекулярной массой: глюкоза, галактоза, арабиноза, ксилоза, рамноза, манноза, фу коза, а также глюкуроновая и галактуроновая кислоты и др. Макромолекулы матрикса образуют гликозидные, ковалентные, водородные связи друг с другом и с микрофибриллами, что значительно повышает прочность оболочки.
Основную роль в образовании оболочки играют аппарат Гольджи и плазмалемма, которые содержат ферментные комплексы, участвующие в биосинтезе полисахаридов. Синтез полисахаридов матрикса оболочки (из Сахаров) начинается в цистернах диктиосом, совокупность которых и составляет аппарат Гольджи, и продолжается в пузырьках Гольджи после их отчленения от края цистерны и во время движения к плазмалемме. После встраивания мембран пузырьков Гольджи в плазмалемму их содержимое с полисахаридами и гликопротеидами матрикса изливается в оболочку.
Согласно современным представлениям микрофибриллы целлюлозы синтезируются на наружной поверхности плазмалеммы с помощью белковых терминальных комплексов двух типов - линейных и в виде розеток. Белковые терминальные комплексы встроены в плазмалемму и движутся в ее плоскости, оставляя после себя готовую микрофибриллу. Они содержат фермент целлюлозосинтетазу, катализирующий синтез целлюлозы. Образование и ориентация микрофибрилл в оболочке происходят одновременно; оба эти процесса контролируются протопластом. Линейные терминальные комплексы в плазмалемме описаны у некоторых видов водорослей (например, виды родов Oocystis Näg., Glaucocystis Itzigs., Boodlea Mur., Chaetomorha Kütz., а также Valonia macrophysa Kütz.), для которых характерны микрофибриллы относительно большого диаметра и высокой кристалличности (рис. 1.16, 1). Количество субъединиц в этих комплексах соответствует числу глюкановых цепочек в микрофибриллах. Терминальные комплексы в виде розеток, состоящих из 6 гранул размером 8 им, с центральной глобулой, обнаружены в плазмалемме у видов родов Micrasterias Ag., Closterium Nitzsch. и Spirogyra Lynk (рис. 1.16, 2, 3). Подобного рода розетки выявлены и на сколе мембран пузырьков Гольджи. Предполагают, что здесь происходит сборка розеток, которые находятся в неактивном состоянии до тех пор, пока не произойдет встраивание мембраны пузырьков в плазмалемму при экзоцитозе.
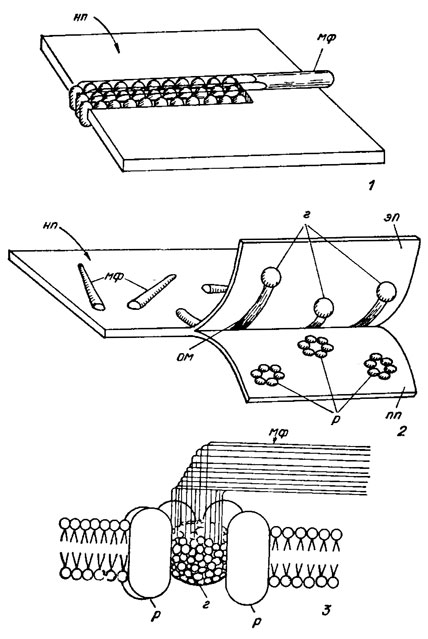
Рис. 1.16. Схемы линейного терминального комплекса (1) и терминального комплекса в виде розеток (2); схема соединения глобула - розетка (3); нп - наружная поверхность плазмалеммы; мф - микрофибрилла; г - глобула; эп - экзоплазм этическая поверхность скола; ом - отпечаток микрофибриллы; пп - протоплазм этическая поверхность скола, р - розетка
Толщина и количество слоев оболочки клеток водорослей неодинаковы у разных видов. В большинстве случаев оболочка состоит из 2-3, реже 4 и более слоев. Толщина оболочки варьирует также в зависимости от возраста и состояния клеток, условий выращивания. Микрофибриллы целлюлозы по-разному ориентированы в различных слоях клеточной оболочки. Так, в первом слое оболочки сифоновой водоросли из рода Valonia, образующемся вокруг голой апланоспоры, редкие микрофибриллы целлюлозы расположены беспорядочно. В последующих слоях оболочки уже зрелого пузырька, прикрепленного к субстрату базальными ризоидами, описаны три типа спиральной ориентации микрофибрилл целлюлозы - две левовращающие спирали, составляющие углы в 80 и 10° с большой осью пузырька, и правовращающая спираль, которая делит пополам угол между двумя спиралями (рис. 1.17). Все спирали идут от одного полюса пузырька к другому.
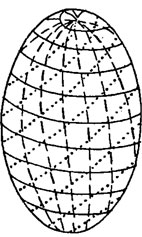
Рис. 1.17. Типы спиральной ориентации микрофибрилл в клеточной оболочке Valonia ventricosa
Оболочка клеток водорослей пронизана порами, размеры которых колеблются в значительных пределах - от 12 до 60 и даже 90 нм. Через поры осуществляется связь протопласта с внешней средой у одноклеточных водорослей, у многоклеточных - также и с соседними клетками слоевища с помощью плазмодесм, которые возникают в процессе цитокинеза и представляют каналы диаметром 25-80 нм, выстланные плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы обычно проходит трубка (десмотрубка), стенки которой непрерывны с мембранами эндоплазматического ретикулюма обеих клеток. После завершения цитокинеза путем локального ферментативного лизиса отдельных микроскопических участков внешней оболочки клетки образуются вторичные поры.
У многих водорослей клеточные оболочки пропитываются солями железа (некоторые вольвоксовые и красные водоросли), кальция (представители харовых, зеленых, бурых и красных водорослей), кремния (многие золотистые, желтозеленые, бурые и диатомовые водоросли). У последних соли создают разнообразную архитектонику поверхностных слоев - своеобразный кремниевый панцирь.
Дополнительный твердый покров - кутикула - на поверхности оболочки ряда видов зеленых, красных и бурых водорослей (например, Oedogonium Link, Caulerpa Lamour., Halimeda Lamour., Udotea Lamour., Porphyra Ag.) образуется за счет выделяемого клеткой кутина. Кутикула может быть гомогенной, электронно-плотной, 20-50 нм толщиной, или слоистой. Так, у видов родов Aurainvillea и Cladocephalus (Siphonophyceae) кутикула толщиной 16-20 нм состоит из трех слоев: двух электронно-плотных (наружного и внутреннего по 4-5 нм толщиной) и среднего, менее электронно-плотного, аморфного, слоя.
Поверхность оболочки может быть также покрыта слоем спорополленина - продукта полимеризации каротиноидов, устойчивого к воздействию ферментов, кислот и щелочей (некоторые виды рода Chlorella Beijer. и др.).
Мощные слоистые чехлы, состоящие из специфических полисахаридов, в частности сульфатированных полисахаридов, пектинов, фукоидина, характерны для многих бурых, красных и зеленых водорослей.
Образование слизистых капсул вокруг клеток из секретируемой клетками слизи отмечено у Volvocales и Chlorococcales, нередко представленных колониальными формами. В неблагоприятных условиях некоторые водоросли (например, Chlamydomonas Ehr.) переходят временно в пальмеллевидное состояние. При этом их клеточные оболочки вследствие гидратации матрикса ослизняются и слипаются, что приводит к образованию больших клеточных скоплений.
У ряда видов водорослей целлюлозная оболочка отсутствует и клеточные покровы представлены преимущественно белковыми слоями - пелликулой, перипластом, текой, располагающимися под плазма леммой, реже над ней. Структура слоев варьирует у водорослей различных отделов. Так, пелликула эвгленовых водорослей - плотный, эластичный слой протопласта толщиной до 70 нм, состоящий приблизительно из 20 полипептидов и тесно прилегающий к плазмалемме. Поверхность таких клеток продольно-ребристая, причем ребра начинаются от глотки и по спирали огибают всю клетку или располагаются более или менее параллельно длинной оси клетки (рис. 1.18, 1). На поперечном срезе ребра клетки имеют вид гребней с крючковатым изгибом, перемежающихся с глубокими бороздами (рис. 1.18, 2-5). В основании изгиба находится электронно-плотный морфогенетический центр, ассоциированный с микротрубочками. Делению клетки предшествует удвоение полос, начинающееся с удвоения морфогенетического центра.
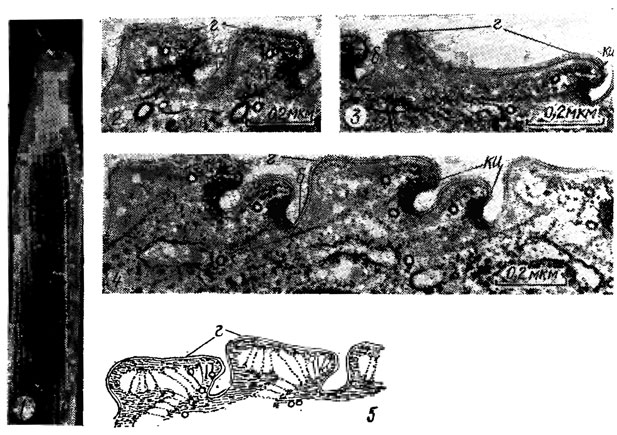
Рис. 1.18. Электронная микрофотография клетки Cyclidiopsis acus Kors., не имеющей целлюлозной оболочки (1), и срезов верхней и средней частей этой клетки (2-4); схема участка пелликулы верхней части клетки (5); г - гребень; б - борозда; ки - крючковидный изгиб
Клеточный покров криптофитовых водорослей - перипласт - состоит из внутреннего белкового компонента и дополнительного внешнего, располагающегося над плазма леммой. Внутренние и внешние пластинчатые структуры одинакового размера и толщины тесно прилегают к плазмалемме. Прямоугольные или шестиугольные пластинки собраны в четкие ряды; пузырьки, содержащие эджективные (стрекательные) органеллы - эджектосомы обычно располагаются в углах пластинок. Поверхность клеток покрыта семиугольными чешуйками; каждая чешуйка диаметром 120-140 нм состоит из центральной оси, от которой к периферии из переплетенного фибриллярного материала отходит 7 полосок. Кроме чешуек, на поверхности клеток наблюдаются отдельные фибриллы 10 нм шириной и до 1,5 мкм длиной (рис. 1.19, 1-4). Перипласт пронизан порами.
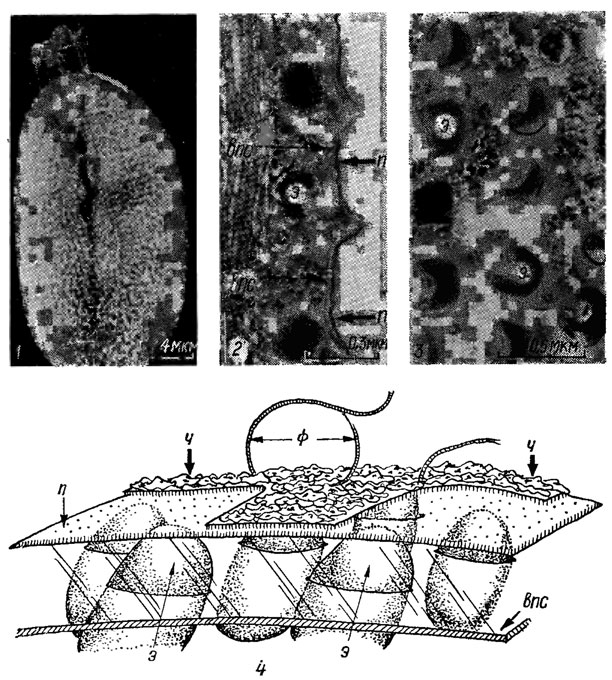
Рис. 1.19. Электронная микрофотография клетки криптофитовой водоросли Cryptomonas cryophila (1) и ее фрагментов (2-3); схема участка перипласта (4): п - плазмалемма; э - эджектосома; впс - внутренняя пластинчатая структура перипласта: ч - чешуйка; ф - фибрилла
Тека динофитовых водорослей, кроме белков, содержит галактозу, арабинозу и галактуроновую кислоту и является многокомпонентной системой. Основной элемент теки - уплощенные пузырьки длиной от 600-800 нм до 0,8-1,2 мкм и толщиной 60-170 нм; они располагаются непосредственно под плазмалеммой и окружены мембраной; их число может варьировать от 2 на клетку до нескольких сотен. Пузырьки заполнены зернистым веществом или содержат одиночные пластинки, которые по структуре и величине весьма разнообразны у разных видов динофитовых водорослей и часто срастаются своими краями, образуя прочный панцирь. К мембране пузырьков со стороны протопласта могут прилегать дополнительные структуры в виде пробок шириной 200 нм с неоднородным содержимым (рис. 1.20, 1, 2). Поверхность клеток различается по текстуре; у ряда видов образуются шипы длиною до 600 нм. Тека пронизана порами.
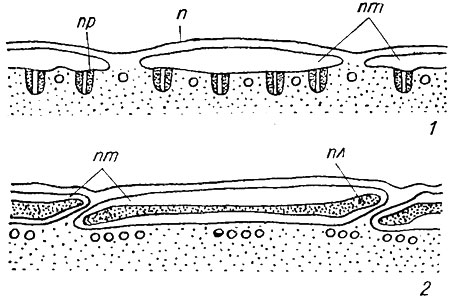
Рис. 1.20. Схема участков теки видов родов Gymnodinium Stein (1) и Glenodinium (Ehr.) Stein (2): пр - пробка; п - плазмалемма; пт - пузырьки трихоциста; пл - пластинка
Эджективные органеллы динофитовых водорослей - трихоцисты - имеют более сложное строение, чем эджектосомы криптофитовых. В одной клетке может находиться несколько сотен трихоцист. Они окружены мембраной и состоят из паракристаллического тела и более узкой фиброзной части. Зрелые трихоцисты обычно располагаются по периферии клетки под порами и при раздражении выстреливают наружу через пору длинную поперечно исчерченную нить ромбовидной или четырехугольной формы, 60-180 нм шириной.
У некоторых видов динофитовых типичные трихоцисты отсутствуют, а выполняющие аналогичную функцию, но отличающиеся по строению органеллы называют рабдосомами (рис. 1.21, 1). Они состоят из цилиндрической структуры, погруженной в свободный футляр различных размеров. Передний конец цилиндрической структуры несколько заострен, около 4 мкм в длину и 150-200 нм в ширину, задний конец округлен. Цилиндрическая структура может быть заполнена аморфным веществом (рис. 1.21, 2) либо содержит несколько продольных элементов и аморфное вещество (рис. 1.21, 3) или состоит из 8 нитей 30 нм толщиной, свободно переплетающихся между собой (рис. 1.21, 4).
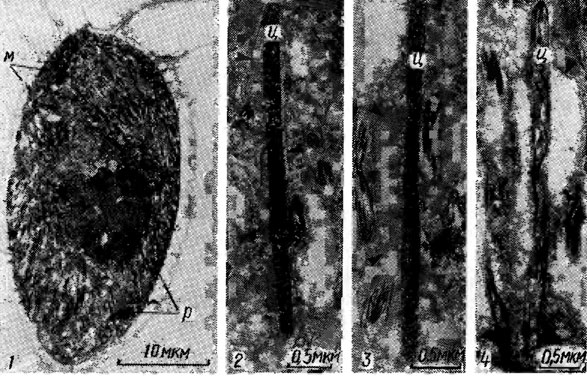
Рис. 1.21. Электронная микрофотография клетки Dinophysis acuminata Clap et Lachm. (1) и ее фрагментов (2-4): м - мукоциста; р - рабдосома; ц - цилиндрическая структура (стрелками указаны границы футляра)
Эджективные органеллы, выполняющие скорее всего защитную функцию, известны также у золотистых (здесь они называются дискоболоцистами), зеленых и эвгленовых водорослей.
Поверхность клеток многих водорослей, особенно одноклеточных, покрыта, как уже отмечалось, чешуйками, а также шипиками и щетинками. Последние представляют собой более тонкие и длинные, до 200 мкм, выросты по сравнению с короткими и толстыми шипиками. Эти дополнительные структуры выполняют разнообразные функции. У одноклеточных образование щетинок и шипиков скорее всего является приспособлением к планктонному образу жизни.
У незначительного числа видов золотистых и зеленых водорослей клеточные покровы не формируются, и самым внешним слоем клеток является плазмалемма. Снаружи эти, т. н. голые клетки большей частью покрыты слизью. В искусственных условиях получены мутантные формы Chlamydomonas reinhardtii Dang., лишенные клеточной оболочки, которые интенсивно растут в культуре. Кратковременное отсутствие клеточных покровов характерно для гамет и зооспор одноклеточных и многоклеточных водорослей, для первых - от момента образования до осуществления процесса оплодотворения, для вторых - до формирования клеточных покровов.
Движение жгутиковых форм, зооспор и гамет неклеточных (сифоновых) и многоклеточных водорослей осуществляется в подавляющем большинстве случаев с помощью жгутикового аппарата - сложной системы, состоящей из собственно жгутиков (ундулиподий), базальных тел (кинетосом) и корней жгутиков (рис. 1.22). Жгутики водорослей, так же как и реснички простейших, способны совершать ритмические волнообразные движения (по ним обычно пробегают квазисинусоидальные волны); на этой основе жгутики и реснички объединяют под общим названием ундулиподий. Термин ундулиподия в переводе с латинского означает "волноножка". Среди водорослей чаще всего встречаются 1- и 2-жгутиковые формы, реже 3-, 4- и 8-жгутиковые; у зооспор Oedogonium Link, Derbesia Sol., Vaucheria D. С. число жгутиков достигает нескольких десятков.
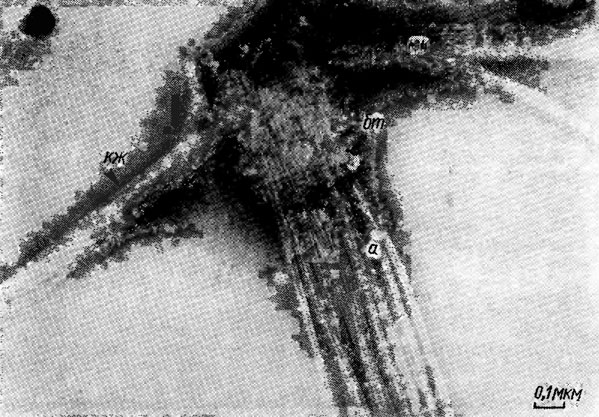
Рис. 1.22. Электронная микрофотография изолированного жгутикового аппарата Dunaliella bioculata Butch.: кж - корень жгутика; бт - базальное тело; а - аксонема
По соотношению длины жгутиков в пределах одной клетки водоросли разделяют на две группы - равножгутиковые и разножгутиковые. У последних жгутики часто занимают неодинаковое положение по отношению к клетке и имеют различия в частоте и интенсивности биений - длинный жгутик направлен вперед и работает энергичнее, чем короткий, который обращен в сторону или назад по ходу движения клетки.
Основные черты внутреннего строения жгутиков и базальных тел вегетативных клеток жгутиковых форм, зооспор и гамет сходны. Жгутик окружен мембраной, которая является продолжением плазма леммы клетки. Внутри жгутика находится матрикс, аналогичный матриксу клетки - гиалоплазме, и аксонема - осевая структура, изгибанием которой и обусловлено биение жгутика. Аксонема представляет собой сложный комплекс микротрубочек (внутренний диаметр 25 нм), образованных белком тубулином, и связанных с ними других белков. Тубулин является димером и образован двумя полипептидами - α- и β-тубулином, которые весьма сходны по аминокислотной последовательности. При формировании микротрубочек молекулы тубулина соединяются друг с другом в протофиламенты - нитевидные комплексы, в которых α-тубулин одного димера связан с β-тубулином следующего. Обычно 13 таких протофиламентов, расположенных параллельно и прилегающих друг к другу, образуют цилиндрическую структуру - собственно микротрубочку. Так как тубулиновые полипептиды в соседних протофиламентах несколько смещены относительно друг друга, то в стенке цилиндра они образуют регулярно чередующиеся спиральные ряды (рис. 1.23, 1).
Аксонема состоит из 9 пар периферических микротрубочек и 2 центральных (рис. 1.23, 2; 1.24). Структура "9 + 2" характерна для жгутиков и ресничек почти всех эукариотических организмов, хотя имеются и отклонения. Центральная пара микротрубочек заключена в центральную капсулу, образованную тонкими загнутыми белковыми отростками, прикрепленными к микротрубочкам. Периферические микротрубочки дуплетов несколько различаются по своей структуре - одна из них полная (субфибрилла А) построена из 13 субъединиц, вторая - неполная (субфибрилла В) - только из 10. Субфибрилла А по всей длине несет динеиновые "ручки" - короткие боковые выступы, отходящие от субфибриллы А каждого дуплета по направлению к соседнему с интервалами 24 нм и состоящие из белка динеина. Динеиновые "ручки" необходимы для изгибания жгутиков, которое обусловлено АТФ-зависимым скольжением микротрубочек аксонемы. Динеин состоит из многих полипептидов и сам по себе действует как АТФаза. Каждая диненновая "ручка" развивает примерно такое же усилие, как одна молекула миозина в сокращающейся мышце. Сила прикладывается к соседнему дуплету микротрубочек, с которым контактируют "ручки", и стремится сдвинуть его по направлению к кончику жгутика.
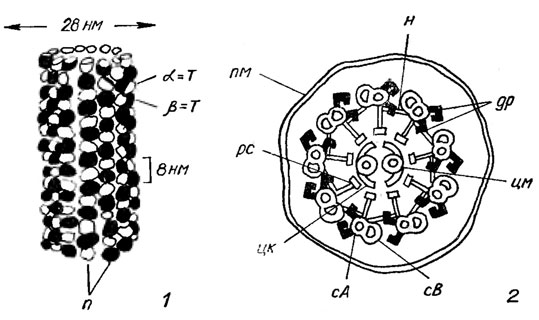
Рис. 1.23. Схема строения микротрубочки (1) и поперечного среза жгутика Chlamydomonas (2): α-m, β-m - α и β - тубулин; m - протофиламенты: н - нексин; др - динеиновые 'ручки'; цм - центральная микротрубочка; сА и сВ - субфибриллы А и В, образующие периферический дуплет , цк - центральная капсула; рс - радиальная спица, пм - плазматическая мембрана
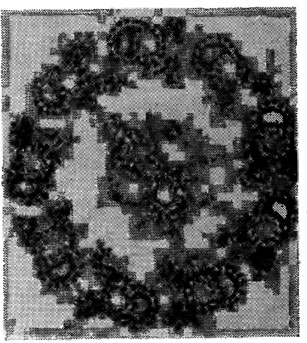
Рис. 1.24. Электронная микрофотография поперечного среза жгутика Chlamydomonas sp
Поперечные эластичные связи между соседними дуплетами микротрубочек образованы белком нексином, благодаря чему аксонема по окружности оказывается как бы скрепленной обручем. От каждого дуплета внутрь аксонемы отходят радиальная "спица", заканчивающаяся глобулярным образованием в непосредственной близости от центральной капсулы (см. рис. 1.23, 2), которая, по-видимому, способствует выполнению центральными микротрубочками регуляторной функции при движении аксонемы.
Поверхность жгутиков может быть гладкой или опушенной за счет латеральных выростов, или мастигонем, длиной 2-3 мкм, имеющих вид тонких волосков, диаметр которых варьирует у разных видов водорослей от 5 (Pedinomonas Korsch.) до 17-19 (Fucus L. и Ascophyllum Stack.) и даже до 25 нм (некоторые желто-зеленые водоросли). Число рядов, длина мастигонем и их расположение могут различаться у жгутиков одной клетки. Как правило, у равножгутиковых водорослей оба жгутика опушены; у разножгутиковых опушен преимущественно двигательный, передний, жгутик (рис. 1.25). У некоторых зеленых и бурых жгутик оканчивается тонким бичеподобным выростом - акронемой. Поверхность жгутиков у ряда видов зеленых, золотистых и бурых водорослей покрыта плотными неминерализованными чешуйками различной формы: пластинчатой, дисковидной, кольцевидной, булавовидной и др. Жгутиковые чешуйки, как и чешуйки клеточных покровов, образуются в пузырьках аппарата Гольджи, транспортируются к периферии жгутика и затем выводятся на его поверхность.
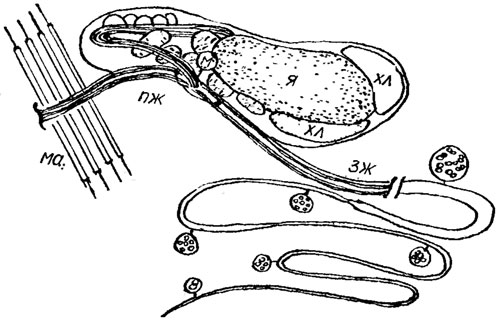
Рис. 1.25. Схема строения спермия Laminaria Lam.: ма - мастигонема; пою - передний жгутик; м - митохондрия; я - ядро; хл - хлоропласт; зж - задний жгутик
Базальные тела, или кинетосомы, жгутиков идентичны по своей структуре центриолям и во многих случаях могут превращаться друг в друга. Центриоли обычно представлены парами, члены которых расположены под прямым углом друг к другу (рис. 1.26, 1). Каждая центриоль состоит из 9 триплетов микротрубочек (рис. 1.26, 2, 3). Новая пара центриолей образуется по соседству с уже существующей, и этот процесс протекает сходным образом у всех эукариотических организмов. Одной из основных функций центриоли (а согласно крайней точке зрения - единственной) является образование жгутиков. Например, у Chlamydomonas reinhardtii Dang, в начале митоза жгутики исчезают, а базальные тела перемещаются в глубь клетки, ближе к ядру, где служат центрами организации митотического веретена деления. По окончании митоза центриоли снова становятся базальными телами, от которых отрастают жгутики.
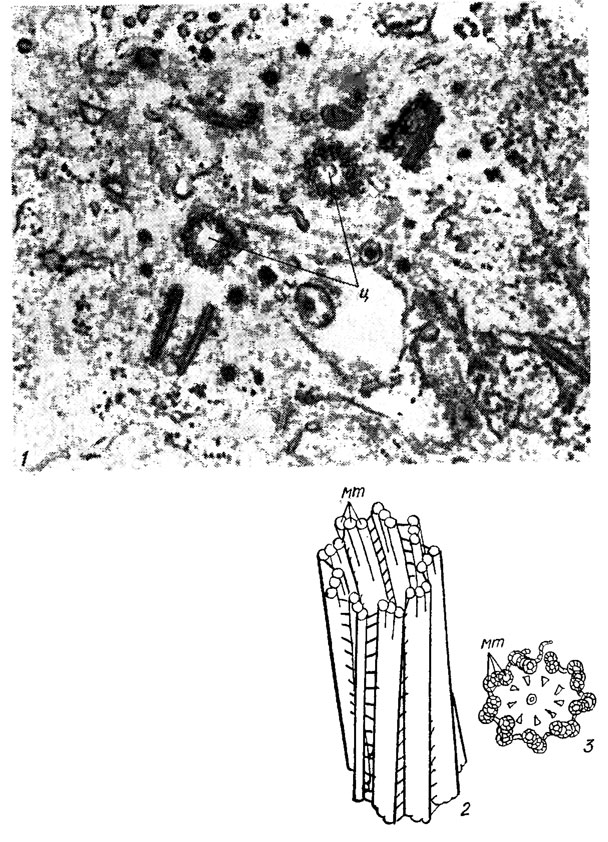
Рис. 1.26. Электронная микрофотография фрагмента клетки эукариотической водоросли с двумя парами центр нолей (1); схема строения центриоли (2 - общий вид, 3 - поперечный срез): ц - центриоль: мт - микротрубочка
Базальное тело имеет форму короткого цилиндра, стенки которого образованы 9 триплетами продольно расположенных микротрубочек; внешний диаметр базального тела равен диаметру аксонемы. Каждый из 9 периферических дуплетов аксонемы продолжается в базальное тело, где к нему присоединяется третья неполная микротрубочка. Обе центральные микротрубочки аксонемы оканчиваются выше базального тела; центр базального тела лишен микротрубочек (рис. 1.27). Микротрубочковые корни жгутиков, выходящие из базальных тел и состоящие преимущественно из 2-4 микротрубочек, у большинства видов располагаются симметрично, образуя характерные крестообразные фигуры.
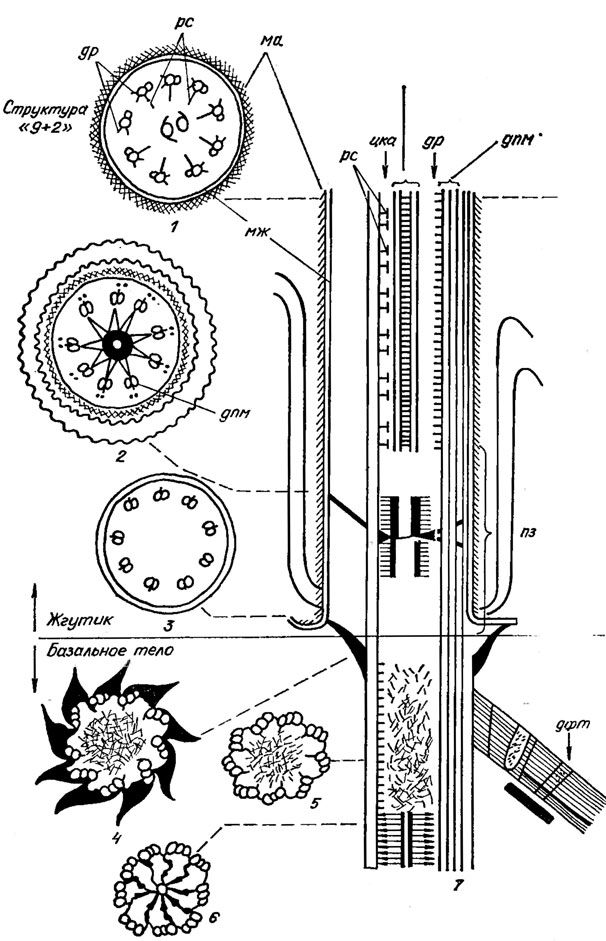
Рис. 1.27. Схема строения жгутика Chlamydomonas reinhardtii Dang, в поперечном (1-6) и продольном (7) разрезах: рс - радиальные спицы; ма - мастигонема; цка - центральная капсула аксонемы; др - динеиновые 'ручки'; дпм - дуплеты периферических микротрубочек аксонемы; мж - мембрана жгутика; пз - переходная зона; дфт - дистальный фибриллярный тяж
Среди фибриллярных структур, ассоциированных с базальными телами, выделено два основных типа: связывающие тяжи, которые связывают различные базальные тела; фибриллярные корни, возникающие при базальных телах и оканчивающиеся в различных частях клетки. Известно два типа фибриллярных корней: исчерченные тяжи, ассоциированные с микротрубочковым корнем жгутика (I система тяжей), в которых полосы чередуются с расстоянием 25-35 нм; исчерченные тяжи, состоящие из пучков тонких филаментов диаметром 5-10 нм (II система тяжей), чередующихся с расстоянием более 80 нм. Число, расположение и субструктура связывающих тяжей и фибриллярных корней значительно варьируют у разных видов водорослей.
У некоторых зеленых и харовых водорослей (Coleochaete Breb. Kiebsormidium Silva, Mattox et Bla Jewell, Chara Vaill., NitellaAg., Trentepohlia Mart.) корневая система жгутиков имеет асимметричное строение, обусловленное наличием т. н. многослойной структуры (например, сперматозоиды многоклеточных зеленых и харовых водорослей с оогамным половым процессом). Многослойная структура имеет сложное строение (рис. 1.28). Ее верхний слой, толщиной 25 нм, состоит из 22-25 тесно расположенных микротрубочек. Под ними находятся два слоя филаментов. В основании структуры лежит электронно-плотная пластинка толщиной около 5 нм. Размер многослойной структуры 600 × 360 × 90 нм. Дополнительные элементы, ассоциированные со жгутиковым аппаратом, четко оформлены в виде многослойной структуры и в сперматозоидах высших растений.
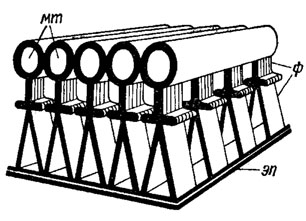
Рис. 1.28. Строение многослойной структуры Heterosigma akashiwo (Hada) Hada: мт - микротрубочки; ф - филаменты; эп - электронно-плотная пластинка основания
Клетки некоторых видов золотистых водорослей, помимо жгутиков, имеют направленный вперед вырост - гаптонему* (рис. 1.29). По длине гаптонема может быть короче, равна или длиннее жгутиков. Например, у разножгутиковой Chrysochromulina apheles длина жгутиков и гаптонемы составляет 7-10; 11-55 и 20-40 мкм соответственно. Описаны два основные типа медленного движения клеток с участием гаптонемы: 1) гаптонема у плавающей клетки направлена вперед, два жгутика со слабым биением держатся почти перпендикулярно гаптонеме, несколько отклоняясь назад, тело клетки при этом не вращается (рис. 1.30); 2) гаптонема направлена назад, жгутнки со слабым биением также направлены назад под острым углом к гаптонеме (рис. 1.30), тело клетки медленно вращается вокруг продольной оси. Этот тип движения может резко переходить в более быстрое плавание. При этом вытянутая гаптонема свертывается в плотно упакованные кольца и ритм биения жгутиков возрастает. Тело клетки вращается в необычном направлении вдоль широкой спиральной траектории (рис. 1.30). Не вызывает сомнения, что необычное движение вызвано свернувшейся гаптонемой, которая меняет центр тяжести клетки.
* (Гаптонема является одним из признаков, на основании которого в отделе золотистых водорослей в настоящее время выделен самостоятельный класс Haptophyceae.)
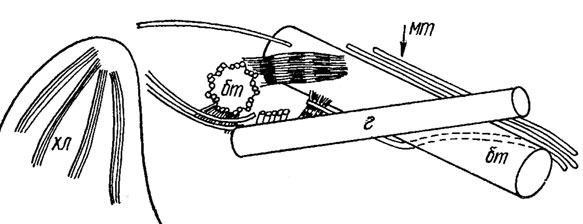
Рис. 1.29. Схема жгутикового аппарата Chrysochromulina apheles с гаптонемой: хл - хлоропласт; бт - базальное тело; г - гаптонема; мт - микротрубочка
Гаптонема состоит из 6 микротрубочек, собранных в круг в ее свободной части и окруженных частично или полностью эндоплазматическим ретикулюмом. В основании гаптонемы число микротрубочек может увеличиваться. Проксимальный свободный конец гаптонемы несколько вздувается, и в этом районе микротрубочки ориентированы в виде полукруга или арки. Два базальных тела прилегают друг к другу и к гаптонеме в высшей степени асимметрично. Четыре микротрубочковых корня образуются в районе базального тела и гаптонемы (см. рис. 1.29).
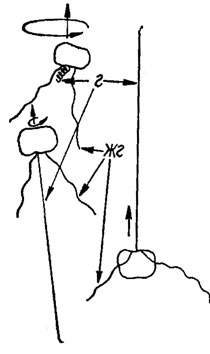
Рис. 1.30. Плавающие клетки Chrysochromulina apheles: г - гаптонема: жг - жгутик
Подавляющее большинство одноклеточных водорослей представлено жгутиковыми формами, однако среди наиболее примитивных одноклеточных встречаются и ризоподиальные, причем некоторые жгутиковые могут терять жгутики и превращаться в ризопод, а часть типичных ризопод обладает способностью к образованию жгутиков.
Клетки эукариотических водорослей, как и высших растений, содержат следующие основные органеллы - ядро, эндоплазматический ретикулюм, аппарат Гольджи, митохондрии, хлоропласты, вакуоли, пероксисомы. В то же время, в отличие от высших растений, в клетках различных форм водорослей присутствуют центриоли и пиреноиды, а у одноклеточных - стигма, сократительные (пульсирующие) вакуоли и некоторые другие специфические образования (рис. 1.31). Органеллы эукариотической клетки суспензированы в цитозоле - клеточном матриксе. Наиболее характерной чертой эукариотических клеток по сравнению с прокариотическими, кроме присутствия ядра, является наличие внутриклеточных органелл и пузырьков, ограниченных мембранами.
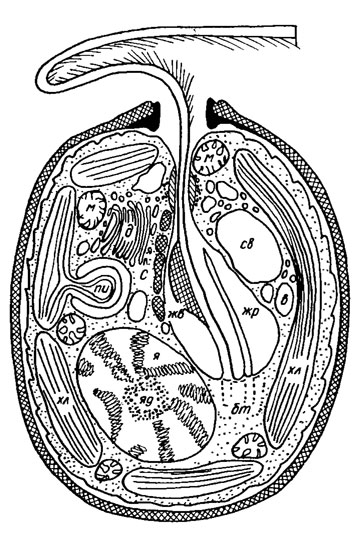
Рис. 1.31. Строение клетки Trachelomonas Ehr.: м - митохондрия; св - сократительная вакуоль; в - вакуоль; жр - жгутиковый резервуар; жв - жгутиковое вздутие; хл - хлоропласт; бт - базальное тело; яд - ядрышко; я - ядро; д - диктиосома; С - стигма
Мембранная система. Мембраны - живые компоненты цитоплазмы. Наружная мембрана цитоплазмы - плазмалемма, или цитоплазматическая мембрана, и внутренние мембраны имеют общие структурные особенности: это тончайшие (4-10 нм) довольно плотные пленки, состоящие в основном из молекул липидов (фосфолипидов) и белков, удерживаемых вместе с помощью нековалентных взаимодействий.
Мембранные фосфолипиды представляют собой амфипатические молекулы, т. е. молекулы, имеющие гидрофильный полярный и гидрофобный неполярный концы. В окружении молекул воды амфипатические молекулы обычно агрегируют. При этом гидрофобные хвосты обращены внутрь, а гидрофильные головки обращены наружу и контактируют с молекулами воды. Так как большинство фосфолипидов в водной среде самопроизвольно образуют двойные слои (бислои), то формирование липидной основы биологических мембран, по существу, процесс самосборки. Липидный бислой, создающий относительно непроницаемый барьер для большинства водорастворимых молекул, составляет структурную основу мембраны.
Молекулы белков расположены на внутренней и внешней поверхностях бислоя, а также погружены в этот слой. Проходя через бислой насквозь, они образуют в мембране гидрофильные поры. Специфические функции мембран осуществляют главным образом белки, большинство которых представлено ферментами. Белки также осуществляют транспорт через мембрану различных веществ и могут выступать в качестве специфических рецепторов.
Характерной особенностью всех мембран является их замкнутость, непрерывность. Концы мембран никогда не бывают открытыми. Большинство мембранных липидов и белков способны перемещаться в плоскости мембраны, что обусловливает ее подвижность, текучесть.
Мембраны асимметричны - оба их слоя неодинаковы по липидному и белковому составу (например, гликолипиды и гликопротеиды располагаются исключительно на той стороне мембраны, которая не контактирует с цитоплазмой).
Одно из основных свойств биологических мембран - их избирательная проницаемость (полупроницаемость), что во многом определяет специфический химический состав цитоплазмы и ее органелл. Плазмалемма, обладая избирательной проницаемостью, регулирует обмен веществ клетки с окружающей средой, выполняет синтетические функции, воспринимает раздражения.
У одноклеточных водорослей плазмалемма часто образует многочисленные складки (рис. 1.32), что увеличивает ее поверхность и может быть связано с усиленным транспортом веществ.
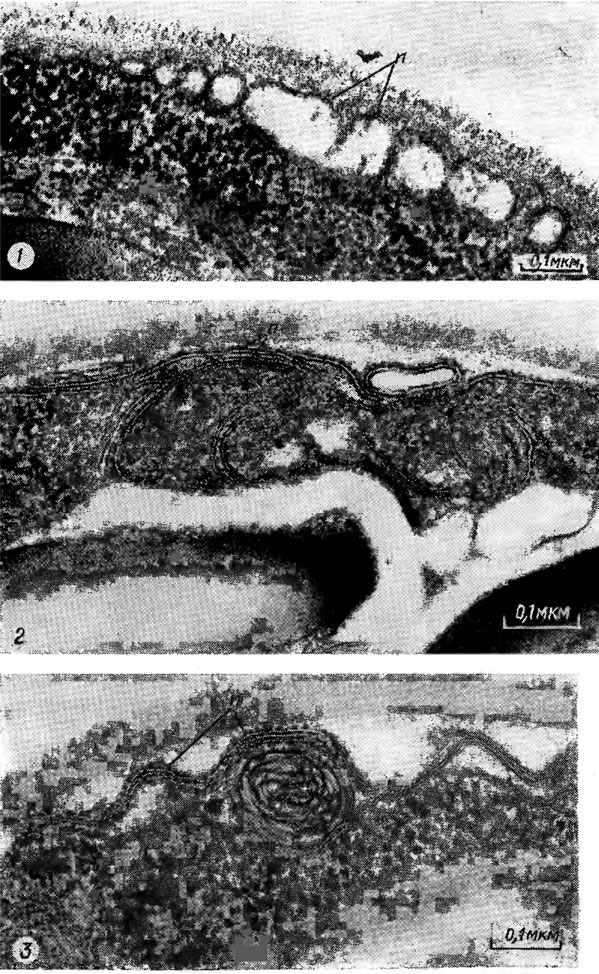
Рис. 1.32. Фрагменты клеток Chlorella vulgaris Beijer. со складками плазмалеммы (n)
Внутренние мембраны, отличались от плазмалеммы и друг от друга по физическим и химическим свойствам, разделяют клетку на изолированные отсеки - клеточные органеллы с уникальным набором ферментов, выполняющие специализированные функции. Это позволяет клетке осуществлять одновременно много несовместимых друг с другом химических реакций. Отдельные ферменты и их комплексы определенным образом локализованы в мембранах, что обеспечивает последовательность протекания химических реакций, лежащих в основе жизнедеятельности клетки.
Эндоплазматический ретикулюм представляет собой ограниченную мембраной систему субмикроскопических каналов, пронизывающих цитозоль. Мембрана эндоплазматического ретикулюма образует одну непрерывную поверхность. Замкнутое пространство, ограниченное мембраной, называют полостью эндоплазматического ретикулюма, или цистернальным пространством. От цитозоля эта полость отделена одиночной мембраной. С наружной мембраной ядерной оболочки мембрана эндоплазматического ретикулюма составляет единое целое, вследствие чего полость эндоплазматического ретикулюма и внутриядерное пространство разделены только внутренней мембраной ядерной оболочки.
Эндоплазматический ретикулюм играет центральную роль в процессе биосинтеза и транспорта ферментов, конституционных белков, липидов и углеводов, необходимых для сборки других клеточных органелл. Через эндоплазматический ретикулюм осуществляется взаимодействие органелл, он выполняет также производительную функцию, давая начало вакуолям и микротелам. У многих видов водорослей каналы эндоплазматического ретикулюма образуют обкладку вокруг хлоропластов.
Описаны две функционально различающиеся области мембраны эндоплазматического ретикулюма - гранулярная (шероховатая), к внешней стороне которой прикреплены рибосомы, и агранулярная (гладкая), лишенная рибосом. Гранулярный эндоплазматический ретикулюм на срезах клетки выявляется, как правило, в виде профилей шириной 20-50 нм, состоящих из двух более или менее параллельных замкнутых мембран с прикрепленными на внешней стороне рибосомами. Агранулярный эндоплазматический ретикулюм, состоящий из сети тонких трубочек или пузырьков, обычно развит слабее, чем гранулярный. Простые или разветвленные каналы эндоплазматического ретикулюма распределены в гиалоплазме неравномерно: они могут быть сосредоточены вокруг ядра, диктиосом, стигмы, а также в периферической части клетки вдоль плазмалеммы.
Аппарат Гольджи в клетках водорослей, как и в других растительных клетках, состоит из отдельных диктиосом - стопок из 2-7 и более (до 30) плоских круглых мешочков, или цистерн, диаметром около 1 мкм и толщиной 20-40 нм, ограниченных одинарной мембраной, и пузырьков Гольджи, которые отчленяются от краев цистерн и различаются по размерам и характеру содержимого. Диктиосома структурно и биохимически поляризована на формирующуюся, или цис-сторону (полюс регенерации), и зрелую, или транс-сторону (секреторный полюс). У ряда видов водорослей в клетке содержится только одна крупная диктиосома, часто характерной формы. Например, у Chrysochromulina apheles (Haptophyceae) она сходна с книгой, страницы которой в части, обращенной к одному из двух хлоропластов, сложены, а к другому - раскрыты. В районе более тесного расположения цистерн часто наблюдается вакуоль (рис. 1.33). У других видов водорослей число диктиосом в клетке может достигать нескольких десятков. Располагаются диктиосомы преимущественно в районе ядра, в подвижных клетках - часто у основания жгутиков, во время деления клетки - в местах образования перегородки. Если диктиосома в клетке одна, то ее удвоение предшествует цитокинезу. Аппарат Гольджи полифункционален. Он является центром концентрации, преобразования, сортировки, упаковки, транспорта и секреции полисахаридов и белков для роста оболочки и других покровов клетки, а также участвует совместно с другими органеллами в сборке и специализации мембран, образовании вакуолей и лизосом (в животной клетке), секретирует материал для сократительной вакуоли, выполняющей роль осморегулятора.
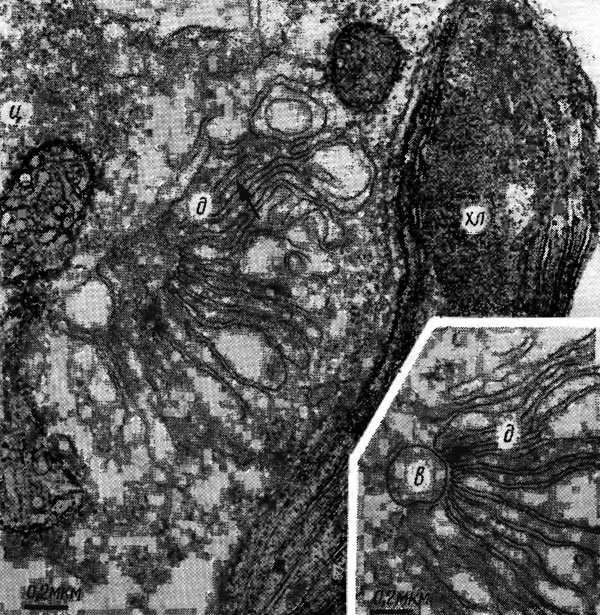
Рис. 1.33. Фрагменты клеток Chrysochromulina apheles: хл - хлоропласт; д - диктиосома; в - вакуоль, ц - цитоплазма (стрелкой указана одна из цистерн)
Ядерный аппарат. Клетки большинства водорослей одноядерны, но в различных таксонах имеются виды, клетки которых содержат 2-3 ядра и более, вплоть до нескольких десятков и сотен. У большинства водорослей ядра очень мелкие, около 1 мкм в диаметре; у ряда видов они значительно крупнее - в среднем до 4 мкм, а у отдельных видов могут достигать 45 мкм (Bryopsis из Siphonales). У одноклеточных водорослей ядро располагается ацентрично, у многоклеточных - чаще всего в центре. Общий план строения, функции ядра во всех отделах водорослей (за исключением динофитовых) не отличаются от таковых других эукариотических организмов.
Ядро - органелла клетки, в которой осуществляются важнейшие процессы, обусловливающие ее жизнедеятельность. В ядре содержится и воспроизводится наследственная информация, определяющая признаки данной клетки и всего организма в целом, осуществляются основные процессы, связанные с реализацией наследственной информации. Ядро является также центром управления обменом веществ клетки, контролирующим деятельность всех других органелл. У динофитовых - уникальных эукариот, в хромосомах отсутствуют гистоновые белки, характерные для хромосом всех других эукариот. В интерфазе хромосомы динофитовых остаются спирализованными; они лишены центромер. Ядрышко имеется или отсутствует (в зависимости от вида), может делиться во время митоза или исчезать и формироваться de novo в телофазе (как у большинства эукариот). Допускается, что структура и поведение хромосом у динофитовых водорослей примитивнее, чем у других эукариот. На этом основании некоторые исследователи даже выделяют Dinophyta в новый тип Mesokaryota (мезокариотические организмы), промежуточный между Prokaryota и Eukaryota.
Энергетический аппарат. Обеспечение энергетических потребностей эукариотических растительных клеток происходит в результате функционирования митохондрий и хлоропластов - полуавтономных самовоспроизводящихся органелл, которые обладают собственным белоксинтезирующим аппаратом, для которого матрицей служит митохондриальная или хлоропластная ДНК, но основная часть белков митохондрий и хлоропластов синтезируется в цитоплазме под контролем ядра клетки.
Митохондрии окружены оболочкой, состоящей из двух мембран, различающихся по строению и химическому составу. Внутренняя мембрана образует во внутреннем пространстве митохондрии многочисленные выросты в виде пластин, реже трубок различной длины, называемых митохондриальными кристами. Поверхность крист, обращенная к матриксу, усеяна частичками, каждая из которых состоит из головки диаметром 9 нм и ножки, связанной с мембраной. Частички являются частью АТФ-синтетазного комплекса и содержат АТФазу. В гомогенном или тонкогранулярном матриксе митохондрии с высоким содержанием белка, заполняющем пространство между кристами, находятся более мелкие по сравнению с цитоплазматическими рибосомы и отдельные кальциевые гранулы, в которых кальций находится в основном в виде фосфата. В светлых зонах матрикса расположены фибриллы митохондриальной ДНК.
Митохондрии являются центрами внутриклеточного окисления и содержат ферменты цикла трикарбоновых кислот, дыхательной цепи переноса электронов, окислительного фосфорилирования. Молекулы богатого энергией аденозинтрифосфата выходят из митохондрий и используются в разнообразных реакциях клеточного метаболизма. Так как окислительное фосфорилирование и большинство реакций окисления питательных веществ протекают на внутренней мембране митохондрий и ее производных - кристах, степень развития последних (размеры и количество) является показателем активности митохондрий: длинные, густо расположенные кристы характерны для митохондрий, находящихся в состоянии высокой функциональной активности, короткие и редкие - для малоактивных органелл.
Размеры, форма и количество митохондрий значительно варьируют в клетках различных видов водорослей. Мелкие митохондрии округлой или овальной формы имеют размеры 0,3-1 мкм. Нитевидные митохондрии могут достигать в длину 2 мкм и более при ширине 0,4 мкм. Длина гигантских митохондрий в гаметах Bryopsis в 2 раза превышает длину клетки. В отличие от высших растений клетки некоторых видов водорослей, например, Chlorella fusca Shihira et Krauss, содержат только одну сильно разветвленную митохондрию, выросты которой простираются по всей цитоплазме, формируя т. н. митохондриальный ретикулюм.
Митохондрии распределены по всему объему цитоплазмы. При этом отмечается скопление митохондрий в зонах клеток с повышенной функциональной активностью, требующих дополнительных затрат энергии. Клеточная популяция митохондрий гетерогенна; ее состав отражает возрастные и функциональные особенности клетки.
Несмотря на чрезвычайное разнообразие морфологических особенностей хлоропластов у водорослей различных отделов, их структурной фотосинтезирующей единицей, как у прокариотических организмов, так и высших растений, является тилакоид - плоский мешочек, окруженный мембраной, который в поперечном сечении имеет вид пары параллельных мембран, более или менее плотно прилегающих друг к другу и соединенных на концах. В хлоропластах осуществляется фотосинтез (ассимиляция углекислого газа воздуха), образование органических веществ из неорганических за счет энергии света.
Фотосинтетический аппарат. Пигментные системы и переносчики электронов встроены в тилакоидные мембраны. Хлорофиллы, каротиноиды и фикобилины - пигменты, участвующие в процессах световой фазы фотосинтеза, входят в состав пигментных систем в виде пигмент-белковых комплексов. У всех фотосинтезирующих растений, в том числе и у водорослей, в преобладающем количестве в тилакоидах содержится хлорофилл а. Хлорофилл b среди водорослей обнаружен только у зеленых и эвгленовых. Хлорофиллы с1 и с2 встречаются у бурых, золотистых диатомовых водорослей, у криптофитовых - только хлорофилл с2. Зеленые водоросли содержат каротиноиды, типичные для высших растений. Наличие больших количеств фукоксантина у видов бурых водорослей Определяет их бурую окраску. Для красных и криптофитовых водорослей характерны фикобилины - красные фикоэритрины и синие фикоцианины. Фикоби ли протеины агрегируют друг с другом, образуя особые гранулы, называемые фикобилисомами, которые упорядоченно расположены на поверхности тилакоидных мембран. Фикобилины выполняют роль дополнительной светособирающей антенны. Темновые реакции - восстановление углекислого газа водородом воды до углеводов (фиксация углекислого газа) - протекают в строме (см. ниже) хлоропластов. В хлоропластах, как и в митохондриях, происходит синтез аденозинтрифосфата из аденозиндифосфата. Так как источником энергии для этого процесса в хлоропластах служит солнечный свет, то его называют фотофосфорилированием.
Строма хлоропластов - их белковая основа, в которую погружены тилакоиды - обычно довольно электронно-плотная. В ней обнаруживаются пластоглобулы, состоящие из особых липидов - пластохинонов, рибосомы, в более светлых зонах - фибриллы ДНК. В строме хлоропластов также может откладываться крахмал в виде зерен и белок в виде кристаллов; иногда наблюдаются структуры, напоминающие микротрубочки.
Нередко внутренняя мембрана оболочки хлоропласта образует выросты в строму в виде сети трубок по периферии органеллы, называемой периферическим ретикулюмом. Хлоропласты водорослей многих отделов, кроме красных, криптофитовых, харовых и зеленых, окружены каналом эндоплазматического ретикулюма.
Морфологическое разнообразие хлоропластов водорослей прежде всего создается за счет расположения тилакоидов, которое весьма специфично в разных отделах. У большинства эукариотических водорослей, за исключением красных, тилакоиды собраны в пучки, состоящие из 2 тилакоидов (криптофитовые, у которых фикобилисомы как структурные единицы отсутствуют), 3 (динофитовые, золотистые, желтозеленые, бурые, диатомовые), 2-6 и более (зеленые и харовые), или образуют граноподобные скопления из 3 тилакоидов (эвгленовые). У красных водорослей фотосинтетическая система представлена одиночными длинными тилакоидами, несущими на своей поверхности фикобилисомы (рис. 1.34). На основе представлений об едином исходном типе организации тилакоидов рассмотрены возможные направления филогенетического становления различных типов организации хлоропластов (рис. 1.35).
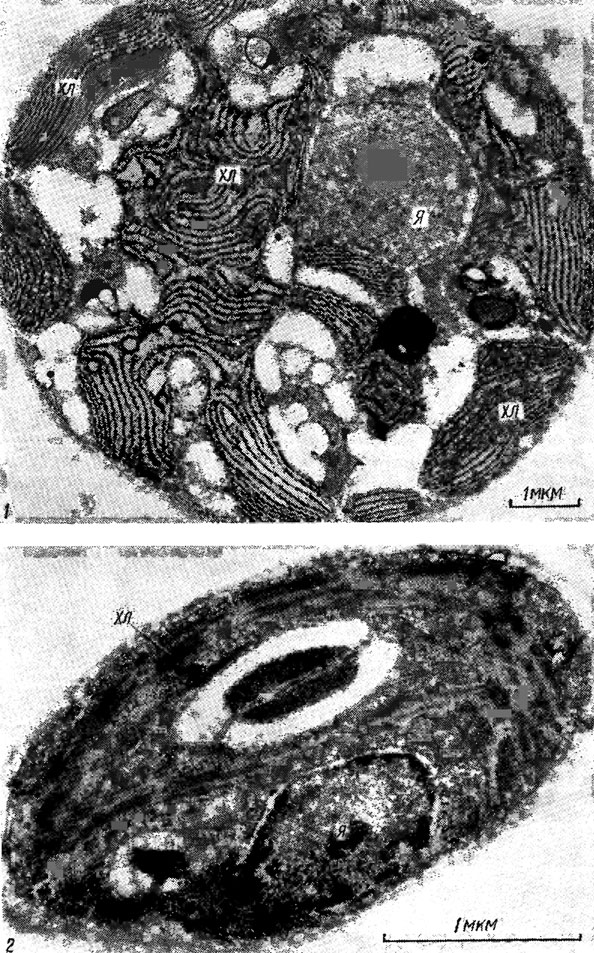
Рис. 1.34. Электронная микрофотография Flintiella sanguinaria Ott. (1) и Chlorella vulgaris Beijer. (2): хл - хлоропласт; я - ядро
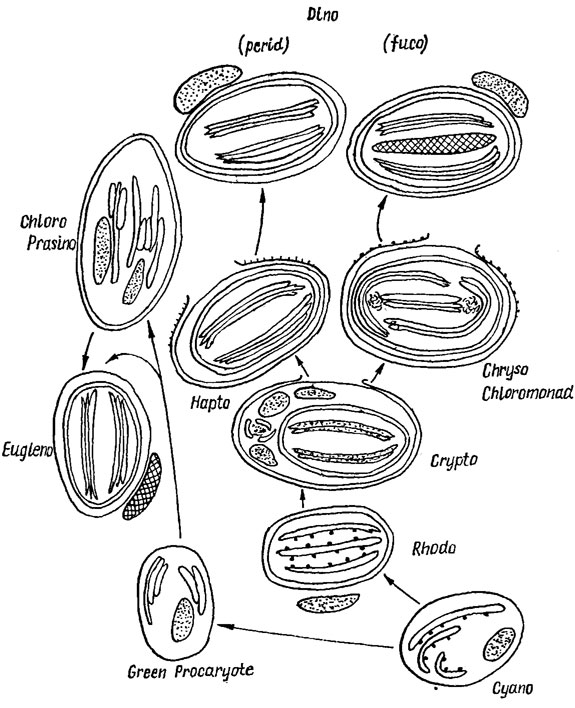
Рис. 1.35. Схема возможных филогенетических взаимоотношений различных типов хлоропластов
У многих видов водорослей, особенно одноклеточных, в клетках содержится по одному очень крупному хлоропласту; относительно часто встречаются виды с двумя хлоропластами чашевидной, поясковидной, звездчатой, спиралевидной формы. При увеличении числа хлоропластов у ряда неклеточных и многоклеточных водорослей они в основном приобретают линзовидную форму. Следует отметить, что развитие фотосинтетической системы хлоропластов меняется под воздействием различных внешних факторов - интенсивности и качества света, температуры, состава питательной среды и т. д.
Дополнительное своеобразие в структурную организацию хлоропластов водорослей вносит наличие пиреноида - полуавтономной системы, функционально и пространственно тесно связанной с хлоропластом. Пиреноид представляет собой образование белковой природы (в основном состоит из фермента рибулезодифосфаткарбоксилазы), размером 3-12 мкм, и обычно окружен крахмальными зернами - амилогенной обкладкой, в образовании которой может принимать участие ламинарии (бурые водоросли), пирамилон (эвгленовые), липиды (золотистые). Гранулярный или тонкофибриллярный матрикс пиреноида обычно плотнее стромы хлоропласта, часто перфорирован 1-2 тилакоидами. У подавляющего большинства водорослей пиреноид находится внутри хлоропласта, у многих желтозеленых, динофитовых и криптофитовых пиреноиды располагаются непосредственно под оболочкой хлоропласта, в ряде случаев несколько выступая наружу. У некоторых видов эвгленовых, динофитовых, криптофитовых, золотистых, желтозеленых, диатомовых и бурых пиреноиды лежат за пределами хлоропласта, но тесно с ним связаны. Количество их варьирует от 1 до 10 и более.
Согласно современным представлениям, функция пиреноида заключается в концентрации рибулезодифосфаткарбоксилазы с целью распределения ее между дочерними клетками и транспорта в места активного функционирования в хлоропласте. Наличие, размер пиреноида и его амилогенной обкладки во многом зависят от окружающих условий (интенсивности освещения, температуры, состава питательной среды), а также определяются эндогенными факторами.
У подвижных форм водорослей в хлоропласте или (значительно реже) вне его, но всегда в функциональной связи с ним, располагается специфическая фоторецепторная органелла этих форм водорослей - стигма, или глазок, имеющая в прижизненном состоянии сферическую, палочковидную, линзовидную или трапециевидную форму. Строгая пространственная ориентация стигмы на жгутиковый аппарат связана с его функцией регуляции направленного движения клеток путем улавливания световых импульсов, их трансформации и передачи жгутиковому аппарату*. В клетках, длительное время находящихся в темноте, стигма исчезает, но при перенесении культуры на свет появляется снова. Структурную основу стигмы составляют тесно прилегающие друг к другу пигментсодержащие глобулы (каротиноиды), 75-660 нм в диаметре; иногда (у эвгленовых) глобулы ограничены мембраной. По способу упорядоченности глобул, положению стигмы, а также наличию своеобразных дополнительных структур - пластинчатых, линзовидных тел и ритеноида, усложняющих ее организацию, выделено 5 основных типов стигм (рис. 1.36). Пластинчатое тело стигмы (некоторые виды динофитовых), длиной около 3 и шириной 1 мкм, состоит из нескольких (до 50) параллельно расположенных уплощенных мешочков, сообщающихся между собой; крайние из них непосредственно переходят в канал эндоплазматического ретикулюма. Линзовидное тело, встречающееся в стигме также ряда видов динофитовых, занимает почти 2/3 стигмы и состоит из группы уплощенных пузырьков, из которых внутренние заполнены зернистым содержимым. В основании линзы находится ретиноид - чашеподобное образование, содержавшее зоны определенным образом ориентированных фибрилл и зернистого вещества, а также пигментные глобулы около 300 нм в диаметре, расположенные на самом дне в один ряд. У многих видов золотистых, бурых, желтозеленых, эвгленовых в основании одного из жгутиков образуется вздутие, или парабазальное тело, которое размещается точно против вогнутости стигмы, оставаясь отделенным от нее клеточным покровом и оболочкой хлоропласта. Типы структурной организации стигм постоянны в каждом отделе водорослей.
* (Существует и другая точка зрения о функции стигмы (см. раздел 1.3.1).)
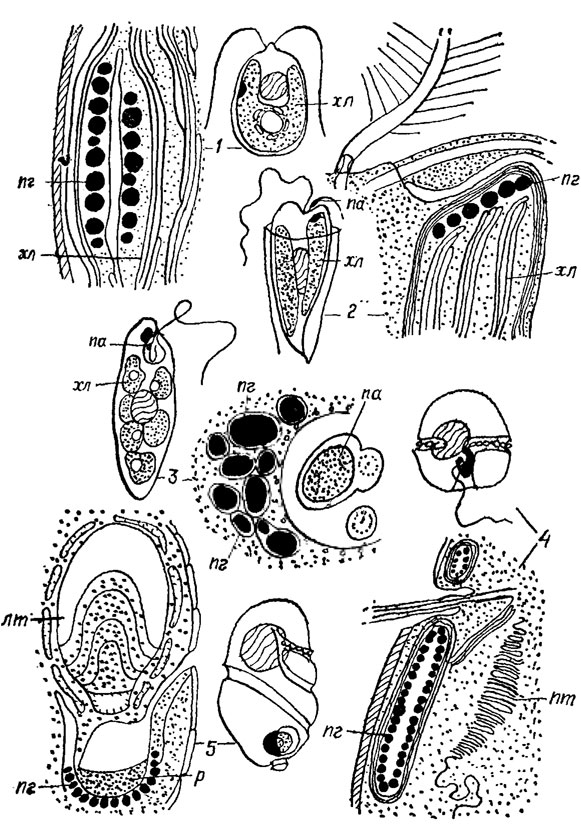
Рис. 1.36. Основные типы стигм: 1 - стигма I типа (Chlamydomonas); 2 - стигма II типа (Dinobryon); 3 - стигма III типа (Euglena); 4 - стигма IV типа (Glenodinium). 5 - стигма V типа (Nematodium); пг - пигментные глобулы; хл - хлоропласт; па - парабазальное тело; лт - линзовидное тело; пт - пластинчатое тело; р - ретниоид
Вакуоли. Вакуоли являются производными эндоплазматического ретикулюма. Они образуются в основном путем локального расширения гранулярных цистерн последнего, ограничены одинарной избирательно проницаемой мембраной - тонопластом - и выполняют в растительных клетках разнообразные функции: поддержание тургора, накопление запасных веществ и отходов жизнедеятельности клетки, осуществление локального автолиза. Для подвижных клеток, а иногда и для неподвижных одноклеточных, характерно наличие сократительных вакуолей, располагающихся обычно у жгутиковых форм в основании жгутиков и выполняющих роль осморегулятора. У динофитовых эту функцию выполняют особые лишенные способности сокращаться вакуоли - пузулы.
Вакуолярная система водорослей может быть представлена вакуолями различных размеров, причем у ряда видов одноклеточных водорослей крупная вакуоль обычно занимает определенное место в клетке. У многоклеточных, особенно у водорослей с паренхиматозным типом структуры (см. раздел 1.3), в процессе дифференцировки клеток меристемальных зон в различные в функциональном отношении клетки - покровные и проводящие - увеличивается объем и количество вакуолей. Дифференцированные клетки содержат крупную центральную вакуоль. Степень развития вакуолярной системы варьирует в клетках водорослей с гетеротрихальной структурой слоевища (например, клетки горизонтальной части слоевища Fritschiella tuberosa Iyengar значительно более вакуолизированы по сравнению с клетками вертикальной части; ризоиды также сильно вакуолизированы).
Другие органеллы. Органеллы, идентифицируемые как лизосомы животных клеток, у автотрофных организмов отсутствуют. В клетках водорослей описаны пероксисомы, характерные для эукариотических организмов. Эти мелкие органеллы, 0,15-0,25 мкм, окружены мембраной и образуются путем отпочкования от цистерн эндоплазматического ретикулюма, но часто остаются соединенными с ними. Пероксисомы, подобно митохондриям, утилизируют кислород в клетке путем осуществления окислительных реакций. Образующаяся при этом перекись водорода используется или разрушается с помощью содержащейся в этих органеллах каталазы.
Цитозоль, в который погружены органеллы, пронизан густой сетью белковых филаментов, составляющих клеточный скелет и обеспечивающих пространственно-временную организацию клетки и движение цитоплазмы. Два их наиболее важных компонента - тубулиновые микротрубочки и актиновые филаменты. Последние состоят из двух цепей глобулярных молекул диаметром около 4 нм, образующих двойную спираль. В настоящее время уделяется большое внимание выяснению роли цитоплазматических микротрубочек в определении формы зооспор, агрегации клеток в колонии - ценобии, а также изучению организации и химического состава микрофиламентов, их функциям во вращательном движении цитоплазмы.
Включения. С локальной концентрацией в цитоплазме или различных органеллах определенных продуктов обмена веществ связано образование включений, которые представляют собой соединения, временно выведенные из клеточного метаболизма, или конечные продукты обмена (например, в клетках бурых водорослей содержатся значительные количества полифенолов). Ирисовые тела, состоящие в основном из белков - глюцидов, описаны в клетках бурых, красных и некоторых зеленых водорослей. Для многих видов водорослей характерно наличие в цитоплазме электронно-плотных сферических структур, содержащих полифосфаты. Запасные липиды обнаруживаются в клетках водорослей в виде капель различных размеров, количество которых связано с функциональным состоянием и возрастом клеток.
Деление клетки
Митоз - универсальная форма деления ядра у всех эукариотических организмов, обеспечивающая одинаковое распределение наследственного материала между двумя дочерними клетками, постоянство числа, формы и размеров структурных единиц наследственности - хромосом. У представителей разных таксонов водорослей митоз обнаруживает вариации и отличается от классического митоза клеток высших растений, основными чертами которого являются спирализация хромосом в профазе, исчезновение ядрышка и ядерной оболочки в поздней профазе или метафазе, формирование митотического веретена из микротрубочек от центромер хромосом к полюсам веретена и от полюса к полюсу, отсутствие центриолей. Наличие центриолей* и сохранение в процессе митоза всей или только части ядерной оболочки описаны у многих видов водорослей, представленных как подвижными, так и неподвижными формами. На этом основании у водорослей выделяют закрытый митоз (ядерная оболочка остается непрерывной), полузакрытый митоз (в ядерной оболочке образуются полярные отверстия либо ядерная оболочка фрагментаруется, но не исчезает) и открытый митоз (ядерная оболочка дезинтегрируется, как у высших растений). Нередко наблюдается формирование перинуклеарной оболочки за счет окружения делящегося ядра каналами эндоплазматического ретикулюма. При митозе варьирует также форма веретена (прямая, согнутая, узкая, широкая и т. д.) и форма полюсов веретена (острая, тупая, округлая), а также продолжительность существования межзонального веретена.
* (Центр ноли делятся в ранней профазе митоза и сохраняются во всех фазах клеточного цикла или же появляются только во время митоза, нередко перед образованием подвижных клеток. В вегетативных клетках ряда многоклеточных водорослей центриол и редуцированы и наблюдаются нерегулярно. В отличие от животной клетки центриоли клеток водорослей могут занимать различное положение в делящихся клетках, располагаясь на полюсах митотического аппарата и структурно участвуя в его формировании (веретено с центр нолями на полюсах) или занимают по отношению к веретену латеральное положение.)
Закрытый митоз присущ, в частности, монадным формам (например, Реdimonas minor Korsch.) и нитчатым (Oedogonium cardiacum (Hass.) Wittr.; рис. 1.37, 1). Причем у видов рода Oedogonium отмечено образование выростов ядерной оболочки в цитоплазму, иногда с отверстиями на концах. Закрытый митоз характерен, в частности, для сифоновой водоросли Acetabularia mediterranea Lamx. (веретено формируется внутри ядра). Полузакрытый митоз происходит у Chlamydomonas reinhardtii Dang., Ulva mutabilis Fθyn., Flintiella sanguinaria Ott (рис. 1.37, 2). Открытый митоз наблюдается, например, у видов родов Closterium Nitzsch., Klebsormidium Silva, Zygnema G. Ag., Coleochaete, Draparnaldia Bory. Пик митозов приходится на темновой период суток. В многоядерных клетках деление ядер может проходить синхронно, асинхронно, волнообразно.
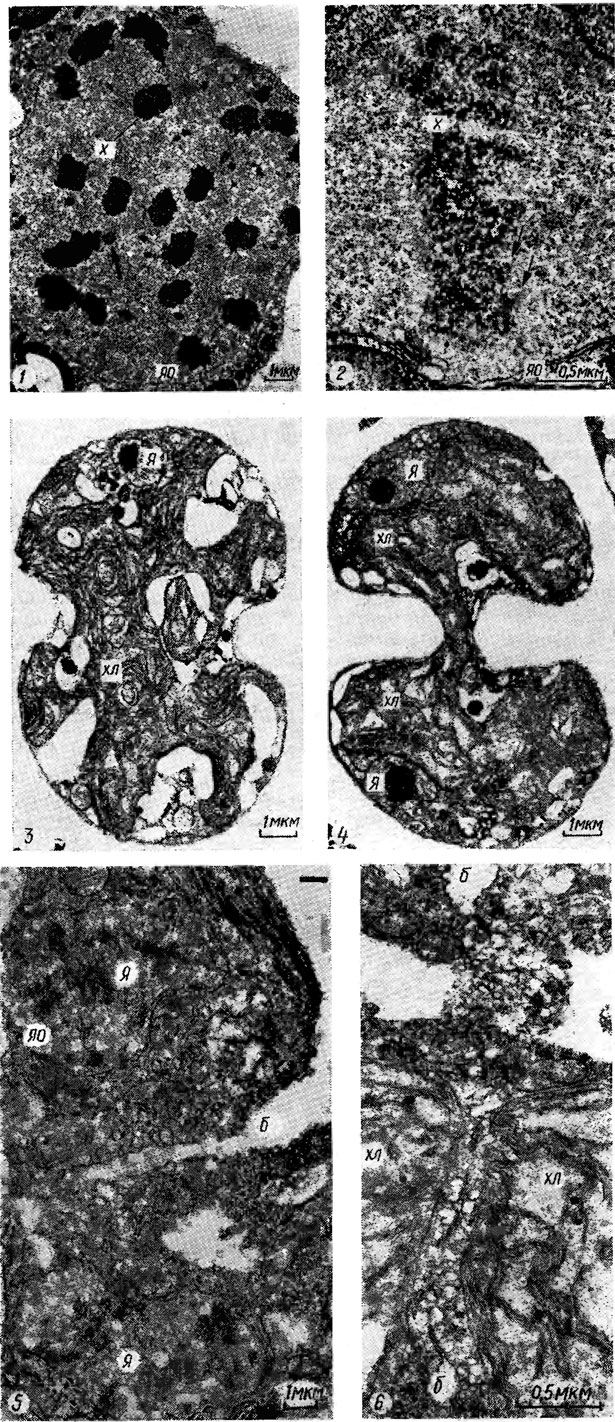
Рис. 1.37. Электронная микрофотография фрагментов клеток в метафазе митоза (1, 2) и в цитокинезе (3-6): 1 - Oedogonium cardiacum (Hass.) Wittr,; 2-4 - Flintiella sanguinaria Ott.; 5, 6 - Ulothrix mucosa Turp.; х - хромосома; яо - ядерная оболочка; я - ядро; хл - хлоропласт; б - борозда; видны кинетохоры (стрелки)
Цитокинез - деление цитоплазмы путем образования борозды деления или клеточной пластинки в экваториальной плоскости клетки, между дочерними ядрами. Цитокинез у водорослей осуществляется путем центростремительного роста новой клеточной оболочки. Борозда деления начинается от внутреннего слоя клеточной оболочки и охватывает кольцом всю клетку (см. рис. 1.37, 3-6). Существуют вариации цитокинеза у видов водорослей, относящихся к различным порядкам. Они могут заключаться, в частности, в наличии или отсутствии фрагмопласта или фикопласта, либо обоих структур вместе, в степени ассоциации фикопласта с центриолям'и, времени исчезновения веретена деления клетки, наличии или отсутствии клеточной пластинки фрагмопласта (фрагмопласт представляет собой микротрубочки, ориентированные параллельно оси веретена деления, и клеточную пластинку, образующуюся путем слияния пузырьков Гольджи, ассоциированных с микротрубочками в экваториальной плоскости клетки, и растущую центробежно; фикопласт - микротрубочки, расположенные перпендикулярно оси веретена и параллельно плоскости деления клетки).
Так, у видов рода Spirogyra Link цитокинез, начинающийся в ранней анафазе, осуществляется как путем одновременного заложения борозды деления, растущей центростремительно, так и путем образования фрагмопласта. В цитоплазме Spirogyra, окружающей края борозды деления, наблюдаются пузырьки различных размеров (очевидно, производные аппарата Гольджи). Иногда содержимое пузырьков включается в борозду. Рост борозды непосредственно не зависит от присутствия митотического аппарата, но положение ядра, возможно, определяет место будущей борозды.
При цитокинезе Kirchniella lunar is (Kirchn.) Moebius, в результате которого образуются 4 автоспоры, после первого митоза между двумя дочерними ядрами закладывается перегородка, которая может частично или полностью исчезать во время второго митоза, в течение которого два дочерних ядра делятся синхронно. Центриоли в клетке К. lunaris в интерфазе не найдены, они формируются в премитотической стадии и присутствуют до окончания цитокинеза после второго митоза. После первого митоза в цитоплазме возникает сложная система микротрубочек, ориентированных в различных направлениях. После второго митоза микротрубочки веретена исчезают в телофазе и снова появляются между ядрами перпендикулярно к оси веретена. Если борозда, закладывающаяся после первого митоза, исчезает во время второго, то после второго митоза она восстанавливается на том же месте. Появление микротрубочек вдоль первой борозды после второго митоза отмечено и в тех случаях, когда она не исчезает во время второго митоза.
Для видов рода Klebsormtdium Silva (Ulotrichales) характерно удлинение веретена деления клетки в анафазе, отсутствие как фикопласта, так и фрагмопласта, наличие борозды деления; для видов рода Microsроra Thur. (Microsporeles) - раннее исчезновение веретена деления, наличие борозды деления, а также фикопласта, ассоциированного с цен триолями; для видов родов Ulotrix Kütz., Uronema Lagerh., Stigeoclonium Kütz., Chaetophora Schrank, Draparnaldia, Schizomeris Kütz. (Chaetophorales) - раннее исчезновение веретена деления, образование клеточной пластинки и фрагмопласта, наличие фикопласта, не ассоциированного с центриолями*.
* (Роль центриолей в процессе деления вегетативных клеток неподвижных одноклеточных, нитчатых и многоклеточных водорослей остается во многом не выясненной. Наиболее распространена точка зрения, что у неподвижных форм центриоли являются нефункционирующими реликтами. В то же время существует мнение, что при цитокинезе с образованием борозд деления центриоли участвуют в определении плоскости деления.)
Особенности ультраструктурной организации клеток водорослей используют как таксономический признак при критическом пересмотре систематического положения многих таксонов и при интерпретации филогении и эволюции водорослей. Ценную информацию о возникновении и эволюции типов митоза и цитокинеза в различных группах растительного и животного мира, эволюции гамет в процессе перехода от изогамии к оогамии, происхождении эукариотической клетки дают сравнительно-цитологические исследования вегетативных и репродуктивных органов водорослей.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'