1.2.1. Строение и деление клетки прокариотических водорослей (Н. В. Кондратьева)
Форма, размеры и основные типы клеток
Среди прокариотических (первичных) зеленых водорослей (Prochlorophyta) лучше всего изучены одноклеточные формы с шаровидными клетками 6-30 мкм в диаметре (см. раздел 96). К синезеленым водорослям принадлежат как одноклеточные, так и нитчатые многоклеточные (трихомальные) организмы, способные образовывать разнообразные колонии с одноклеточными или многоклеточными цианоидами (см. разделы 9а.1 - 9а.4).
Ширина клеток синезеленых водорослей чаще всего, по-видимому, находится в пределах 2-10 мкм. Но у многих видов клетки уже (иногда менее 1 мкм)* или шире (до нескольких десятков микрометров).
* (В последнее время в фотической зоне океанов и морей обнаружены в большом количестве фотосинтезирующие организмы (т. н. пикопланктон), ширина клеток которых находится в пределах 0,2-2 мкм. Некоторые из них рассматриваются как хроококкоподобные синезеленые водоросли (см., например, [625]).)
Одноклеточные индивиды и одноклеточные цианоиды (см. раздел 9а) колоний синезеленых водорослей чаще всего шаровидные или эллипсоидные (после деления полушаровидные), реже вытянутые, до цилиндрических и веретеновидных, прямые или согнутые, иногда грушевидные и др. У прикрепленных одноклеточных индивидов (а иногда и у одноклеточных цианоидов) нередко наблюдается гетерополярность клеток и образование слизистых ножек и дисков, которыми клетки прикрепляются с субстрату (см. разделы 9а.2 и 9а.3). Интеркалярные (срединные) клетки трихомов (рис. 1.2) шаровидные, овалоидные, дисковидные или цилиндрические, на поперечном срезе круглые или слегка сжатые, и только у представителей рода Gomontiella Teod. серповидные, а у Starria Lang. - трехрадиальные (рис. 1.3). Апикальные (конечные) клетки на верхушке округленные, несуженные или конические, но бывают острыми и головчатыми (рис. 1.4).
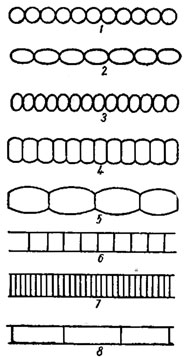
Рис. 1.2. Основные формы трихомов и клеток, образующих трихомы. Клетки: 1 - шаровидные; 2 - удлиненно-эллипсоидные (овалоидные); 3 - короткоэллипсоидные (сжато-шаровидные); 4 - коротко (сжато)-боченковидные; 5 - удлиненно-боченковидные; 6 - квадратно-цилиндрические (изодиаметрические); 7 - дисковидно-цилиндрические; В - удлиненно-цилиндрические. Трихомы: 1-3 - четковидные (монолиформные); 4, 5 - перетянутые (перешнурованные) около поперечных перегородок; 6-8 - цилиндрические, не перетянутые около поперечных перегородок
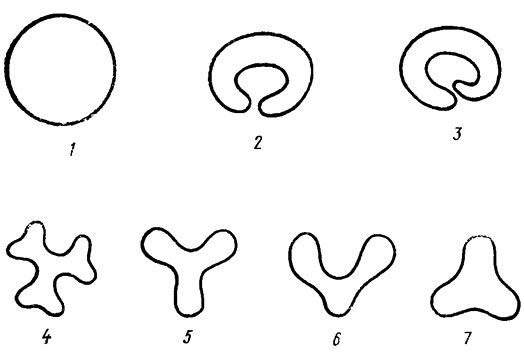
Рис. 1.3. Форма клеток гормогониевых водорослей на срезе, поперечном к главной оси нити: 1 - круговидная (у подавляющего большинства форм); 2, 3 - полукольцевидная, или серповидная (у Gomontlella marthae Claus): 4-7 - трехрадиальная (у Starria zimbabwensis Lang)
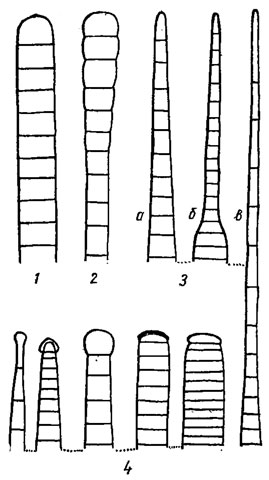
Рис. 1.4. Основные формы концов трихомов, конечных клеток трихомов и их верхушек. Концы трихомов: 1 - несуженный; 2 - булавовидный: 3 - суженный (а - постепенно суженный, волосковидный; б - внезапно суженный; в - постепенно суженный, волосковидный); 4 - головчатые (второй слева - с калиптрой). Конечные клетки: 5 - полушаровидная; 6 - почти шаровидная; 7 - дисковидная; 8 - головчатая; 9 - конические (а - удлиненно-коническая); 10 - цилиндрические (а - удлиненно-цилиндрическая). Верхушки конечных клеток: 11, 12 - округленные (12 - широкоокругленная); 13 - головчатая; 14 - притупленные; 15 - острые (заостренные); 16 - внезапно суженная
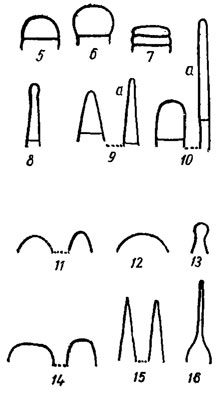
Рис. 1.4. Основные формы концов трихомов, конечных клеток трихомов и их верхушек. Концы трихомов: 1 - несуженный; 2 - булавовидный: 3 - суженный (а - постепенно суженный, волосковидный; б - внезапно суженный; в - постепенно суженный, волосковидный); 4 - головчатые (второй слева - с калиптрой). Конечные клетки: 5 - полушаровидная; 6 - почти шаровидная; 7 - дисковидная; 8 - головчатая; 9 - конические (а - удлиненно-коническая); 10 - цилиндрические (а - удлиненно-цилиндрическая). Верхушки конечных клеток: 11, 12 - округленные (12 - широкоокругленная); 13 - головчатая; 14 - притупленные; 15 - острые (заостренные); 16 - внезапно суженная
Трихомальные индивиды Cyanophyta могут состоять только из клеток, внешне сходных между собой, т. е. недифференцированных по форме и, вероятно, функции - т. н. гомоцитные трихомальные индивиды, или из клеток, явно отличающихся по форме и функциям - т. н. гетероцитные трихомальные индивиды (рис. 1.5).
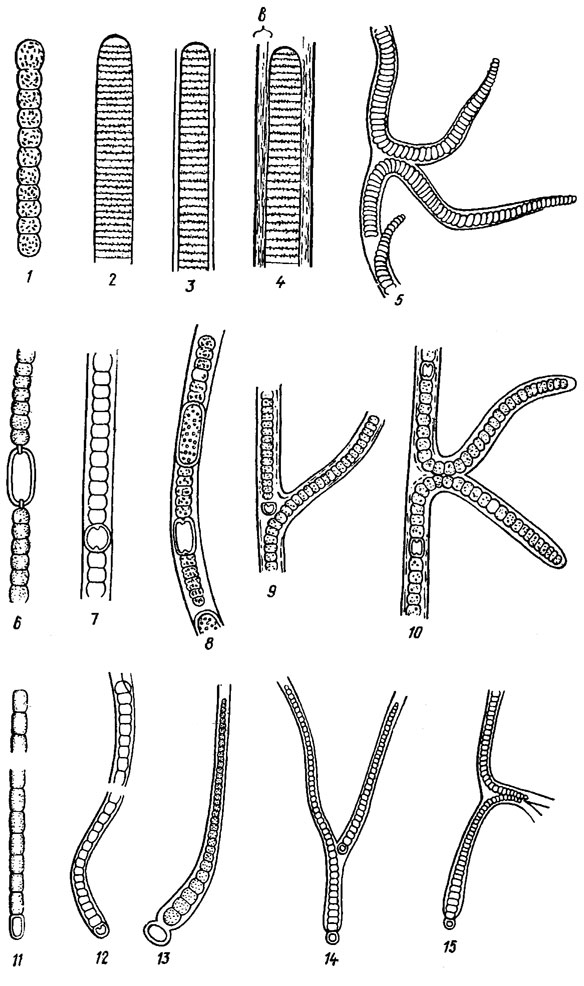
Рис. 1.5. Разные состояния (status) индивидов гормогониевых водорослей: 1-5 - гомоцитные (1, 2 - st. oscillatorioideus, 3 - st. phromidioideus, 4 - st. iyngbyoideus. 5 - st. plectonematoideus); 6-15 - гетероцитные (6 - st. anabaenoideus. 7, 8 - st. aulosiroideus, 9 - st. tolypotrichoideus. 10 - st. scytonematoideus; 11 - st. cylindrospermoideus, 12 - st. microchaetoideus; 13-15 - st. calotrichoideus); 1 - Osciliatoria lacustris (Klebs) Geitl.; 2-4 - Lyngbya aestuarii (Mert.) Liebm. (в - влагалище); 5 - Plectonema battersii Gom.; 6 - Anabaena aequalis Borge; 7 - A. variabilis Kütz. f. rotundospora Hojlerb.; 8 - Aulosira planctonica Elenk.; 9 - Tolypothrix distorta (Fl. Dan.) Kütz.; 10 - Scytonema ocellatum Lyngb.; 11 - Cylindrospermum majus Kütz.; 12 - Microchaete tener Fremy f. minor Hollerb.; 13-15 - Calothrix elenkinii Koss
В общем у синезеленых водорослей различают такие основные типы клеток: 1) клетки одноклеточных индивидов и одноклеточные цианоиды колоний; 2) клетки гомоцитных трихомальных индивидов; 3) вегетативные клетки гетероцитных трихомальных индивидов; 4) особые клетки (гетероцисты, акинеты и др.) индивидов.
Ниже, когда речь идет о строении (и размерах) клеток Cyanophyta без указания их типа, имеются в виду клетки трех первых типов. При описании особых клеток везде приведено их наименование.
Строение клетки
В связи с тем, что основная масса известных видов прокариотических водорослей принадлежит к отделу Cyanophyta (из Prochlorophyta пока описано только два вида; см. раздел 96), сведения, приведенные в настоящем разделе, касаются по сути клеток синезеленых водорослей.
Клетка синезеленых водорослей состоит из клеточных покровов и внутреннего содержимого - протопласта, к которому относятся плазмалемма и цитоплазма с различными внутрицитоплазматическими структурами (фотосинтетический аппарат, ядерный эквивалент, рибосомы и другие).
К клеточным покровам принадлежат все структуры, окружающие протопласт: клеточные оболочки, слизистые обвертки и особые трубчатые образования - т. н. влагалища*.
* (Влагалища и нередко слизистые обвертки окружают многие клетки. В последнее время эти образования совместно с клеточной стенкой (англ. cell wall) и цитоплазматической мембраной (см. ниже) предложено называть [674] клеточной обверткой (англ. cell envelope).)
Клеточная оболочка (клеточная стенка). Клетки синезеленых водорослей всегда окружены хорошо развитыми слоистыми жесткими оболочками (рис. 1.6), которые обусловливают форму клеток, выполняют опорную и защитную функцию. Толщина оболочки в среднем равна 35-50 нм, реже больше*. У акинет (рис. 1.7) и гетероцист (рис. 1.8) оболочка толще, чем у вегетативных клеток. Части клеточных оболочек, расположенные на границе двух смежных клеток одной нити, называются поперечными перегородками. Считают, что в поперечных перегородках присутствуют субмикроскопические поры и микроплазмодесмы, соединяющие протопласты соседних клеток трихома. Микроплазмодесмы следует отличать от плазмодесм, упоминаемых в диагнозах видов. Последние крупнее, подчас различимы уже при среднем увеличении светового микроскопа и, скорее всего, являются тяжами протоплазмы, подходящими к неутолщенным местам поперечных перегородок, которые (как и субмикроскопические поры) называют порами. Субмикроскопические поры обнаружены не только в поперечных, но и в продольных (см. рис. 1.6) частях оболочек клеток трихомальных индивидов. Не исключено, что они служат для выделения слизи.
* (1 мм = 106 нм.)
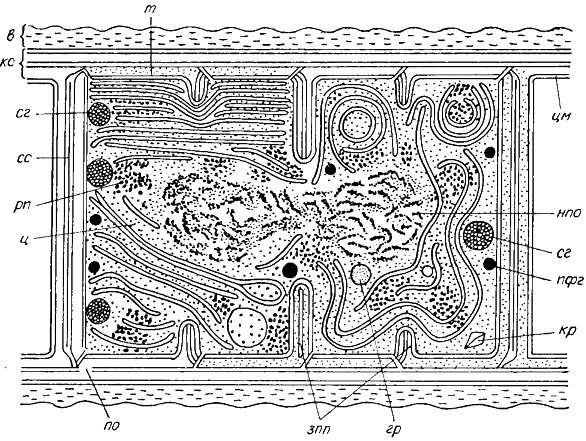
Рис. 1.6. Схематическое изображение тонкого продольного среза через клетку синезеленой водоросли: в - влагалище; гр - гранула неизвестного состава и функций; зпп - зачатки новых клеточных перегородок; кр - кристалл; кс - клеточная стенка (оболочка клетки); нпо - нуклеоплазматическая область; по - пора клеточной оболочки; пфг - полифосфатная гранула (волютин); рп - рибосомы и полисомы; сг - структурированная гранула (цианофициновое зерно); сс - срединный слой поперечной части клеточной стенки (поперечной перегородки); т - тилаконд; ц - цитоплазматический матрикс; цм - цитоплазматическая мембрана
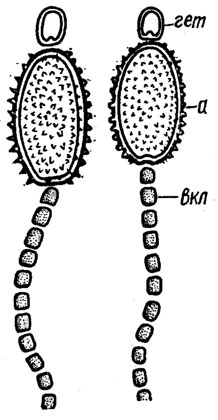
Рис. 1.7. Части нитей Cylindrospermum majus Kütz: гет - терминальная гетероциста: а - акинета; вкл - вегетативная клетка
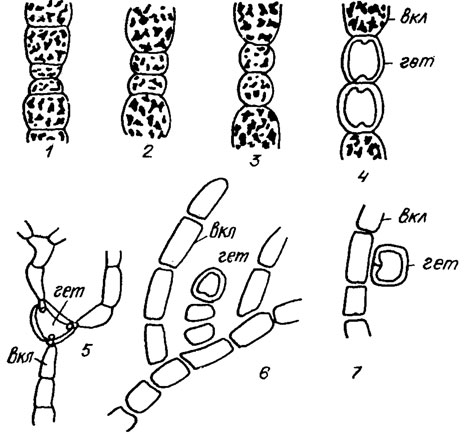
Рис. 1.8. Гетероцисты: 1-4 - схема развития гетероцист у Anabaenopsis arnoldli Aptek.: 5 - трехпоровая нитеркалярная гетероциста у Brachytrichia balani (Lloyd) Born, et Flan.; 6, 7 - гетероциста Nostochopsis lobatus Wood. (6 - терминальная гетероциста на конце малоклеточной ветки; 7 - боковая, или латеральная, гетероциста, образованная на месте одноклеточной боковой ветви); вкл - вегетативная клетка; гет - гетероциста
В оболочках гетероцист, в местах примыкания их к вегетативным клеткам, имеются поровые каналы, нередко закрытые пробойками (см. рис. 1.8). Наличие таких поровых каналов (пор) и пробочек наряду с присутствием утолщенных оболочек и обесцвеченным содержимым клетки, является характерной чертой зрелых гетероцист, отличающей их от вегетативных клеток.
При больших увеличениях светового микроскопа поверхность клеток кажется гладкой. Но на поверхности акинет нередко встречаются разнообразные украшения (см. рис. 1.7).
В клеточных оболочках синезеленых водорослей обнаружены пектины, углеводы, аминокислоты, жирные кислоты, липополисахариды и др. Однако наиболее характерной их чертой является присутствие особого гетерополимера - муреина (пептидогликана), образующего особый пептидогликановый слой, что сближает Cyanophyta с грамотринательными бактериями и принципиально отличает от эукариотических организмов.
Слизистые обвертки и влагалища. Клетки синезеленых водорослей обычно окружены слизистыми обвертками, разнообразными по своим свойствам и происхождению. В одних случаях слизь образуется в небольшом количестве, в других вокруг клетки возникают более или менее стойкие и четкие слизистые обвертки. Слизистые продукты, образованные разными клетками, могут объединяться, что нередко ведет к возникновению колониальной слизи. Внешний слой колониальной слизи иногда уплотнен, кажется двухконтурным и называется "перидермом" (рис. 1.9). Вокруг трихомов часто образуются также трубчатые структуры - т. н. влагалища (чехлы) (см. рис. 1.5; 1.6), которые бывают тонкими или толстыми, неслоистыми или слоистыми и обычно имеют фибриллярное строение.
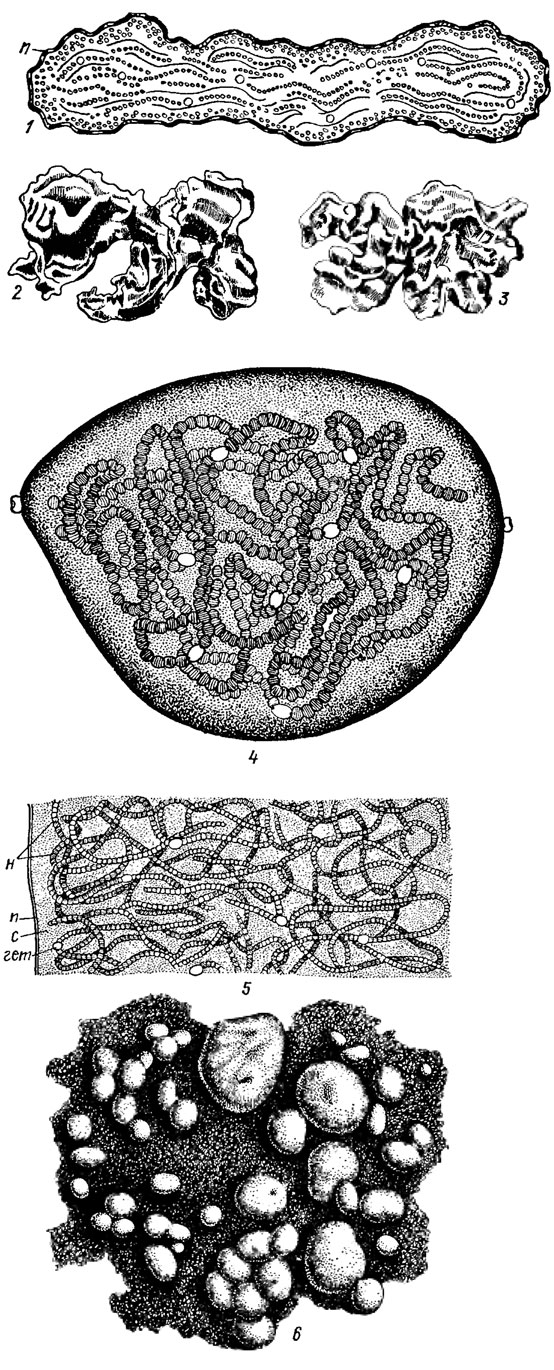
Рис. 1.9. Колонии (колониальные индивиды) Nostoc commune Vauch. sensu Elenk. (1-3 - f. commune, 4-6 - f. sphaericum (Vauch.) Elenk.): 1, 5 - срезы через колонии; 2-4 - отдельные колонии (эвколонин); 6 - общий вид группы колоний на субстрате (н - нити, п - перидерм, с - колониальная слизь, гет - гетероциста)
Химическая природа слизистых обверток и влагалищ изучена недостаточно, но, вероятно, они состоят из очень сложных полисахаридов. Слизистые обвертки (в том числе колониальная слизь) и влагалища нередко окрашены, чаще всего в желто-коричневые цвета. Особенности окраски и строения слизистых обверток и влагалищ синезеленых водорослей зависят от возраста и условий существования индивидов, но все же их нередко используют как видовой признак.
Цитоплазма и плазмалемма. Цитоплазма синезеленых водорослей вязкая, гелеобразная; ее движения и броуновского движения находящихся в ней структур не наблюдали. Плазмолиз получить обычно не удается. Типичных вакуолей (заполненных жидкостью) в здоровых клетках Cyanophyta не найдено*, но в ряде случаев, например в волосках представителей семейства Rivulariaceae, закономерно наблюдается необратимая вакуолизация, ведущая к отмиранию клеток. Иногда вакуолизация возникает как патологическое явление. У отдельных видов встречается своеобразная форма вакуолизации - керитомия, которая обратима.
* (О газовых вакуолях см. ниже.)
Цитоплазма окружена цитоплазматической мембраной - плазмалеммой, примыкающей к внутреннему слою клеточной оболочки (см. рис. 1.6). Считают, что плазмалемма Cyanophyta принципиально не отличается от плазмалеммы бактерий и эукариот. Плазма лемму иногда относят к клеточным покровам, но она является живой составной частью клетки, поддерживающее ее физиологическую целостность, и после разрушения клеточной оболочки лизоцимом сохраняется в составе освобождающихся при этом протопластов (и сферопластов, которые в отличие от протопластов сохраняют остатки разрушенной оболочки), а поэтому ее лучше рассматривать как компонент протопласта. Форму клетки плазмалемма фиксировать не способна. Нередко она образует многочисленные складки (втягиваний, инвагинации), направленные внутрь клетки.
Характерной для эукариот эндоплазматической сети (эндоплазматического ретикулюма) (см. раздел 1.2.2) у Cyanophyta нет, но у них все же существует хорошо развитая мембранная система, к которой, кроме плазма леммы, относят еще мембраны тилакоидов (см. ниже), мезосомы, полимембранные структуры и др.
Фотосинтетический аппарат. Синезеленые водоросли, в отличие от фототрофных бактерий и подобно другим водорослям и высшим растениям, выделяют в процессе фотосинтеза кислород, т. е. обладают способностью к оксигенному фотосинтезу. Типичных хлоропластов они не имеют, но их клетки, как и клетки других водорослей, содержат пластинчатые мембранные структуры (мешочки, диски, ламеллы) - тилакоиды (рис. 1.10; см. рис. 1.6), с которыми связаны ассимиляционные пигменты. Только у одного вида Cyanophyta (Gloeobacter violaceus Rippka) тилакоиды не обнаружены.
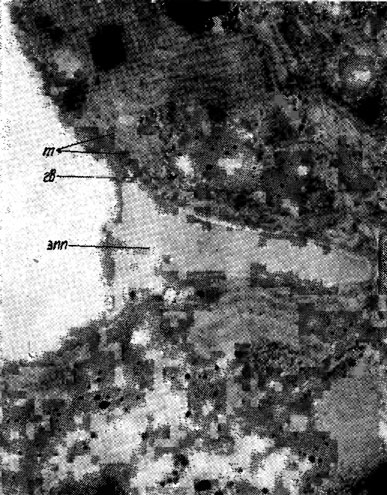
Рис. 1.10. Электронная микрофотография части клетки Microcystis wesenbergii Komarek в состоянии деления: т - тилакоид; гв - газовые вакуоли; зпп - зачаток поперечной перегородки (кольцевидный валик)
Тилакоиды синезеленых водорослей в общем сходны с соответствующими структурами других водорослей. Они располагаются в цитоплазме одиночно или группами. Последние никогда не бывают окружены оболочками, как это наблюдается в эукариотических растительных клетках. Тилакоиды Cyanophyta нередко расположены исключительно в периферической части клетки, параллельно клеточной оболочке, но у многих видов они закономерно заходят в центральную часть клетки или располагаются без особого порядка. В первом случае принятое ранее (до использования методов электронной микроскопии) деление протопласта клетки на хроматоплазму и центроплазму (см. ниже) оправдано, во втором - такое деление затруднительно или невозможно. Тилакоиды могут иметь различные включения. Иногда они контактируют с плазмалеммой и, по-видимому, способны к делению.
К пигментам фотосинтетического аппарата Cyanophyta относятся хлорофилл а, каротиноиды (каротин, ксантофилл) и фикобилипротеиды (аллофикоцианин, фикоциан, фикоэритрин). Последние встречаются в виде специальных структур - фикобилисом, расположенных на поверхности мембран тилакоидов. Кроме си незеленых водорослей фикобилипротеиды найдены только у красных и некоторых криптомонадовых. Хлорофилл b, свойственный растениям, у Cyanophyta не встречен. Имеются и другие отличия.
Таким образом, несмотря на сходство тилакоидов Cyanophyta с соответствующими структурами эукариотических водорослей, а также на наличие у них хлорофилла а, их фотосинтетический аппарат все же существенно отличается от такового других водорослей. Вместе с тем он принципиально отличен и от фотосинтетического аппарата фотоавтотрофных бактерий и Prochlorophyta. Отметим еще, что количество тилакоидов в клетках Cyanophyta, их расположение, концентрация пигментов и другие особенности варьируют не только в зависимости от видовой принадлежности, но и от возрастного и жизненного состояния этих организмов, а также от условий их существования (освещенности, температуры и др.). Поэтому использовать те или иные особенности тилакоидов в целях систематики можно только в тех случаях, когда упомянутые выше факторы хорошо контролированы.
Ядерные эквиваленты. Генетический аппарат. Настоящие, окруженные мембраной, ядра у Cyanophyta отсутствуют. Митоз и типичные хромосомы не обнаружены. Но как и у других организмов, в их клетках содержится дезоксирибонуклеиновая кислота (ДНК) - носитель генетической информации [673]. ДНК располагается обычно в центральной части клеток Cyanophyta в виде мелких гранул или тонких фибрилл (около 2-3 нм в диаметре). Ту часть клетки си незеленых водорослей, в которой содержится ДНК, называют нуклеоплазматической, или ядерной, областью (см. рис. 1.6). Нередко в клетке содержится более одной нуклеоплазматической области. Нуклеоплазматическая область синезеленых водорослей является аналогом ядра эукариот и нуклеоида бактерий и ее можно рассматривать как ядерный эквивалент.
Внешний вид ядерных эквивалентов зависит от методов их исследования. При электронно-микроскопическом исследовании нуклеоплазматическая область клетки имеет фибриллярно-зернистый вид, но при определенных способах фиксации может казаться гомогенной. Еще до начала апектронно-микроскопических исследований с помощью светового микроскопа в протопластах си незеленых водорослей были выделены две области: периферическая окрашенная область - хроматоплазма и внутренняя - центроплазма (центральное тем). Такое деление оправдано только в случае центрального расположения нуклеоплазматической области и периферического расположения тилакоидов. Центроплазма иногда заметна в живых клетках без предварительного их окрашивания. В этих случаях под световым микроскопом середина клетки кажется более светлой. При использовании специальных ядерных красителей с помощью светового микроскопа удается обнаружить особые структуры, содержащие ДНК, - хроматиновые элементы [427]. Последние, по-видимому, соответствуют ядерным эквивалентам. Хромосомами их считать нельзя*.
* (Возможно, что в нуклеоплазматической области клеток синезеленых водорослей имеется структура, сходная с бактериальной хромосомой, имеющей кольцевидную форму и представляющей собой двух цепочечную молекулу ДНК, не связанную (в Отличие от эукариот) с основными белками - гистонами. Правда, в последнее время появились данные о нахождении у синезеленых водорослей белков, подобных гистонам [97]. Имеются также сведения о встречаемости у Cyanophyta небольших внехромосомных молекул ДНК-плазмид.)
Внешний вид и очертания хромата новых элементов зависят от методов окраски и фиксации препаратов водорослей. Обычно они имеют вид отдельные комочков, ниточек, палочек, иногда соединенных между собой и образующих сетчатую или компактную структуру. Полагают, что один хромата новый элемент содержит только один комплекс генетической информации (один геном), но так как хромата новые элементы способны соединяться, о числе геномов в клетке Cyanophyta на основании обычных цитохимических методов исследования судить трудно. Полагают, однако, что большинство мелкоклеточных синезеленых водорослей содержит в клетках по одному геному, тогда как большинство форм с более крупными клетками - полигеномны, и что разные клетки одной многоклеточной особи могут содержать разное количество геномов, что может быть связано с процессом их деления [427].
В настоящее время генетический аппарат синезеленых водорослей изучают более тонкими методами. Показано, что у большинства изученных в этом отношении штаммов одноклеточных Cyanophyta размеры генома составляют (1,6-2,7) ⋅ 109 дальтон*, т. е. сходны с таковыми у бактерий, тогда как у более высокоорганизованных плеурокапсальных и трихомальных форм синезеленых водорослей геномы оказались крупнее (до 8,6 ⋅ 109 дальтон у Calothrix).
* (Дальтон - единица молекулярной массы, равная массе атома водорода.)
Молекула ДНК синезеленых водорослей, как и других организмов, состоит из двух цепочек нуклеотидов. В каждый нуклеотид входит три компонента: азотистое основание, остаток сахара и остаток фосфорной кислоты. Известно четыре типа азотистых оснований ДНК: аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). Разные нуклеотиды отличаются только по азотистым основаниям, а поэтому их названия связывают с названиями входящих в их состав нуклеотидов (например, нуклеотид, содержащий аденин, называется адениловым) и говорят о нуклеотидном составе ДНК организмов, используя при этом символические (буквенные) обозначения, принятые для азотистых оснований. Отношение азотистых оснований А+Т/Г+Ц (а следовательно, и нуклеотидный состав) в ДНК разных организмов различен и может использоваться в систематике. Если нуклеотидный состав двух сравниваемых организмов неодинаков, есть основание говорить об их систематическом отличии. Однако сходство нуклеотидного состава еще не является доказательством таксономического сходства сопоставляемых организмов. В этом случае вероятно как сходство, так и различие. Более тонким методом оценки генетического сходства организмов (в том числе прокариотических) считают метод гибридизации их ДНК. позволяющий судить о ее гомологичности.
Нуклеотидный состав ДНК синезеленых водорослей, определяемый по содержанию (доле) ГЦ в молярных процентах, у синезеленых водорослей варьирует в широких пределах, сходных с пределами, известными для всех прокариот: 35-71 мол.% (см. [674]). Причем встречаются как представители с сильно выраженным АТ-типом (сумма А + Т больше суммы Г + Ц), так и с сильно выраженным ГЦ-типом ДНК. Сведения о содержании ГЦ рекомендуют использовать при классифицировании одноклеточных синезеленых водорослей.
Рибосомы (см. рис. 1.6). Как и у эукариот, рибосомы прокариотических организмов представляют собою рибонуклеопротеидные частицы, включающие рибонуклеиновую кислоту (рибосомальную РНК, или pРНК) и белки, и играют существенную роль в процессе синтеза белка. У синезеленых водорослей рибосомы имеют вид электронно-плотных гранул, размером 10-15 нм, и относятся к 70 S-частицам*. По константе седиментации они сходны с рибосомами в хлоропластах растений и отличаются от находящихся в цитоплазме растительной клетки.
* (70S - константа седиментации, характеризующая скорость, с которой эти частицы осаждаются при центрифугировании в определенных стандартных условиях (S - от имени автора метода - Сведберга).)
Газовые вакуоли (псевдовакуоли). Клетки многих синезеленых водорослей содержат по одной - много газовых вакуолей, видимых при наблюдении в световой микроскоп. Они представляют собой красновато- или темно-коричневые (до черных) тельца, округлой или неправильной формы (рис. 1.11), которые при электронно-микроскопическом наблюдении оказались состоящими из многих палочковидных, плотно расположенных газовых пузырьков, многогранных на поперечном срезе (рис. 1.12). Толщина (ширина) газовых пузырьков у исследованных видов составляет 50-112 нм, а длина в несколько раз больше. Стенка пузырьков белковая, проницаемая для газов. Давление газов внутри пузырьков сходно с давлением в окружающей среде. При повышении давления газов в среде газовые пузырьки сплющиваются (коллапсируют), что сказывается на степени плавучести водоросли [673].
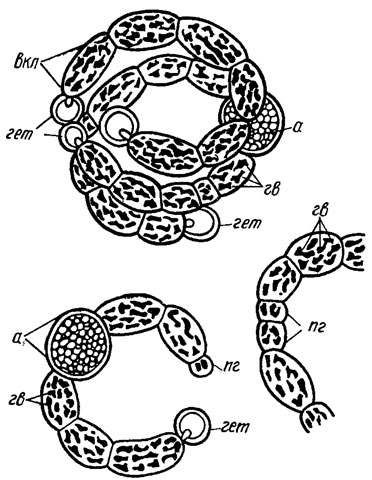
Рис. 1.11. Нити Anabaenopsis elenkinii Miller: а - акинета; гв - газовые вакуоли; гет - гетероциста; вкл - вегетативная клетка; пг - прогетероцисты
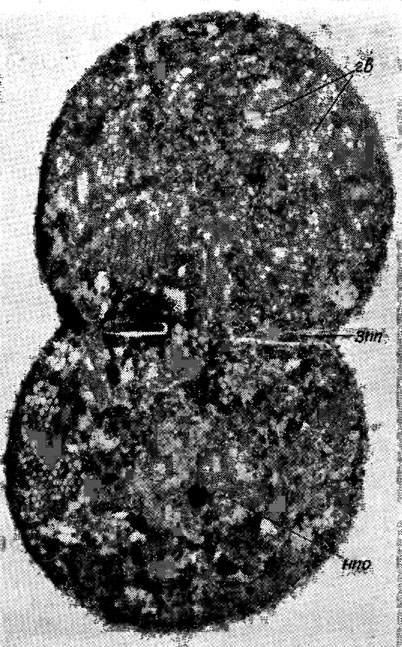
Рис. 1.12. Электронная микрофотография клетки Microcystis wesenbergii Komarek в состоянии деления: гв - газовые вакуоли; зпп - зачаток поперечной перегородки; нпо - нуклеоплазматическая область
Количество газовых вакуолей и способность к их образованию зависит от видовой принадлежности, возраста и условий существования индивида. У некоторых видов (в обычных условиях и местах произрастания водоросли) газовые вакуоли присутствуют в клетках постоянно, у других появляются только на определенных этапах индивидуального развития или при определенных условиях существования.
Другие структуры. Помимо описанных выше структур в клетках Суanophyta содержатся также многие другие. Одни из них встречаются постоянно, присутствие других не обязательно. Ряд структур относится к числу запасных включений, возникающих в результате метаболизма клетки. Многие структуры еще плохо изучены. Некоторые (после их более глубокого исследования) могут оказаться органоидами. Наиболее известны полиэдральные тела (карбоксисомы), цианофициновые (структурированные) гранулы, волютиновые гранулы (гранулы полифосфатов), гранулы гликогена и липидные включения.
Цианофициновые гранулы считают образованиями, специфичными для синезеленых водорослей и являющимися местом запасания азота. Карбоксисомы встречаются также у других прокариот. Их сравнивают с пиреноидами эукариотических водорослей* [648]. В клетках синезеленых водорослей обнаружены также структуры неизвестной природы.
* (Типичные пиреноиды у Cyanophyta отсутствуют.)
Завершая описание строения клетки Cyanophyta, назовем наиболее характерные их черты: отсутствие типичных ядер, окруженных ядерными мембранами; наличие развитой фотосинтетической ламеллярной системы при отсутствии хлоропластов; наличие фибриллярно-зернистой нуклеоплазматической области; присутствие разнообразных включений; наличие ригидных (жестких, негибких) слоистых клеточных оболочек; образование в большинстве случаев слизистых обверток. В связи с тем, что нуклеоплазматаческая область не отграничена мембраной от смежных частей протопласта, полагают, что клетку синезеленых водорослей можно рассматривать как единую физиологическую единицу с тесной биохимической связью между субклеточными структурами, несущими разные функции.
Клетки прокариотических зеленых водорослей (Prochlorophyta) в общем сходны с клетками Cyanophyta. Типичные ядра, окруженные ядерной мембраной, а также хлоропласты, как и у Cyanophyta, отсутствуют. Нет принципиальных отличий и в размере генома (3,59 ⋅ 109 дальтон). Содержание суммы гуанина и цитозина (ГЦ) находится в пределах, характерных для Cyanophyta (31-41 мол.%). Как и у синезеленых водорослей, коэффициент седиментации рибосом Рrосhlоrоn сходен с таковым рибосом хлоропластов эукариот и отличается от установленного для рибосом цитоплазмы эукариот.
В клетках Prochlorophyta имеются тилакоиды и хлорофилл а. Их каротиноидный комплекс сходен с таковым синезеленых водорослей. Имеется сходство также в тонком строении и биохимическом составе клеточных оболочек, Однако, как уже упоминалось выше, в клетках прокариотических зеленых водорослей, в отличие оттого, что характерно для Cyanophyta, наряду с хлорофиллом а присутствует хлорофилл b, а фикобилипротеиды не найдены. Не обнаружен также цианофицин, являющийся характерным компонентом клеток синезеленых водорослей.
Деление клетки
Для прокариотических водорослей характерно деление клеток надвое, происходящее без сложных структурных изменений, сопровождающих митоз у эукариотических организмов. При делении клеточного тела происходит формирование поперечных перегородок, способ образования которых принципиально отличается от свойственного для эукариот. Новая поперечная перегородка первоначально возникает в виде кольцевидного валика (см. рис. 1.6; 1.10), который постепенно растет от периферии к центру клетки, где смыкается. В местах образования зачатка новой поперечной перегородки нередко образуются перетяжки (перешнуровки) (см. рис. 1.2, 4, 5), наличие которых у трихомальных форм Cyanophyta нередко рассматривается как таксономический признак. У одноклеточных форм (в том числе Prochlorophyta) перетяжка со временем смыкается, что ведет к отделению дочерних клеток. Формирование новых поперечных перегородок у ряда видов Cyanophyta может начинаться еще до окончания предыдущего деления клетки. Это явление, особенно характерное для представителей семейства Oscillatoriaceae, получило название ускоренного деления (рис. 1.13, 1).
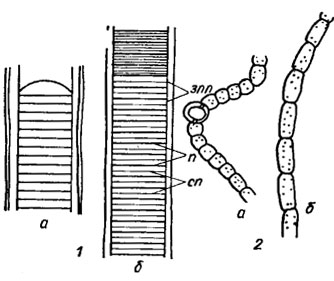
Рис. 1.13. Части нитей Lyngbya majuscula (Dillw.) Harw. (1) и Nostoc linckia (Roth.) Born. et Flah f. spongiaeforme (Ag.) Kütz. (2), клетки которых находятся в состоянии менее интенсивного (а) и более интенсивного (б) деления. нпп - новые поперечные перегородки, закладывающиеся до окончания формирования предыдущих (ускоренное деление); п - старые поперечные перегородки; сп - поперечные перегородки среднего возраста
По немногочисленным данным, полученным при помощи светового микроскопа, делению клеток Cyanophyta предшествует деление хроматиновых элементов, происходящее, вероятно, в продольном направлении, поскольку хроматиновые элементы расходятся к полюсам, скользя один вдоль другого [427]. Флуорометрическими методами, на примере отдельных видов Cyanophyta, показано, что количество ДНК перед делением клеток удваивается, а после деления уменьшается вдвое.
Деление клеток одноклеточных форм Cyanophyta происходит в одной, двух или трех (многих?) плоскостях. У колониальных форм это приводит к возникновению разных типов размещения клеток. Для многоклеточных трихомальных индивидов синезеленых водорослей характерно деление клеток в одной плоскости - поперечной к продольной оси трихома. Но у некоторых форм клетки одного трихома способны делиться в двух-трех плоскостях, что ведет к образованию трихомов с настоящей многорядностью (рис. 1.14).
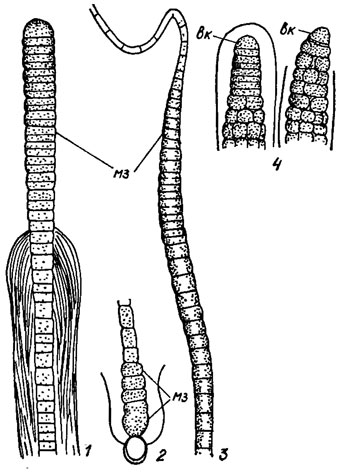
Рис. 1.14. Зоны активного деления клетки ('меристемальные зоны'): 1 - верхушечная (Scytonema myochrous (Dillw.) Ag.); 2 - базальная (Calothrix wembaerensis Hieron. et Schmidle); 3 - интеркалярная (Rivularia haematites (D. C.) Ag.); 4 - нарастание трихома с 'настоящей' многорядностью (Stigonema sp.) за счет верхушечной клетки; мз - 'меристемальная' зона; вк - верхушечная клетка
К делению способны фактически все клетки Cyanophyta, которые имеют живое содержимое (наблюдали, например, деление протопласта гетероцист, приостановку развития молодых акинет и возврат их к делению, свойственному вегетативным клеткам). Но у морфологически наиболее прогрессивных форм синезеленых водорослей встречаются особые зоны активного деления клеток, т. н. меристемальные зоны (базальная, интеркалярная, верхушечная), а иногда даже специальные верхушечные клетки, напоминающие верхушечные клетки более высокоорганизованных эукариотических водорослей (см. рис. 1.14).
В результате деления клеток прокариотических водорослей чаще всего возникают две внешне одинаковые клетки. Но у некоторых таксонов (например, у представителей рода Anabaenopsis (Wotosz.) Miller перед образованием гетероцист; см. рис. 1.8; 1.11) наблюдается неравное деление клеток, в результате которого одна из возникших клеток имеет меньшие размеры. В отдельных случаях неравное деление клеток напоминает почкование.
Закономерности клеточных циклов у прокариотических водорослей не изучены. Известно, что в одних случаях делению предшествует рост клетки, а в других - клетки делятся без предварительного удлинения. На уровне световой микроскопии (без применения цитохимических методов) у форм с округленными клетками (шаровидными, овалоидными, эллипсоидными) имеются по крайней мере три возрастных состояния клетки: зрелости, цитотомии (т. е. деления клеточного тела) (см. рис. 1.12) и молодости, между которым клетка находится в переходных состояниях. Есть основания полагать, что частота встречаемости клеток, пребывающих в определенном состоянии, у разных видов неодинакова и зависит от условий существования водорослей.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'