Введение (Н. П. Масюк, Н. В. Кондратьева)
Мир растений, насчитывающий около 0,5 млн. видов, занимает огромные площади континентов земного шара и акватории Мирового океана, на которых образует почти сплошной покров. Даже в Антарктиде на свободных ото льда местах растут мхи и лишайники, а поверхность ледяных и снежных пустынь нередко покрыта разрастаниями одноклеточных водорослей. Растительность суши и океана осуществляет на Земле уникальную функцию превращения в процессе фотосинтеза под воздействием солнечного света неорганических соединений в органические, создавая тем самым возможность существования на нашей планете иных форм жизни, включая человека. Хотя в мире растений существуют и нефотосинтезирующие организмы, наиболее характерной и важной в глобальном масштабе особенностью растений является их способность к фотосинтезу, к биоконсервации солнечной энергии и углекислого газа атмосферы в виде органических соединений, к выделению в ходе этого процесса кислорода, необходимого для дыхания всех аэробных организмов, доминирующих на нашей планете. Таким образом, зеленый экран суши и океана представляет собой гигантский фотосинтетический реактор, обеспечивающий жизнь на Земле в целом.
Кроме способности к фотосинтезу растения имеют и другие своеобразные черты, отличающие их от представителей животного мира. Это и специфическое строение клетки, и преимущественно неподвижный образ жизни, и тенденция к неограниченному росту в течение вегетативной фазы жизненного цикла при высокой степени расчлененности тела. Однако мир растений настолько многообразен, что ни один из указанных выше признаков не является общим для всех его представителей. Таких общих для всех растений признаков практически вообще нет. У некоторых простейших одноклеточных растений, как и у животных, могут отсутствовать клеточная оболочка, тонопласт и даже пластиды. Наряду с неподвижными существует множество простейших растений, активно передвигающихся в пространстве в вегетативном состоянии и неотличимых по этому признаку от представителей животного мира. Гиганты растительного мира, населяющие воду и сушу, уживаются с микроскопическими организмами, размеры которых не превышают долей микрометра; сложно расчлененные многоклеточные формы - с простейшими одноклеточными, шаровидными или каплевидными, не дифференцированными на какие-либо органы. Наряду с зелеными существуют растения, окрашенные в сине-зеленый, желто-зеленый, голубой, фиолетовый, розовый, красный, до темно-красного и почти черного, желтый, бурый, золотистый цвета, а также бесцветные. Наряду с фотоавтотрофными к миру растений относят также некоторые связанные с ними тесным родством гетеротрофные - сапротрофные, паразитические и голозойные организмы. Известны растения, онтогенез которых продолжается сотни лет, и организмы, жизнь которых от рождения до смерти длится всего несколько часов.
С давних времен все растения разделяют на высшие, объединяющие около 300-350 тыс. видов, и низшие - около 150-200 тыс. видов. В качестве основного критерия этого деления используют морфологический признак - наличие или отсутствие дифференциации вегетативного тела растения на многоклеточные органы, выполняющие различные функции (корень, стебель и листья). К низшим относят организмы, тело которых не расчленено на органы, а представлено недифференцированным талломом, или слоевищем. Поэтому низшие растения получили название слоевцовых, или талломных (Thallophyta), в отличие от высших - листостебельных, или кормофитов (Cormophyta). Половые органы низших растений, если они имеются, - одноклеточные, высших - многоклеточные. В отличие от высших, низшие растения характеризуются более простым анатомическим строением. У них отсутствует стела - центральный цилиндр, включающий в себя проводящие элементы (сосуды, ситовидные трубки). Поэтому низшие растения называют также бессосудистыми (Plantae eellulares), в отличие от высших - сосудистых (Plantae vasculares). Термин "бессосудистые растения" шире по объему, чем термин "низшие растения", поскольку кроме собственно низших растений, к которым обычно относят вирусы, бактерии, водоросли, грибы и лишайники (сравни [35])*, охватывает также мохообразные. В отличие от высших цветковых, или семенных, растений низшие растения называют также споровыми, в связи с тем, что они никогда не образуют цветков и семян, но могут размножаться спорами. Однако понятие "споровые растения" еще шире, чем "низшие" или "бессосудистые", поскольку к споровым принадлежат также хвощи, плауны и папоротники. Отсутствие цветков послужило основанием для присвоения низшим растениям еще одного названия - тайнобранные (Cryptogamеn). Более четкое представление об объеме упомянутых терминов (в их традиционном понимании) дает следующая схема.
* (Сейчас объем царства растений, а следовательно и группы низших растений, пересматривается (см. ниже).)
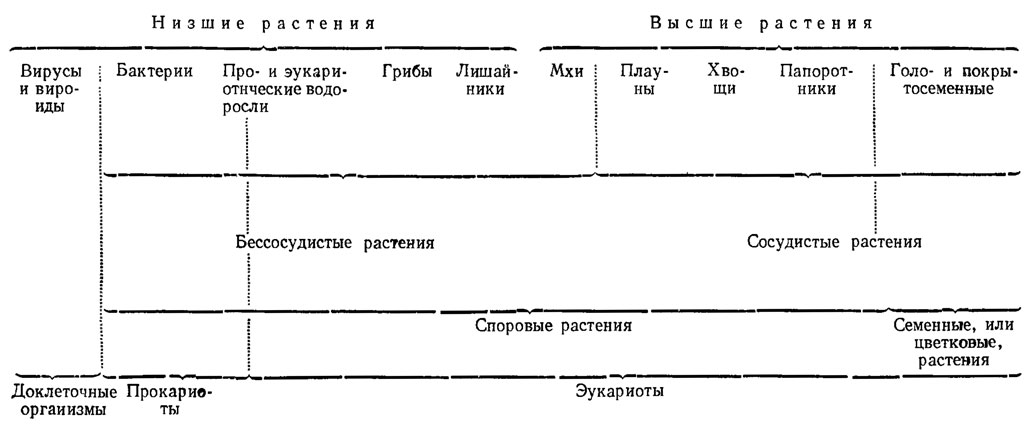
Схема о представление об объеме упомянутых терминов (в их традиционном понимании)
Открытие в органическом мире двух кардинально различающихся типов строения клетки - прокариотического и эукариотического, между которыми отсутствуют какие-либо промежуточные формы*, привело к пересмотру критериев деления органического мира на царства. В свете этих данных многие ученые разделяют органический мир на царства прокариот (Procaryota) и эукариот (Eucaryota) [120, 372, 376, 584 и др.]. Вирусы и вироиды, не имеющие клеточной организации, в этой системе составляют отдельную группу до клеточных организмов.
* (У так называемых мезокариотических организмов (см. разделы 1.2 и 8) в отличие от прокариот имеется ядро, окруженное ядерной оболочкой, в связи с чем их правильнее рассматривать среди эукариот.)
Царство Procaryota насчитывает около 5 тыс. видов, включая бактерии (в широком смысле) и прокариотические водоросли. Все остальные живые существа, населяющие нашу планету (примерно 1,5 млн. видов) и имеющие клеточное строение, относятся к царству Eucaryota.
Некоторые ученые рассматривают прокариот и эукариот в ранге надцарств [54, 142, 303], а в пределах иадцарства Procaryota выделяют два царства: бактерии (Bacteria) и фотосинтезирующие прокариотические организмы (Photoprocaryota) [142, 143], а в пределах надцарства Eucaryota - три царства: растения (Vegetabilia), животные (Animalia) и грибы (Мусеtalia) [303]*. Концепции пяти царств придерживаются Л. Маргелис и К. Шварц [192, 542], причем в одно из этих царств (Мопега) они объединяют все прокариотические организмы, а к остальным четырем относят эукариотические (растения - Plantae, животные - Animalia, грибы - Fungi и примитивные эукариоты - Protoctista). В последнее время, в биологической литературе наблюдается тенденция к еще большему увеличению количества царств живой природы - до семи или даже до девяти [368]. С другой стороны, Л. Диллон [389], напротив, считает, что биохимическое и клеточное единство жизни наилучшим образом может быть отражено в системе, включающей одно единственное царство живых существ.
* (Всего в системе А. Л. Тахтаджяна [303] насчитывается четыре царства; дробянки, растения, животные, грибы.)
Имеются также разногласия по поводу того, на каком уровне - прокариотическом или эукариотическом - произошла дифференциация живых организмов на растения и животные. Нередко это важное событие в биологической эволюции относят к эукариотическому уровню, в связи с чем прокариотические водоросли исключают из системы растительного мира [54, 120, 303, 405]. Однако известно, что фотоавтотрофный тип питания возник еще на прокариотическом уровне жизни и уже на этом уровне определил развитие живой материи по растительному пути [142].
Водоросли - слоевцовые бессосудистые споровые растения - являются типичными представителями низших растений. Они характеризуются способностью к оксигенному фотосинтезу (фотоавтотрофному способу питания). Правда, среди них известны и бесцветные, и даже беспластидные (апопластидные) организмы, питающиеся гетеротрофно (сапротрофы, паразиты и организмы с голозойным способом питания). Однако в преобладающем большинстве случаев их генетические связи с окрашенными фотоавтотрофными формами не вызывают сомнений, и поэтому они рассматриваются как вторично гетеротрофные организмы, утратившие способность к фотоавтотрофному способу питания в результате приспособления к особым условиям существования в богатых органикой биотопах.
Водоросли - преимущественно обитатели водной среды, но многие (по-видимому, вторично) приспособились к жизни во вневодных местообитаниях - в почве и на ее поверхности, на скалах, стволах деревьев и в других наземных биотопах.
Понятие "водоросли" (Algae) является не систематическим, а биологическим. К водорослям относится ряд отделов растительного мира. Возникнув в разное время и, по-видимому, от разных предков, различные группы водорослей в дальнейшем развивались самостоятельно, но в результате конвергентной эволюции в сходных условиях обитания приобрели много сходных черт. Эта гетерогенность группы Algae нашла отражение в современных системах органического мира [120, 368, 371, 372 и др.], в которых водоросли распределены между разными царствами (надцарствами) живой природы и расчленены на большое количество отделов и классов (см. раздел 8.5). В основу настоящего издания положено деление водорослей на отделы, принятое Д. К. Зеровым [120], который относил водоросли к двум царствам: Procaryota и Eucaryota. Однако в систему Д. К. Зерова (см. табл. 8.1) внесены некоторые изменения. В нее введен недавно открытый отдел прокариотических зеленых водорослей (Prochlorophyta). Согласно требованиям "Международного кодекса ботанической номенклатуры" название отдела Chloromonadophyta заменено на Raphidophyta. Из-за неоднозначной трактовки объема отдела Pyrrhophyta мы употребляем наименование Dinophyta. Зеленые водоросли рассматриваем в качестве самостоятельного отдела Chlorophyta [306], из которого выделен отдел харовых - Charophyta. В связи с этим в настоящем справочнике принято следующее деление водорослей на отделы:
* (Систематическое положение прокариотических водорослей и вопросы номенклатуры Cyanophyta рассмотрены в разделе 9а. 1.)
Отдел 1. Синезеленые водоросли - Cyanophyta
Отдел 2. Прокариотические (первичные) зеленые водоросли - Prochlorophyta
Отдел 1. Эвгленовые водоросли - Euglenophyta
Отдел 2. Динофитовые водоросли - Dinophyta
Отдел 3. Криптофитовые водоросли - Cryptophyta
Отдел 4. Рафидофитовые водоросли - Raphidophyta
Отдел 5. Золотистые водоросли - Chrysophyta
Отдел 6. Диатомовые водоросли - Bacillariophyta
Отдел 7. Желтозеленые водоросли - Xanthophyta
Отдел 8. Красные водоросли - Rhodophyta
Отдел 9. Бурые водоросли - Phaeophyta
Отдел 10. Зеленые водоросли - Chlorophyta
Отдел 11. Харовые водоросли - Charophyta
Науку о водорослях - альгологию (от лат. alga - морская трава, водоросль и греч. λoγoς - слово, учение), или фикологию (от греч. φüχоς - водоросль), - рассматривают как самостоятельный раздел ботаники. Водоросли изучают в морфологическом, цитологическом, генетическом, онтогенетическом, физиолого-биохимическом, экологическом, флористическом, систематическом, эволюционно-филогенетическом и некоторых других направлениях. Нередко их используют в качестве объектов для решения вопросов, относящихся к области других наук (биохимии, биофизики, генетики и др.). Данные альгологии нередко учитывают при разработке общебиологических проблем, а также хозяйственных задач.
Развитие прикладной альгологии происходит в трех основных направлениях: 1) обоснование способов непосредственного использования водорослей в медицине и в различных областях хозяйства; 2) поиски путей использования водорослей для решения природоохранных вопросов; 3) накопление данных о водорослях для решения задач других отраслей.
В течение последних десятилетий в развитии альгологии произошел качественный скачок, выдвинувший ее в ряд передовых наук современности. Особое значение для развития альгологии имело усовершенствование электронно-микроскопической техники и методов биохимического анализа, а также разработка способов выделения аксенических культур и создание в разных странах коллекций штаммов водорослей. Все это стимулировало развитие экспериментальных работ. Расширение объема альгологических исследований привело к существенному увеличению информации о водорослях. В результате произошла дальнейшая дифференциация альгологии, и на стыке альгологии с другими науками возникли новые научные направления (палеоальгологическое, радиобиологическое и др.).
В недалеком будущем следует ожидать вычленения из альгологии дисциплин, аналогичных тем, которые уже выделились из раздела ботаники, посвященного высшим растениям. Стремление к новому, в общем созвучное с современным стилем научного мышления, привело к определенному перераспределению интересов исследователей водорослей: намного возросла, например, доля физиолого-биохимических и экологических работ, но значительно сократилась доля исследований, проводимых в традиционных (морфологическом, систематическом и др.) направлениях. Вместе с тем недооценка значимости работ в традиционных направлениях (содержание которых ничем заменить нельзя) опасна, она может привести даже к кризисному состоянию альгологии. Необходимо гармоническое развитие всех направлений альгологии (как новых, так и традиционных) в сочетании с глубоким целенаправленным теоретическим и методологическим анализом и синтезом полученных знаний. Такой подход приведет не только к вычленению из альгологии частных отраслей, но и к становлению общей альгологии как самостоятельного раздела науки о водорослях, специально посвященного общим закономерностям структурной организации и функционирования низших фотоавтотрофных организмов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'