Ауксоспоры
Диатомовые водоросли - единственная группа среди растительных организмов, у которых в жизненном цикле имеется стадия ауксоспорообразования (однако, по последним данным, у некоторых диатомей эта стадия не наблюдалась). Первые исследователи рассматривали процесс образования ауксоспор как стадию омоложения вегетативной клетки. Было известно, что по мере деления клеток последние прогрессивно уменьшаются в размерах до определенного предела, свойственного виду, после которого клетка, сбросив панцирь, остается в пектиновой оболочке и начинает увеличиваться до первоначального своего размера, а затем вырабатывает новый панцирь. (Табл. VII, 1-4). Этот процесс и считали омоложением клетки. Однако дальнейшие наблюдения показали, что процесс ауксоспорообразования у некоторых видов сопровождается половым процессом,, это вызвало большой интерес, и многие исследователи занялись изучением полового процесса у диатомей (сводки работ см.: Fritsch, 1935; Geitler, 1957). Особенно много работ этой проблеме посвятили Гейтлер (Geitler; более 20 работ в период 1939-1966 гг.) и Штош (Stosch; 8 работ с 1950 по 1966 г.).
Этими исследованиями были охвачены очень многие виды диатомей. Полученные данные показали следующее: не у всех исследованных видов наблюдались ауксоспоры, созреванию ауксоспоры предшествует мейозис с последующим формированием гаплоидных гамет (Geitler, 1952а, 1952b; Stosch, 1954); было окончательно подтверждено, что все диатомовые водоросли диплоидные, гаплоидными являются только гаметы. Диатомеи однодомные, некоторые гомота л личные, до сих пор известен один двудомный вид Rhabdonema adriaticum (Stosch, 1958). Половой процесс бывает нормальным и редуцированным, встречаются апогамия и партеногенез, тип полового процесса независим от систематического положения диатомей, у близких видов могут быть различные типы, а у далеких сходные.
Таким образом, подтвердилось мнение Шмидта (Schmidt, 1927b) о диплоидности всех диатомей и данные Б. М. Персидского (1932, 1935) о наличии полового процесса у представителей класса Centricae, тогда как ранее считалось, что диатомеи класса Pennatae диплоидные, а класса Centricae гаплоидные и что у представителей последнего класса половой процесс отсутствует. В период 1952-1968 гг. опубликован ряд работ с гистологическим описанием оогамного процесса у диатомей из родов Lithodesmium (Stosch, 1954; Manton, Stosch, 1966), Biddulphia, Streptotheca (Stosch, 1954), Stephanopyxis (Stosch, Drebes, 1964; Drebes, 1964, 1966), Melosira (Stosch, 1958; Erben, 1959), Rhabdonemaw др. (Stosch, 1958; Rozume 1968; Schultz, Trianor, 1968). Исследования Штоша показали, что оогамнь половой процесс свойствен представителям не только класса Centricae, я также и класса Pennatae, чем он окончательно установил монолитной отдела Bacillariophyta и ошибочность деления его на 2 класса.
Исследования полового процесса у диатомей при ауксоспорообразовании (Fritsch, 1935; Geitler, 1952а, 1952b, 1957; Patrik, Reimmer, 1967 Takano, 1967a) выявили несколько типов: нормальный половой процесс - изогамный, анизогамный и оогамный и 3 формы редуцированного полового процесса.
1. Изогамный процесс протекает различно. Две материнские клетки производят в результате деления по две неподвижные гаметы. При этом обе материнские клетки сближаются, покрываются слизистым чехлом раздвигают створки и неподвижные гаметы копулируют попарно, образуя две зиготы, производящие две ауксоспоры. Этот тип свойствен многим видам родов Amphora, Amphiprora, Epithemia, Rhopalodia и Surirellt ovalis.
2. Анизогамный (гетерогамный) процесс протекает двояко. В первом случае две материнские клетки сближаются и покрываются слизистым чехлом. Ядро в каждой клетке делится дважды, первое деление мейотическое, второе митотическое, в результате образуется два одноядерных протопласта (гаметы), а остальные ядра дегенерируют. Одна гамета подвижная, другая неподвижная. Подвижная гамета из своей материнской клетки передвигается в другую материнскую и сливается с неподвижной, в результате в каждой материнской клетке образуется одна зигота, которая развивается в ауксоспору. Это характерно для большинства представителей сем. Naviculaceae и некоторых видов Nitzschia. (Табл. VII, 5-8). Во втором случае в одной материнской клетке обе гаметы неподвижные, а во второй - обе подвижные, переходящие в материнскую клетку с неподвижными гаметами, в результате их слияния образуются две зиготы, дающие две ауксоспоры (это наблюдается у Navicula halophila, Synedra ulna, S. rumpens var. fragilarioides и др.
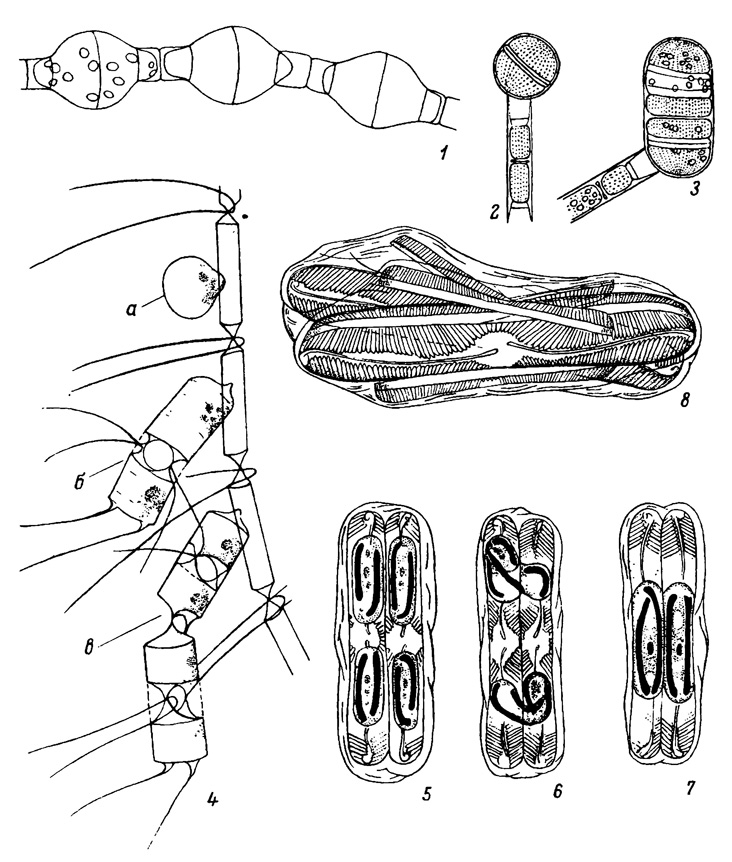
Таблица VII. Образование ауксоспор и половой процесс (1-3 - по Скабичевскому, 1960; 4 - по Прошкиной-Лавренко, 1955; 5-8 - по Hendey, 1964). 1 - Melosira varians Ag., в цепочке три ауксоспоры; 2,3 - M. baicalensis (Meyer) Wisl.: 2 - зрелая ауксоспора, 3 - прорастание ауксоспоры в цепочку; 4 - Chaetoceros curvisetus Cl. с одной молодой ауксоспорой (а) и двумя прорастающими в цепочки (б, в); 5-8 - Pinnularia sp., анизогамный половой процесс: 5 - в двух сближенных клетках, окруженных общей слизью, путем редукционного деления возникают две гаметы, подвижная и неподвижная, 6 - подвижная гамета мигрирует в соседнюю клетку и сливается с неподвижной гаметой, 7 - образовавшиеся две зиготы, увеличиваясь в размерах, превращаются в две ауксоспоры, 8 - ауксоспоры покрылись панцирем, образовав новые вегетативные клетки, рядом с ними - остатки панцирей материнских клеток
3. При оогамном процессе женская материнская клетка (оогоний) производит одну яйцеклетку (Stephanopyxis, Melosira varians) или две (Biddulphia mobiliensis), а мужская материнская клетка (сперматогоний) производит два сперматозоида (Melosira varians) или четыре (Biddulphia rhombus, Cerataulina smithii, Rhabdonema adriaticum, Cyclotella sp.). Сперматозоид оплодотворяет яйцеклетку, образующую зиготу, которая без стадии покоя дает ауксоспору. (Табл. VIII).
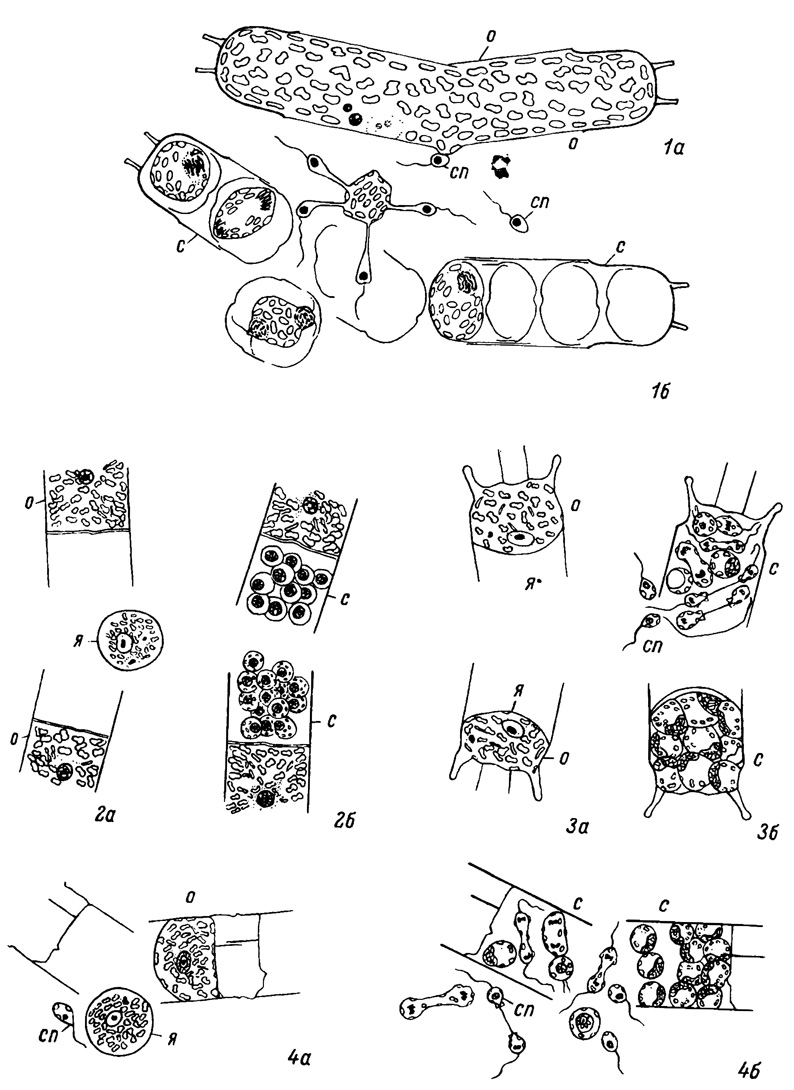
Таблица VIII. Оогамия у центричных планктонных диатомей (1 - по Stoscb, Drebes, 1964; 2-4 - по Stosch, 1954). 1а, 1б - Stephanopyxis turris (Grev. et Am.) Ralfs; 2a, 2б - Lithodesmium sp.; 3a, 3б - Biddulphia . mobiliensit Bail.; 4a, 4б - Streptotheca sp. о - оогоний, с - сперматогоний, сп - сперматозоид, я - яйцеклетка
У всех диатомей (пеннатных и центричных) митоз и мейозис протекают одинаково (Stosch, 1951-1958; Geitler, 1957; Erben, 1959; Stosch, Drebes, 1964), причем при митозе в стадии метафазы хромосомы образуют экваториальное кольцо, а не пластинку, которая более свойственна животным клеткам.
Время, необходимое для слияния мужской и женской гамет, сильно варьирует, так, например, у Navicula seminulum это занимает 2-3 мин., а у Amphipleura pellucida - до часа. После слияния половых клеток образуется зигота, которая немедленно увеличивается в размерах, образуя ауксоспору, оболочкой которой служит перизониум. Когда ауксоспора созреет, в ней возникает вегетативная клетка, в которой сначала образуется эпитека, края ее загибаются, но поисковый ободок отсутствует, часто эпитека имеет не вполне точную структуру, характерную для вида. Позже образуется гипотека, вполне нормальная, с поясковым ободком. Таким образом, обычно первая вегетативная клетка по внешнему виду не совсем симметричная, но в процессе дальнейших делений эпитека принимает нормальную форму.
Редуцированный половой процесс протекает в различных формах, .... ауксоспоре наблюдаются процессы партеногенеза, ...., происходит вегетативное развитие зиготы:
.... клетка развивается в зиготу (и ауксоспору) через .... (Chaetoceros borealis, Ch. densus, Grammatophora и др.);
.... развивает партеногенетически одну ауксоспору, Cymatopleura, Cymbella, а также Meridion .... Surirella .... и др.);
.... ауксоспору вегетативно (Melosira .... требует проверки.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'