Фотосинтез
Диатомовые водоросли преимущественно автотрофные растения, у них, как и у прочих автотрофных организмов, процесс образования органического вещества происходит в хлоропластах с помощью пигментов в процессе фотосинтеза. Первоначально было установлено, что пигменты у диатомей состоят из смеси хлорофиллов с ксантофиллами и фукоксантином. Позже для уточнения состава пигментов у диатомей был применен хроматографический метод, с помощью которого было выяснено присутствие в хлоропластах диатомей восьми пигментов (Dutton, Manning, 1941; Strain, Manning, 1942, 1943; Strain a. oth., 1943, 1944; Wassink, Kersten, 1944, 1946; Cook, 1945; Hendey, 1964). Эти пигменты следующие: хлорофилл α, хлорофилл с, β-каротин, фукоксантин, диатоксантин, диадиноксантин, неофукоксантин А и неофукоксантин В. Последние четыре пигмента входят в состав диатомина, открытого ранее. Некоторые авторы указывают еще на минимальное присутствие ксантофилла и феофитина (Strain а. oth., 1944).
Общее количество пигментов у диатомей составляет в среднем около 16% фракции липидов, но содержание их разное у различных видов. В литературе имеется очень мало данных о количественном содержании пигментов у морских планктонных диатомей, а для бентосных видов, которые особенно богаты желтыми и бурыми пигментами, данные почти отсутствуют (табл. 1 и 2).
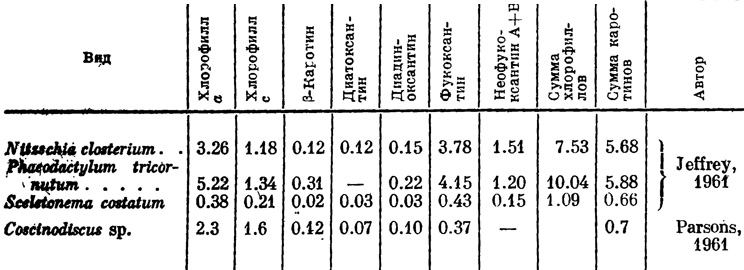
Таблица 1. Содержание пигментов в морских диатомеях (в мг/г сухого веса) (по Lewin, Guillard, 1963 : 405)
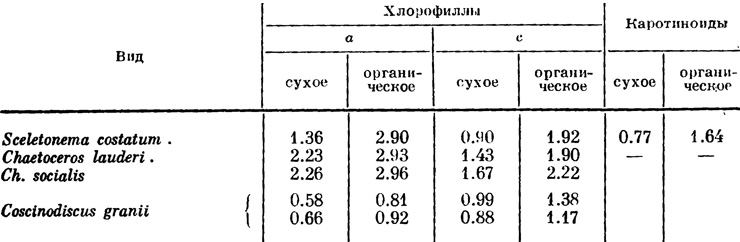
Таблица 2. Содержание пигментов в черноморских морских планктонных диатомеях (в % от вещества) (по Финенко, Ланской, 1968 844)
Вышеприведенные данные показывают, что содержание пигментов варьирует даже у одного и того же вида. Имеются сведения о том, что содержание пигментов подвержено колебаниям в зависимости от интенсивности света, его качества, содержания питательных веществ в среде, от состояния клетки и ее возраста. Так, например, изобилие в среде питательных веществ при относительно слабой интенсивности света стимулирует продуктивность пигментов, и наоборот, высокая интенсивность света при недостатке питательных веществ в среде ведет к уменьшению концентрации пигментов. При недостатке фосфора и азота содержание хлорофилла а может снизиться в 2.5-10 раз (Финенко, Ланская, 1968). Установлено, что с возрастом клетки уменьшается содержание хлорофилла с.
Функции пигментов, кроме хлорофиллов, у диатомей еще недостаточно выяснены. Хлорофилл α является основным пигментом, поглощающим световую энергию всех лучей спектра, причем он имеет две формы, отличающиеся усвоением света: одна из них возбуждается непосредственно красным светом, а вторая, кроме того, также и энергией, передаваемой вспомогательным пигментом фукоксантином (Emerson, Rabinowitch, 1960). Остальные пигменты являются вспомогательными к хлорофиллу а, но они также играют относительно важную роль при фотосинтезе. Хлорофилл с имеет более высокий максимум поглощения в синей области, чем в красной, и следовательно, он способен утилизировать световые лучи меньшей длины волн, максимум поглощения у него лежит при 520-680 нм и падает до нуля при длине волны 710 нм, поэтому его абсорбция более интенсивна в зоне голубого света, т. е. на глубинах 10-25 м от поверхности воды, где хлорофилл а менее эффективен. Роль β-каротина недостаточно ясна, его спектр поглощения резко обрывается при 500 нм, что свидетельствует о его способности абсорбировать в лучах длины волн 500-560 нм, т. е. в области зелено-желтого света (в воде на глубинах 20-30 м). Таким образом, β-каротин передает усвоенную энергию хлорофиллу α (Dutton, Manning, 1941). Это известно, например, для Nitzschia dissipata, которая абсорбирует энергию в области зелено-желтого света (Wassink, Kersten, 1944, 1946). Бурые пигменты из группы фукоксантинов имеют максимум поглощения при длине волн около 500 нм и, по-видимому, обеспечивают фотосинтез диатомовых водорослей на глубинах 20-50 м путем передачи хлорофиллу поглощенной ими энергии. Даттон и Меннинг (Dutton, Manning, 1941), а позже Вассинк и Керстен (Wassink, Kersten, 1946) показали, что фукоксантин у диатомей является главным из дополнительных пигментов. Свет, абсорбируемый фукоксантином, утилизируется для фотосинтеза почти с такой же эффективностью, как свет, абсорбируемый хлорофиллом. Этого не наблюдается у зеленых и синезеленых водорослей, лишенных фукоксантина. Танада (Tanada, 1951) также выяснил, что у пресноводной диатомеи Navicula minima var. atomoides фукоксантин поглощает сине-голубой свет (450-520 нм) и утилизирует его так же эффективно, как свет, абсорбированный хлорофиллом. Хенди (Hendey, 1964) указывает длину световых волн, при которых происходит максимальная абсорбция света различными пигментами диатомей. В ацетоне они следующие (в ммкм): хлорофилл α - 430 и 663-665, хлорофилл с - 445 и 630, β-каротин - 452-456, фукоксантин - 449, диатоксантин - 450-452, диадиноксантин - 444-446, неофукоксантин А - 448 - 450 и неофукоксантин В - 448.
Химизм фотосинтеза у диатомей, по-видимому, несколько иной, чем у других растительных организмов, у которых конечным продуктом фото-синтеза являются углеводы, тогда как у диатомей - жиры. Исследования с помощью электронного микроскопа не обнаружили присутствия крах-мала ни в строме хлоропластов, ни вблизи пиреноидов. Фогг полагает, что у диатомей конечным продуктом ассимиляции также являются углеводы, но в быстро идущих дальнейших процессах метаболизма они превращаются в жиры (Collyer, Fogg, 1955; Fogg, 1956). Химический состав жиров у диатомей неизвестен ни для продуктов ассимиляции, ни для запасных питательных масел и масляных телец (Goulon, 1956).
В океанах, морях и пресноводных водоемах у поверхности воды условия для фотосинтеза близки к условиям в воздушной среде, но с погружением в глубину они меняются в связи с изменением интенсивности и качества света. В отношении освещенности различают три зоны: эуфотическую - от поверхности до 80 м глубины, в ней происходит фотосинтез; дисфотическую - от 80 до 2000 м, здесь некоторые водоросли еще встречаются, и афотическую - ниже, в которой свет отсутствует (Das, 1954 и др.). Фотосинтез морского и пресноводного фитопланктона в поверхностном слое воды достаточно исследован как в природных, так и в культуральных условиях (Wassink, Kersten, 1944, 1946; Вотинцев, 1952; Tailing, 1955, 1957а, 1966; Ryther, 1956; Edmondson, 1956; Ryther, Menzel, 1959; Steemann Nielsen, Hensen, 1959,1961, и др.). В частности, круглогодичные наблюдения в Черном море показали, что наибольшая интенсивность фотосинтеза фитопланктона совпадает с наибольшей солнечной радиацией. Летом максимальный фотосинтез фитопланктона наблюдается в период от И до 16 час. (Ланская, Сивков, 1949; Бессемянова, 1957). У разных планктонных видов максимальная интенсивность фотосинтеза имеет пределы изменений, свойственных тому или иному виду. При этом большое значение имеет широтное расположение акваторий (Doty, 1959 и др.).
Среди диатомей (как планктонных, так и бентосных) существуют светолюбивые и тенелюбивые виды, у которых различны интенсивность фотосинтеза и коэффициент использования солнечной энергии при одинаковой радиации. У светолюбивых видов, как Cerataulina bergonii (планктонная) и Navicula pennata var. pontica (сублиторальная), фотосинтез идет параллельно радиации и достигает максимума в полдень, а у тенелюбивых - Thalassionema nitzschioides (планктонная) и Nitzschia closterium (тихопелагическая) - днем наблюдается депрессия фотосинтеза, а максимум интенсивности этого процесса приходится на утренние и послеполуденные часы (Бессемянова, 1959). Такое же течение фотосинтеза наблюдается в культурах северных пелагических видов Coscinosira polychorda и Coscinodiscus excentricus (Marshall, Огг, 1928; Jenkin, 1937). У бентосных форм интенсивность фотосинтеза на единицу биомассы значительно больше, чем у планктонных (Бессемянова, 1959). Это вполне закономерно" потому что бентосные диатомеи имеют крупные, интенсивно окрашенные пигментами хлоропласты, т. е. общее количество фотосинтезирующих пигментов у них значительно больше. Наблюдения показали, что фотосинтез протекает активнее у подвижных форм, чем у неподвижных и заметно активизируется в период деления диатомей (Talling, 1955). Фотосинтез не прекращается и при лунном свете, но в этих условиях кислорода выделяется в 10-15 раз меньше, чем днем. В верхнем горизонте водной толщи ночной фотосинтез составляет только 7-8% от суточного (Ивлев, Мухаревская, 1940; Subrahmanyan, 1960).
С глубиной сила света резко падает. Измерение на различных глубинах в зал. Пьюджет-Саунд (сев.-вост. часть Тихого океана) с помощью фотоэлектрической камеры Кунца показало, что интенсивность освещения (у поверхности воды принимаемая за 100%) на глубине 10 м падает до 9.6%, на глубине 20 м равна 4%, а на 35 м - 2.4%, практически на этой глубине совсем темно (Grein, in: Feldmann, 1938; Gessner, 1955-1959, I). Параллельно падению освещения укорачивается световой день. В океане на широтах 30-40° при наибольшей прозрачности воды на глубине 20 м длина летнего дня около И час., на 30 м - 5 час., на 40 м - всего 5 мин.
С глубиной не только убывают интенсивность освещения и световой период, но изменяется также качество света вследствие неодинакового поглощения лучей солнечного спектра разной длины световых волн. В табл. 3 приведены изменения поглощения световых лучей и цвета сумеречного освещения на разных глубинах.
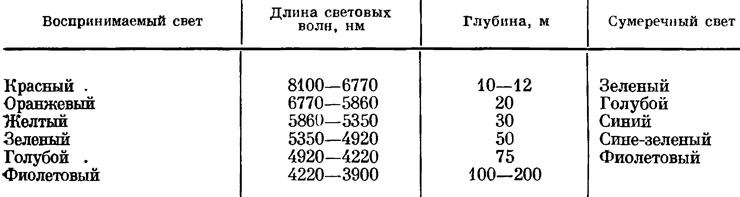
Таблица 3. Изменение поглощения световых лучей в море и цвета сумеречного освещения на разных глубинах
Эта таблица показывает, что поглощение света в воде моря обратно пропорционально длине световых волн, т. е. чем длиннее световые волны лучей спектра, тем быстрее они поглощаются водой. По мере поглощения световых лучей на соответственных глубинах изменяется цвет сумеречного освещения. То и другое лимитирует фотосинтез на глубинах. Убывание интенсивности различных лучей спектра на разных глубинах в море представлено в табл. 4.
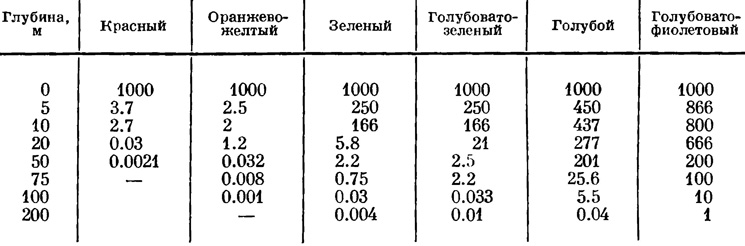
Таблица 4. Количественные соотношения световых лучей разной длины волн (в %) по глубинам в Средиземном море (по Grein, in: Feldmann, 1938 : 69)
Данные этой таблицы свидетельствуют, что некоторые морские бурые и красные водоросли еще могут вегетировать на глубине 75 м и, вероятно, глубже при условии очень высокой прозрачности воды. Как известно, прозрачность воды сильно варьирует не только в различных водоемах, но и в одном и том же водоеме. В пелагической области морей и океанов вода прозрачна до глубины от 40 до 160 м, а в морской сублиторали прозрачность воды падает до 20 м и ниже. Нижняя граница распространения водорослей определяется той интенсивностью света, при которой ассимиляция и дыхание взаимно уравновешиваются, т. е. когда достигается так называемая компенсационная точка (Marshall, Orr, 1928). Естественно, что компенсационная точка у водорослей зависит от прозрачности воды, от состава пигментов и ряда других факторов. В этом отношении имеются некоторые данные для морских водорослей-макрофитов, имеющих различные пигментные системы (Levring, 1966), но для диатомей таких сведений нет (табл. 5).
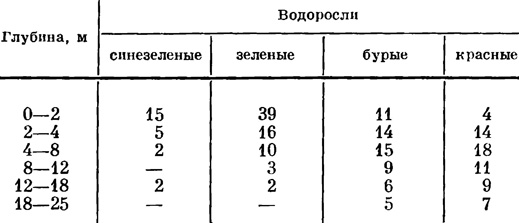
Таблица 5. Количественное распределение видов различных отделов водорослей-макрофитов по глубинам в Балтийском море (по Lakowitz, in: Gessner, 1957 : 320)
При равных условиях освещения компенсационная точка у водорослей разных отделов зависит от функции их пигментов. У синезеленых водорослей (имеющих пигменты: хлорофиллы а и b, β-каротин, кетокаротиноид, миксоксантофилл) компенсационная точка находится на глубине около 8 м, у зеленых (пигменты: хлорофиллы а и b, β-каротин, ксантофилл) - около 18 м, а у бурых и красных водорослей, имеющих кроме хлорофилла, каротина и ксантофилла дополнительные пигменты (у бурых фикоксантин, у красных - фикоэритрин и фикоциан), компенсационная точка опускается значительно ниже 30 м.
У некоторых видов диатомей сублиторали Черного моря компенсационная точка, по-видимому, может опускаться до глубины 35 м. Современная методика сбора сублиторальных диатомей не дает точного показателя условий обитания отдельных видов. На основании последних данных установлена общая закономерность расселения сублиторальных диатомей по глубинам. В условиях сублиторали Черного моря они обитают до глубины около 30 м (Прошкина-Лавренко, 1963а), в Средиземном море - до глубины 60 м (Aleem, 1951), что вполне естественно при прозрачности воды в этом море 60 м. Есть указания на обитание диатомей до 110 м (Smyth, 1955), до 200 м (Bougis, 1946) и до 7400 м (Wood, 1956), причем Вуд утверждает, что на этой глубине обнаружены живые диатомеи (обычно сублиторальные морские виды вместе с пресноводными!). Данные последних двух авторов недостоверны и требуют проверки.
Компенсационная точка у одного и того же вида диатомей непостоянна, она зависит от географической широты обитания вида, от сезона года, прозрачности воды и других факторов. Маршалл и Opp (Marshall, Orr, 1928) экспериментально установили, опуская культуру диатомей на раз-ные глубины в заливе (Лох Страйвен; Шотландия), что Coscinosira polychorda летом имеет компенсационную точку на глубине 20-30 м, а зимой близ поверхности воды. Подобные же результаты получены ими для Chaetoceros sp.
Бентосные диатомеи несомненно обладают хроматической адаптацией, этим объясняется способность многих из них обитать на некотором диапазоне глубин в условиях изменения спектрального света и его интенсивности; возможно, что они имеют различные расы (некоторые виды Amphora, Carrtpylodiscus, Diploneis, Navicula). Экспериментально установлено, что процесс приспособления к интенсивности освещения происходит довольно быстро. Так, например, пресноводная неподвижная планктонная диатомея Cyclotella meneghiniana приспосабливается к освещению от 3 тыс. до 30 тыс. лк в течение 24 час., она способна выносить значительно большую интенсивность света - до 60 тыс. лк и даже до 100 тыс. лк (Jorgensen, 1964а, 1964b). Фотосинтетический аппарат подвижных сублиторальных видов (Tropidoneis, Nitzschia) приспосабливается к условиям освещения на глубинах их обитания 1-3 м, где интенсивность света колеблется от 10 до 1% (Taylor, 1964). Вообще вопросу о хроматической адаптации у диатомовых водорослей посвящена большая литература (Talling, 1955, 1957а; Ryther, 1956; Ryther, Menzel, 1959; Steemann Nielsen, Hensen, 1959; Jørgensen, 1964a).
Планктонные диатомеи могут обитать значительно глубже сублиторальных, что связано преимущественно с большей прозрачностью воды в пелагиали. Известно, что в морях и океанах диатомовый планктон распространяется до глубины 100 м и более. В Черном море на глубине 75-100 м фитопланктон состоит из Thalassionema nitzschioides и нескольких видов Nitzschia, причем здесь они обитают в значительно большем количестве, чем в слое воды 0-50 м (Морозова-Водяницкая, 1948-1954). Многие виды Nitzschia, как известно, легко переходят от автотрофного питания к миксотрофному и гетеротрофному. По-видимому, этим же свойством обладают планктонные виды, обитающие в дисфотической и афотической зоне морей, они создают глубоководный теневой планктон. Впрочем, Стиман Нильсен и Хенсен (Steemann Nielsen, Hensen, 1959) рассматривают поверхностный фитопланктон как "световой" в условиях интенсивности радиации 600-1200 л к и как "теневой" в условиях низкой радиации: 200-450 лк. По мнению этих исследователей, зимний поверхностный фитопланктон в умеренном поясе является типичным "теневым". Однако зимний фитопланктон состоит из позднеосенних и ранневесенних видов, которые к "теневым" видам причислять нельзя. Следует признать, что проблема фитосинтеза у диатомовых водорослей пока находится на первоначальной стадии исследования, и по многим актуальным вопросам этой проблемы имеются только отрывочные и непроверенные данные.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'