Особенности систематической структуры лихенофлоры
С целью выяснения особенностей лихенофлоры МНР и определения ее положения в ряду близких флор было бы желательно провести сравнение ее состава с составом флор соседних с Монголией территорий. Но при попытке такого сравнительного анализа мы столкнулись с рядом трудностей. Как отмечал А. И. Толмачев (1941), сравнивать можно лишь равноценные флоры, которые могут быть сопоставимы по площади, разнообразию экологических условий и степени изученности. На современном этапе исследований в пределах умеренной Азии лихенофлора Монголии относится к числу наиболее планомерно и полно изученных. Сведения о составе лихенофлор соседних с Монголией территорий (Южной Сибири, Центральной Азии) далеко не полные. Наиболее значительные данные имеются только по флоре лишайников Горной Шории (Седельникова, 19776); для этого района известен список 322 видов лишайников. Для остальных же районов Южной Сибири публиковались лишь небольшие разрозненные списки видов лишайников (Еленкин, 1901а, 19016, 1901в, 1902, 19046, 1912; Zahlbruckner, 1911; Окснер, 1926; Рассадина, 1936, 1938, 1961; Савич, 1936; Будаева, 1976, и др.). Например, для Саян в пределах СССР известно немногим более 150 видов лишайников (Еленкин, 1901, 1902, 1903а, 1903б, 1904, 1906 - 1911, 1957; Савич, 1936, 1950; Рассадина, 1940, 1961; Дубровский, 1953). К сожалению, результаты более поздних исследований лишайников этого района, а также некоторых других территорий Южной Сибири нашли слабое отражение в печати. С. В. Кравчук (1981), например, указывает, что для Ойского хребта Западного Саяна ею установлено 322 вида лишайников, но при этом почти совсем не освещается систематический состав лихенофлоры, даже не называются ведущие семейства лишайников во флоре этого района. Слабо изучена и лихенофлора пограничного с Монголией Китая. По данным Цальбрукнера (Zahlbruckner, 1930), в 1930 г. общее число видов лишайников, известных для обширной территории этой страны, составило всего 717 (117 родов). К более поздним, наиболее крупным сводкам относится монография Магнуссона (Magnusson, 1940, 1944], посвященная лишайникам центральноазиатской части Китая. Магнуссон на основании обработки коллекций из 870 образцов, собранных участниками Китайско-Шведской экспедиции 1927 - 1935 гг. в Северо-Западном Китае, привел для провинции Ганьсу, Цинхай, Цайдам и Внутренней Монголии 216 видов лишайников, из них 142 вида были описаны как новые. Вряд ли этот список видов можно считать исчерпывающим и отражающим в полной мере лихенофлору Северо-Западного Китая, однако монография Магнуссона расширила представления в познании лишайников Центральной Азии, показав своеобразие лихенофлоры этого региона земного шара.
Как считает Р. В. Камелин (1973), отсутствие эталонных данных по соотношению видового количества различных групп в эталонных флорах крупных регионов Земли сильно обесценивает значимость статистических выкладок при изучении естественных флор. Это в большей степени относится к лишайникам. Можно с достаточной уверенностью сказать, что сравнимые статистические данные по лихенофлорам отдельных флористических областей земного шара найти довольно трудно. Однако в настоящее время в сравнительной флористике все большее значение приобретает анализ систематической структуры флор (состав и последовательность расположения ведущих по числу видов семейств) (Толмачев, 1970; Заки, Шмидт, 1972; Ребристая, Шмидт, 1972; Шмидт, 1974а, 19746, 1979, и др.). На примере анализа количественного соотношения различных систематических групп цветковых растений во флорах различных флористических областей земного шара показано, что в целом набор "ведущих семейств" оказывается весьма устойчивым (Толмачев, 1970). Очевидно, существует паритет, регламентирующий численность видов определенных систематических групп растений в пределах обширной флористической области. Причем это равновесие, выражающееся в устойчивости относительного числа видов определенных семейств, сочетается со значительной дифференциацией родового и видового составов флор, с богатым эндемизмом и т. п. При этом постоянство систематической структуры флор в пределах естественных флористических областей отмечается даже для различных по площади территорий. Кроме того, оно с достаточной определенностью выявляется даже при относительно неполной инвентаризации крупных (региональных) флор, причем систематическая структура крупных флор в пределах одной и той же флористической области более единообразна, чем структура мелких, например, конкретных флор в смысле А. И. Толмачева (Ребристая, Шмидт, 1972). Некоторые количественные соотношения, характеризующие систематическую структуру флоры, имеют, по-видимому, очень глубокие корни и отражают существенные ботанико-географические закономерности (Толмачев, 1970). Количественные соотношения между видами различных семейств закономерно изменяются в зависимости от широтного положения соответствующих пространств, они специфичны прежде всего для определенных флористических областей и несомненно должны отражать присущие им особенности флорогенеза и связи между ним и комплексом природных факторов. А. И. Толмачев (1970) отмечает, что для характеристики флор области как таковой определенные количественные соотношения между видами семейств цветковых растений выдвигаются на положение одного из существеннейших диагностических признаков.
Подобного сравнительного анализа систематической структуры лихенофлор отдельных флористических областей Голарктики до настоящего времени не проводилось, хотя несомненно было бы интересно проследить, наблюдается ли такое же постоянство в количественных соотношениях различных систематических групп лишайников в пределах отдельных ботанико-географических районов этого региона, как и в. случае цветковых растений. Грумман (Grummann, 1963) провел сравнение хорошо изученных лихенофлор ФРГ, ГДР, Скандинавии, Британских островов и Северной Америки. Хотя еще Декандоль (De Candolle, 1855) указывал на нецелесообразность сравнения данных как по узколокальным флорам (например, флора окрестностей города), так и флорам обширных стран (Франция, ФРГ, ГДР и т. п.) вследствие их разнородности, сравнительный анализ, проведенный Грумманом, представляет определенный интерес. На его основе можно сделать заключение, что, несмотря на различия в таксономической структуре этих лихенофлор, в численности видов в отдельных систематических группах и другие отличия, состав ведущих семейств в лихено-флорах этих районов довольно однороден. Основу европейских и североамериканских лихенофлор составляют лишайники чрезвычайно политипных семейств из числа накипных лишайников - Lecideaceae, Lecanoraceae, Verrucariaceae, занимающие в европейских лихенофлорах первые 3 места, а также других накипных лишайников семейств Caloplacaceae, Buelliaceae, Acarosporaceae, Pertusariaceae. Из листоватых и кустистых лишайников высокое положение в этих лихенофлорах занимают семейства Usneaceae, Parmeliaceae и Cladoniaceae. Некоторой спецификой отличается лихенофлора Северной Америки: в число ведущих семейств ее входит сем. Arthoniaceae. Видимо, лишайники вышеперечисленных семейств и составляют ядро лихенофлоры умеренной Голарктики. Однако их роль во флорах лишайников различных районов этого региона земного шара, по-видимому, неоднозначна.
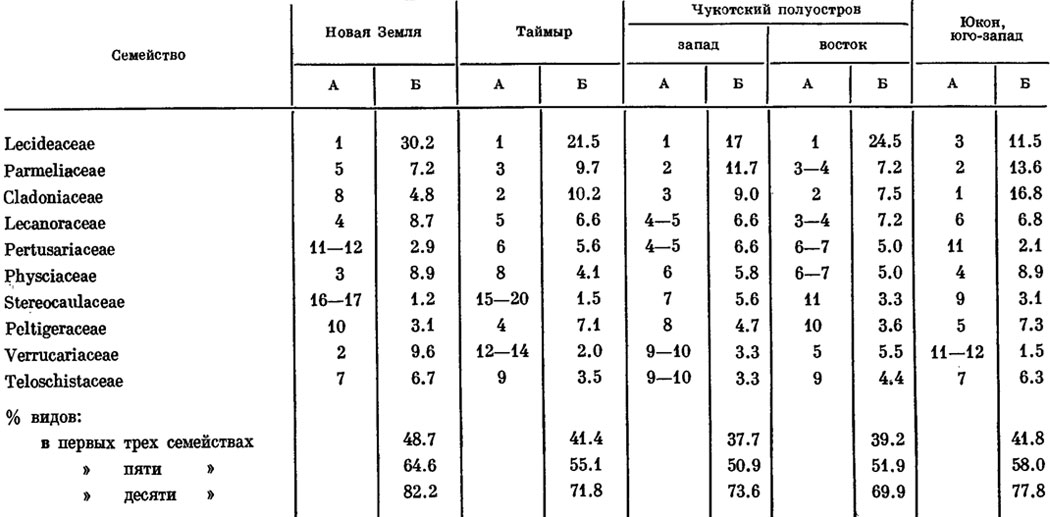
Некоторые особенности бореальных (арктических и лесных) лихенофлор умеренной Голарктики позволяют выявить данные, полученные советскими лихенологами (Голубкова и др., 1979; Макарова, 1979а, 19796; Андреев, 1980). Так, И. И. Макарова (1979а, 1979б), проанализировав данные ряда авторов, сравнила лихенофлоры различных районов Арктики - Новой Земли, п-ова Таймыр, западной и восточной частей Чукотского полуострова, юго-запада территории Юкон и некоторые другие (табл. 5)1. Она пришла к выводу, что в лихенофлорах этих районов состав десяти первых по числу видов семейств один и тот же: Lecideaceae, Parmeliaceae, Cladoniaceae, Lecanoraceae, Pertusariaceae, Verrucariaceae, Stereocaulaceae, Physciaceae, Peltigeraceae, Teloschistaceae. Причем, если их расположить в определенной последовательности по числу убывания видов, то места, занимаемые этими семействами в различных арктических лихенофлорах, будут довольно близкими, на основании чего И. И. Макарова делает вывод о целостности арктической лихенофлоры. Анализируя данные И. И. Макаровой (19796) и М. П. Андреева (1980), можно заключить, что к числу первых трех семейств, играющих видную роль во флорах лишайников различных районов Арктики, относятся семейства Lecideaceae, Parmeliaceae и Cladoniaceae. Особенностью арктических (тундровых) лихенофлор является то, что в их составе высокое положение занимают семейства Cladoniaceae, Peltigeraceae, Stereocaulaceae, Parmeliaceae (род Cetraria), т. е. систематические группы, объединяющие значительное число эпигейных и эпибриофитных лишайников, характерных для тундровых ценозов Арктики. При этом процент числа видов, охватываемых первыми десятью семействами в арктических лихенофлорах, оказался очень высоким - 70 - 80 (табл. 5).
1(В таблице 5 и далее объем семейств принимается по филогенетической системе лишайников, опубликованной в работе Пельта (Poelt, 1973): сем. Teloschistaсеае включает лишайники сем. Caloplacaceae; сем. Physciaceae - лишайники сем. Buelliaceae; сем. Verrucariaceae - Dermatocarpaceae; сем. Lichinaceae - виды сем. Pyrenopsidaceae.
Здесь и далее приводятся проценты от общего числа видов лишайников.)
Из сопоставления численных соотношений в систематической структуре флоры лишайников отдельных районов европейской части СССР и Сибири, расположенных в пределах лесной зоны (Седельникова, 1977а; Голубкова и др., 1979), видно, что спектры ведущих семейств лихенофлор этих районов характеризуются большим сходством, особенно спектры лихенофлор Эстонии, Московской области и Белоруссии (табл. 6). В составе этих лихенофлор в спектре ведущих семейств наиболее высокое положение занимают лишайники 5 семейств - Lecideaceae, Cladoniaceae, Lecanoraceae, Physciaceae, Parmeliaceae.
Таблица 5
Специфической особенностью лихенофлор лесных районов умеренной Голарктики является наличие в составе ведущих семейств систематических групп, объединяющих значительное количество эпифитных лишайников (например, сем. Usneaceae (роды Usnea, Bryoria) на 6-, 7-м месте). Видовой полиморфизм сем. Parmeliaceae во флорах этих районов также в основном проявляется за счет эпифитных лишайников рода Parmelia. Характерно наличие в составе спектра ведущих семейств лишайников сем. Caliciaceae (7-, 8-е места), среди представителей которого встречается большое число лесных бореальных и неморальных видов. Однако, как и в арктических флорах, в лесных лихенофлорах высокое положение занимают семейства Cladoniaceae (1 - 3-е места) и Peltigeraceae (9-, 10-е места), виды которых играют значительную роль в формировании напочвенных лесных ценозов.
К сожалению, при выявлении особенностей систематической структуры бореальных лесных лихенофлор приходится опираться главным образом на данные по европейским лихенофлорам, так как до настоящего времени, как уже отмечалось выше, отсутствуют достаточно полные сводки по флорам лишайников Сибири, за исключением Горной Шории. Сравнение состава ведущих семейств лихенофлоры Горной Шории и последовательности их расположения по числу убывания видов показывает их значительное сходство с лесными лихенофлорами европейских районов СССР (табл. 6). Некоторая специфика флористического спектра лишайников Горной Шории проявляется в высоком положении в составе лихенофлоры сем. Umbilicariaceae (9-е место), что можно объяснить горными условиями этого района.
Таблица 6
| Семейство | Эстонская ССР | Московская обл. | Белорусская ССР | Татарская АССР | Горная Шория | Бореальная монголия | ||||||
| А | Б | А | Б | А | Б | А | Б | А | Б | А | Б | |
| Lecideaceae | 1 | 15.0 | 1 | 19.6 | 1 | 17.3 | 5 | 10.3 | 1 | 10.5 | 1 | 11.2 |
| Cladoniaceae | 2 | 9.6 | 2 | 11.8 | 2 | 11.8 | 1 | 14.2 | 3 | 9.0 | 5 | 6.7 |
| Lecanoraceae | 3 | 8.0 | 3 | 9.7 | 3 | 7.6 | 2 | 11.6 | 5 | 6.2 | 4 | 6.9 |
| Physciaceae | 4 | 7.0 | 5 | 7.0 | 5 | 6.8 | 4 | 10.3 | 4 | 7.4 | 3 | 9.2 |
| Parmeliaceae | 5 | 6.7 | 4 | 6.7 | 4 | 7.6 | 3 | 11.2 | 2 | 9.6 | 2 | 9.5 |
| Teloschistaceae | 6 | 5.9 | 8 | 3.4 | 9 | 3.2 | 6 | 5.6 | 6 | 6.2 | 10 | 4.9 |
| Usneaceae | 7 | 4.7 | 6 | 5.2 | 6 | 5.2 | 7 | 5.2 | 8 | 4.9 | 7 | 5.6 |
| Caliciaceae | 8 | 4.4 | 7 | 4.4 | 8 | 4.1 | 13 | 1.7 | 11 | 2.6 | 22 | 1.1 |
| Verrucariaceae | 9 | 4.1 | 11 | 2.4 | 13 | 2.1 | 8 | 4.3 | 7 | 5.2 | 6 | 5.9 |
| Peltigeraceae | 10 | 2.1 | 9 | 3.1 | 11 | 2.4 | 10 | 3.4 | 10 | 3.1 | 11 | 4.3 |
| % видов: | ||||||||||||
| в первых трех семействах | 32.6 | 41.1 | 36.7 | 37.0 | 25.7 | 24.3 | ||||||
| " пяти " | 46.3 | 56.7 | 51.1 | 57.6 | 42.7 | 43.5 | ||||||
| " десяти " | 68.0 | 74.9 | 71.5 | 80.0 | 64.7 | 73.7 | ||||||
Далее попытаемся проследить особенности систематической структуры лихенофлор второго подцарства Голарктики - Древне-средиземноморского (в понимании А. Л. Тахтаджяна (1978)). При этом следует отметить, что в лихенологическом отношении территория этого региона изучена крайне неравномерно и выбор районов (табл. 7, 8) в основном диктуется наличием наиболее полных сводок по составу их лихенофлор. Лихенофлоры различных районов Средиземноморской области по составу ведущих семейств проявляют значительное сходство (табл. 7). Наиболее высокое положение в этих лихенофлорах занимают семейства Lecideaceae, Lecanoraceae, Physciaceae, Teloschistaceae, а также Verrucariaceae, за исключением лихенофлоры Марокко, где 5-е место занимает сем. Parmeliaceae. Специфической особенностью лихенофлор Средиземноморской области является наличие в составе ведущих семейств Ramalinaceae и Collemataceae. Как отмечал А. Н. Окснер (1956), Средиземноморская область, а также приокеанические районы Европы относятся к числу областей наиболее интенсивного видообразования рода Collema. По данным Сатала (Szatala, 1948), Средиземноморье является одним из центров развития рода Ramalina. Следует отметить, что в составе ведущих семейств этого региона отсутствуют семейства Cladoniaceae и Usneaceae. занимающие высокое положение в бореальных лихенофлорах (табл. 5 - 6). Однако появляется сем. Acarosporaceae, большинство видов которого - ксерофитные эпилитные и эпигейные лишайники. Как будет показано ниже, район Древнего Средиземья можно причислить к числу районов наиболее интенсивного видообразования ряда родов из акароспоровых лишайников.
Таблица 7
| Семейство | Марокко | О-в Корсика | О-в Крит | Израиль | ||||
| А | Б | А | Б | А | Б | А | Б | |
| Physciaceae | 1 | 8.7 | 2 | 9.1 | 4 | 9.1 | 3 | 14.0 |
| Lecanoraceae | 2 | 8.1 | 3 | 8.9 | 1 | 16.3 | 2 | 16.6 |
| Teloschistaceae | 3 | 7.9 | 5 | 7.0 | 3 | 13.4 | 1 | 17.2 |
| Lecideaceae | 4 | 6.8 | 1 | 13.1 | 2 | 15.0 | 4 | 12.1 |
| Parmeliaceae | 5 | 6.8 | 6 | 6.7 | 7 | 5.0 | 9 | 3.2 |
| Acarosporaceae | 6 | 4.1 | 12 | 2.4 | 8 | 5.0 | 10 | 3.2 |
| Collemataceae | 7 | 3.7 | 11 | 2.7 | 9 | 3.1 | 6 | 4.5 |
| Verrucariaceae | 8 | 3.5 | 4 | 8.2 | 5 | 7.2 | 5 | 6.4 |
| Ramalinaceae | 9 | 3.3 | 8 | 4.2 | 11 | 2.1 | 7 | 3.8 |
| Pertusariaceae | 10 | 3.3 | 7 | 4.7 | 9 | 3.4 | 11 | 2.5 |
| % видов: | ||||||||
| в первых трех семействах | 24.8 | 31.1 | 44.7 | 47.8 | ||||
| " пяти " | 38.4 | 46.3 | 61.0 | 66.3 | ||||
| " десяти " | 56.3 | 70.2 | 83.8 | 81.6 | ||||
Примечание. Данные для Марокко - по: Gattefosse, Werner, 1931; для о-ва Корсика - по: Werner, 1973; для о-ва Крит-по: Kleinig, 1966; для Израадя - по: Galun, 1970.
Набор ведущих семейств в лихенофлорах ряда горных районов Ирано-Туранской области Древнесредиземноморского подцарства проявляет большое сходство с таковым Средиземноморской области, особенно если сравнивать состав первых 4 - 5 семейств (табл. 8). Но для лихенофлор этих районов характерно высокое положение во флорах лишайников семейств Acarosporaceae (2-, 5-е места) и Aspiciliaceae (3-, 5-, 6-е места), что подчеркивает аридные черты лихенофлор. Специфично также для них наличие в числе ведущих семейств сем. Lichinaceae.
Таблица 8
| Семейство | Иран | Центральный Копетдаг | Горный Зеравшан | Восточный Памир | Северо-Западный Китай | Центральноазиатская Монголия | ||||||
| А | Б | А | Б | А | Б | А | Б | А | Б | А | Б | |
| Lecanoraceae | 1 | 14.5 | 1 | 18.1 | 1 | 19.3 | 1 | 23.3 | 3 | 12.0 | 4 | 8.4 |
| Teloschistaceae | 2 | 11.7 | 2 | 15.0 | 3? | 11.9? | 4 | 10.4 | 6 | 10.2 | 5 | 8.0 |
| Physciaceae | 3 | 11.3 | 3 | 13.8 | 4 | 10.4 | 7 | 8.0 | 7 | 8.8 | 3 | 10.1 |
| Lecideaceae | 4 | 10.1 | 4 | 10.6 | 2 | 12.4 | 5 | 9.8 | 4 | 11.1 | 2 | 11.2 |
| Acarosporaceae | 5 | 9.3 | 6 | 5.5 | 5 | 9.9 | 2 | 19.0 | 2 | 12.5 | 1 | 11.9 |
| Aspiciliaceae | 6 | 8.9 | 8 | 3.9 | 6 | 9.4 | 3 | 12.9 | 5 | 10.2 | 6 | 7.3 |
| Verrucariaceae | 7 | 8.1 | 5 | 5.5 | ? | ? | 6 | 8.0 | 1 | 13.1 | 7 | 7.0 |
| Parmeliaceae | 8 | 2.8 | 7 | 4.7 | ? | ? | 14 | 0.6 | 9 | 2.3 | 8 | 6.6 |
| Lichinaceae | 9 | 2.8 | - | - | 8 | 3.5 | 9 | 3.7 | 8 | 4.2 | 13 | 0.3 |
| Ramalinaceae | 10 | 2.4 | - | - | - | - | - | - | - | - | 26 | 0.2 |
| % видов: | ||||||||||||
| в первых трех семействах | 37.5 | 46.9 | 43.6 | 55.2 | 37.9 | 26.5 | ||||||
| " пяти " | 56.9 | 63.0 | 53.9 | 75.4 | 59.2 | 49.6 | ||||||
| " десяти " | 81.9 | 77.1 | 66.8 | 95.7 | 74.2 | 73.7 | ||||||
Таким образом, проведенный сравнительный анализ систематической структуры ряда лихенофлор Голарктики показывает, что, как и в случае цветковых растений, в пределах лихенофлор отдельных ботанико-географических областей наблюдается заметное постоянство в характерном наборе ведущих семейств и достаточно определенном порядковом расположении их по числу видов. По-видимому, систематическая структура лихенофлор естественных флористических районов может быть использована как один из важнейших показателей в сравнительно-флористических исследованиях. Характерно, что процент числа видов, охватываемых десятком ведущих семейств в сравниваемых лихенофлорах, оказался довольно высоким - от 56.3 до 83.8 (95.7). Как уже отмечалось ранее (Голубкова и др., 1979), этот показатель значительно превышает аналогичный показатель для флор высших растений, который составляет обычно 50 - 60% (Толмачев, 1974; Шмидт, 1974б).
А. И. Толмачев (1970) отмечал одну особенность состава флор: чем беднее состав флоры, тем меньшее число семейств содержит половину ее видового состава. Так, например, особой бедностью видового состава отличается пустынная лихенофлора Израиля (Galun, 1970) и флора лишайников холодных пустынь восточного Памира (Голубкова, 1973а, 1975), характеризующиеся простотой структуры, т. е. имеющие в своем составе ограниченный круг семейства. Семейства Lecanoraceae, Teloschistaceae, Physciaceae, занимающие первые 3 места в спектре ведущих семейств лихенофлоры Израиля, охватывают половину ее состава (47.8%), а 10 первых семейств - 81.6% состава лихенофлоры. Еще более показательна специфика лихенофлоры восточного Памира. Первые 3 семейства охватывают 55.2% ее состава, первые 5 - 75.4, первые 10 - 95.7%. Как полагает А. И. Толмачев (1970), определяющее значение здесь могут иметь специфические, экстремальные условия среды обитания, возможность приспособления к которым, очевидно, присуща разным группам растений в различной степени, в результате чего флора неизбежно формируется из представителей более или менее ограниченного круга семейств. Роль же экстремальных условий среды в развитии флор Средиземья, особенно его аридных частей, как считает А. И. Толмачев, проявляется в общем повышении роли ведущих семейств в сложении состава флор, сочетающемся с некоторым обеднением его. В этом получают отражение особенности флорогенеза этого региона, переживающего в геологически новейшее время значительное обогащение состава флор за счет видообразования в рамках определенных систематических групп, обусловленного в какой-то степени аридизацией условий существования (Толмачев, 1970).
Анализируя состав ведущих семейств флоры лишайников Монголии в свете проведенного сравнительного изучения спектров ведущих семейств отдельных регионов умеренной Голарктики, можно сделать следующее заключение. Состав ведущих семейств лихенофлоры Монголии типичен для лихенофлор умеренной Голарктики. Основу состава флоры лишайников Монголии составляют представители полиморфных семейств - Lecideaceae, Lecanoraceae, Verrucariaceae, Teloschistaceae, Physciaceae, входящих в комплекс ведущих семейств лишайников во флорах других районов этого региона. Высокое положение в составе флоры лишайников Монголии семейств Cladoniaceae, Usneaceae, Pelti- geraceae сближает ее с бореальными лихенофлорами Голарктики (табл. 3, 5, 6). В то же время высокое положение в составе лихенофлоры МНР семейств Acarosporaceae, Aspiciliaceae, а также семейств Collemataceae и Lichinaceae подчеркивает ее близость с флорами лишайников Древнего Средиземья, особенно его восточных районов. Таким образом, в флористическом спектре лишайников Монголии сочетаются черты, характерные, с одной стороны, для бореальных лихенофлор, с другой - для средиземных лихенофлор Голарктики, что указывает на гетерогенность состава флоры лишайников Монголии.
Подобная особенность прослеживается и в составе флоры сосудистых растений МНР (Грубов, Юнатов, 1952; Грубов, 1955, 1976). Флора сосудистых растений МНР, как подчеркивал В. И. Грубов (1976), не представляет собой нечто целостное и резко неоднородна. Это обусловлено прежде всего природными условиями Монголии, где северные таежные и горно-лесостепные районы резко отличаются ландшафтом, флорой и растительностью от южных - территорий пустынь, полупустынь и сухих степей. Однако различия в составе флоры этих двух частей страны объясняются не только современными физико-географическими условиями, но и особенностями исторического становления флор в составе двух резко различающихся между собой флористических областей Голарктики: с одной стороны - Бореального, с другой - Древнесредиземноморского подцарств.
Как указывалось выше, приблизительно в средней части МНР по главному хребту горной страны Хангай и р. Керулен проходит линия мирового водораздела между бассейнами Северного Ледовитого и Тихого океанов и территорией внутреннего стока Центральной Азии, рассекающая эту страну как бы на две части (Грубов, 1976). Северная часть является окраинными поднятиями Южной Сибири, продолжением Саяно-Южнобайкайльской горной системы. Для этих районов Монголии характерна горная кедровая и кедрово-лиственничная тайга, переходящая к югу в осветленные лиственничники. Южная часть страны - степи, полупустыни и пустыни Центральной Азии. Согласно ботанико-географическому районированию (Тахтаджян, 1978), эти два района относятся к различным флористическим областям: северная часть МНР - к Циркумбореальной области и Восточноазиатской области (самые северо-восточные окраины МНР) Бореального подцарства Голарктики, южная - к Центральноазиатской подобласти Ирано- Туранской области Древнесредиземноморского подцарства.
Если сравнить флористические спектры бореальной и центральноазиатской частей МНР, то они будут различны. Это различие особенно проявляется в составе первых 5 семейств (табл. 9). Как можно видеть, набор первых 5 семейств бореальной части МНР совпадает с составом первых 5 семейств в лихенофлорах лесных районов бореального подцарства Голарктики (табл. 6, 9). Высокое положение во флоре лишайников этой части Монголии семейств Parmeliaceae (2-е место) и Cladoniaceae (3-е место) характеризует ее принадлежность к лихенофлорам бореального типа. Как можно видеть из табл. 9, в составе лихенофлоры этой части МНР значительную роль играют также семейства Usneaceae (6-е место) и Peltigeraceae (9-е место), что характерно, как уже указывалось выше, также для бореальных лесных флор Голарктики. Лишайники этих полиморфных семейств широко участвуют в растительном покрове северной части МНР (Прихубсугулье, Хангай, Хэнтэй). Представители семейств Cladoniaceae, Usneaceae (роды Usnea, Bryoria, Evernia), а также сем. Parmeliaceae (роды Parmelia, Cetraria) в условиях Монголии встречаются главным образом в лесных и высокогорных растительных сообществах горно-таежных, лесостепных и высокогорных районов северной части страны. В напочвенном покрове лиственничных, кедрово-лиственничных и сосновых лесов Монголии велика роль лишайников семейств Cladoniaceae и Peltigeraceae. Значительное участие в составе кобрезиевников и высокогорных тундр принимают лишайники семейств Cladoniaceae и Parmeliaceae (род. Cetraria), а также сем. Usneaceae (роды Cornicularia, Evernia, Dactylina). Специфические черты флористическому спектру лихенофлоры бореальной части МНР придает наличие в составе ведущих семейств сем. Acarosporaceae (7-е место), что свидетельствует о ксерофитных чертах лихенофлоры.
Таблица 9
| Семейство | Число видов | % от общего числа видов | Семейство | Число видов | % от общего числа видов |
| Бореальная часть Монголии | Центральноазиатская часть Монголии | ||||
| Lecideaceae | 68 | 11.2 | Acarosporaceae | 51 | 11.9 |
| Parmeliaceae | 58 | 9.5 | Lecideaceae | 48 | 11.2 |
| Physciaceae | 56 | 9.2 | Physciaceae | 43 | 10.1 |
| Lecanoraceae | 42 | 6.9 | Lecanoraceae | 36 | 8.4 |
| Cladoniaceae | 41 | 6.7 | Teloschistaceae | 34 | 8.0 |
| Verrucariaceae | 36 | 5.9 | Aspiciliaceae | 31 | 7.3 |
| Usneaceae | 34 | 5.6 | Verrucariaceae | 30 | 7.0 |
| Aspiciliaceae | 33 | 5.4 | Parmeliaceae | 28 | 6.6 |
| Acarosporaceae | 32 | 5.3 | Usneaceae | 22 | 5.2 |
| Teloschistaceae | 30 | 4.9 | Peltigeraceae | 16 | 3.7 |
| Peltigeraceae | 26 | 4.3 | Collemataceae | 13 | 3.0 |
| Collemataceae | 19 | 3.1 | Lichinaceae | 13 | 3.0 |
| Umbilicariaceae | 17 | 2.8 | Cladoniaceae | 11 | 2.5 |
| Всего | 492 | 80.8 | 365 | 87.9 | |
Анализ состава ведущих семейств флоры лишайников центральноазиатской части Монголии показывает сходство состава первых 5 семейств с другими лихенофлорами Ирано-Туранской области Древнесредиземноморского подцарства (табл. 8, 9), что характеризует ее принадлежность к аридным флорам средиземного типа. Ксерофитные черты лихенофлоры этой части МНР подчеркиваются очень высоким положением во флоре сем. Acarosporaceae (1-е место), а также сем. Aspiciliaceae (9-е место). Ксерофитные лишайники этих семейств - характерные представители растительных сообществ пустынно-степных районов Монголии. Большой удельный вес во флоре лишайников центрально- азиатских районов Монголии имеют и другие группы накипных лишайников, особенно семейств Lecanoraceae (виды секции Placodium рода Lecanora), Lecideaceae (роды Lecidea, Lecidella, Psora), Verrucariaceae (роды Endopyrenium, Endocarpon), Teloschistaceae (род Caloplaca). Большинство из них - эпилитные лишайники, они широко распространены на поверхности скал, валунов, галечно-щебнистого панциря, чем достигается их заметная роль в составе петрофитных растительных сообществ горных склонов, а также в лишайниковых ценозах горных, сухих и пустынных степей. Из листоватых лишайников значительный удельный вес во флоре и растительности пустынно-степных районов Монголии принадлежит представителям сем. Physciaceae (2-е место). Более скромная роль отводится здесь лишайникам сем. Parmeliaceae (6-е место), хотя отдельные виды рода Parmelia - P. vagans Nyl., P. taractica Krempelh., P. tominii Oxn., P. conspersa (Ehrh.) Ach. - распространены в этих районах Монголии довольно широко. Как и в других лихенофлорах территории Древнесредиземноморского подцарства, значительное участие во флоре пустынно-степной части Монголии принимают лишайники сем. Lichinaceae (виды родов Gonohymenia, Psorotichia, Peccania), хотя это семейство не вошло в число первых десяти ведущих семейств флоры лишайников центральноазиатской Монголии. Однако, как можно видеть из табл. 9, в состав ведущих семейств лихенофлоры центрально-азиатской части МНР входят также семейства Usneaceae и Peltigeraceae. В этой части Монголии представители этих семейств обитают исключительно в горных районах, в высокогорных степях Монгольского Алтая, реже Гобийского Алтая, в лиственничных лесах Монгольского Алтая и совершенно не встречаются в плакорных растительных сообществах пустынно-степных районов.
Следует подчеркнуть, что крупные горные поднятия на территории Монголии в значительной степени затушевывают различия между лихенофлорами бореальной и центральноазиатской частей МНР как в количественном отношении, так и в составе лихенофлор. При этом необходимо отметить, что видовое разнообразие лихенофлоры центральноазиатских районов Монголии (табл. 2) в значительной степени обусловлено богатством флоры лишайников горных районов, особенно Монгольского Алтая, в то время как лихенофлора плакорных пространств пустынно-степных районов Монголии бедна и однообразна в видовом отношении и носит все черты ксерических флор аридных областей земного шара (Голубкова, Цогт, 1974а, 19746).
Флора лишайников бореальной части МНР несравненно богаче лихенофлоры центральноазиатской части Монголии по численности видов, родов и семейств, хотя в показателях систематического многообразия (или пропорциях флоры - отношения в/с, р/с, в/р) различия выражены довольно слабо (табл. 2). По сравнению с лихенофлорами бореальных районов Монголии во флоре центральазиатской части этой страны отсутствуют представители 9 семейств - Lecanactidaceae, Sphaerophoraceae, Ostropaceae, Gyalectaceae, Nephromatacea, Lobariaceae, Coccarpiaceae, Baeo- mycetaceae, Agyriaceae, в то время как в лихенофлоре бореальной Монголии - лишь одно семейство - Microglaenaceae. Еще более наглядна разница в родовом разнообразии этих двух районов МНР. Во флоре лишайников центральноазиатской части Монголии отсутствуют представители 38 родов, представленные в лихенофлоре бореальных районов этой страны, - Lecanactis, Leptorhaphis, Arthopyrenia, Polyblastiopsis, Microthelia, Polycoccum, Polyblastia, Chaenothecopsis, Calicium, Stenocybe, Sphaerophorus, Conotrema, Coenogonium, Porocyphus, Synalissa, Anema, Lempholemma, Phylliscum, Polychidium, Moelleropsis, Nephroma, Lobaria, Pseudocyphellaria, Sticta, Parmeliella, Trpelia, Huilia, Catinaria, Mycoblastus, Platismatia, Neuropogon, Cladina, Icmadophila, Baeomyces, Strangospora, Heterodermia, Scoliciosporum, Xylographa.
Это объясняется прежде всего тем, что обитание многих из них связано на территории МНР с лесными ценозами, например представителей родов Nephroma, Sticta, Lobaria, Cladonia, Leptorhaphis, Xylographa, Icmadophila и др., других - с высокогорным поясом, где они встречаются в горных тундрах или на кобрезиевых пустошах, например лишайники родов Cladina, Sphaerophorus. Представители некоторых из них являются синтопными лишайниками и приурочены к обитанию на карбонатном каменистом субстрате, где, как известно, поселяются лишайники несколько специфического состава (Rasanen, 1927; Седельникова, 1980). Так, на карбонатных породах только в районе Прихубсугулья были найдены лишайники родов Synalissa и Anema. И в то же время во флоре бореальной части Монголии не обнаружены представители лишь 5 родов лишайников - Glypholecia, Microglaena, Hymenelia, Psorotichia, Amphoridium, встречающиеся в центральноазиатских районах этой страны.
На примере ведущих семейств (табл. 9) можно проследить обеднение видового состава целого ряда систематических групп во флоре лишайников центральноазиатской Монголии по сравнению с лихенофлорой бореальной части. Особенно это ярко проявляется в группах листоватых и кустистых лишайников. Например, сем. Parmeliaceae во флоре бореальной Монголии насчитывает 58 видов, в центральноазиатской Монголии - 28, сем. Usneaceae соответственно 34 и 22, сем. Cladoniaceae - 41 и 11 видов. Лишь одно сем. Acarosporaceae отличается большим видовым разнообразием в лихенофлоре центральноазиатской части Монголии (51 вид) по сравнению с флорой бореальных районов страны (32 вида).
Более наглядно обеднение видового состава некоторых систематических групп лишайников во флоре южных районов Монголии проявляется на примере родов. Особенно это хорошо заметно у представителей кустистых и листоватых лишайников. Так, в бореальной Монголии род Parmelia насчитывает 36 видов, а в центральиоазиатской - 17, род Cladonia соответственно 37 и 11, род Usnea - 12 и 6, Hypogymnia - 11 и 4, Cetraria - 14 и 8, Вгуоria - 11 и 8 видов. При этом необходимо подчеркнуть, что представители этих родов не характерны для лихенофлоры центрально-азиатской части МНР и встречаются здесь почти исключительно в горах Монгольского Алтая, в лесных и высокогорных растительных сообществах, и отсутствуют в плакорных растительных сообществах этих районов. Исключение составляют лишь некоторые виды рода Cladonia (С. symphycarpa (Ach.) Fr., С. pocillum (Ach.) О.-J. Rich., С. pyxidata (L.) Hoffm.), которые изредка встречаются в степных ценозах этой части МНР, а также некоторые виды рода Parmelia, довольно широко распространенные в пустынно-степных районах Монголии. У представителей некоторых родов накипных лишайников в центральиоазиатской части МНР также наблюдается обеднение видового состава, например в родах Lecidea, Rinodina, Biatora. У целого ряда родов - Aspicilia, Lecanora, Buellia, Rhizocarpon, Physcia, Candelariella - видовое разнообразие приблизительно, одинаковое в обеих частях МНР. Род Acarosроrа проявляет значительное видовое богатство во флоре центральноазиатских районов Монголии (49 видов) по сравнению с лихенофлорой бореальной Монголии (29 видов). В целом, по нашим данным, лихенофлора хангайской (бореальной) части МНР богаче флоры лишайников гобийской (центральноазиатской) части Монголии на 181 вид.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'