Органы размножения лишайников
Лишайники размножаются либо спорами, которые образует микобионт половым или бесполым путем, либо вегетативно - фрагментами слоевища, соредиями и изидиями.
Половое размножение. При половом размножении на слоевищах лишайников в результате полового процесса формируются половые спороношенрш в виде плодовых тел. Среди плодовых тел у лишайников различают апотеции, перитеции и гастеротеции. Большинство лишайников, свыше 250 родов, формируют открытые плодовые тела в виде апотециев - дисковидных образований (рис. 305, 1, 2; 306). Около 70 родов лишайников имеют плодовые тела в форме перитеция - закрытого плодового тела, имеющего вид маленького кувшина с отверстием наверху (рис. 307). Небольшое количество лишайников, в основном представители семейства графидовых, образуют узкие плодовые тела удлиненной формы, которые называют гастеротециями (рис. 305, 4). В апотециях, перитециях и гастеротециях споры развиваются внутри сумок - особых мешковидных образований (рис. 308). Лишайники, формирующие споры в сумках, объединяются в большую группу сумчатых лишайников (Ascolichenes). Они произошли от грибов класса аскомицетов и представляют основную эволюционную линию развития лишайников.
У небольшой группы лишайников споры образуются не внутри сумок, а экзогенно, на вершине удлиненно-булавовидных гиф - базидий (рис. 303, 2). На верхушке базидий имеются четыре отростка - стеригмы, на концах которых развиваются четыре споры. Лишайники с таким образованием спор объединяются в группу базидиальных лишайников (Ваsidiolichenes).
Несмотря на то что базидиальные лишайники насчитывают всего 20 видов, это совершенно самостоятельная эволюционная линия развития лишайников, берущая свое начало от грибов класса базидиомицетов. Базидиальные лишайники в основном тропические растения. По внешнему облику они напоминают тонкослойные трутовые грибы - разрушители древесины (рис. 303, 1). На нижней стороне их слоевищ, имеющих вид тонких полукруглых пластинок, образуется открытый плодущий слой, который состоит из базидий и бесплодных удлиненных гиф - парафиз (рис. 303, 2б). Совсем недавно базидиальные лишайники были обнаружены и в Европе. Однако плодовые тела базидиальных лишайников, произрастающих в горах Средней Европы и в северных тундрах, иного строения. Они имеют вид пальцевидных или коралловидных выростов либо напоминают по форме маленький шляпочный гриб (рис. 303, 3, 4, 5).

Рис. 303. Базидиальные лишайники: 1 - тропический лишайник Cora pavonia; 2-4 - базидиальные лишайники с плодовыми телами в форме шляпочного гриба (2,3 - Omphalina luteolilacina, внешний вид и поперечный разрез через плодовое тело; 4 - Omphalina ericetorum): а - базидии, б - парафизы, в - споры; 5 - роговидные плодовые тела базидиального лишайника Clavulinopsis septentrionalis
Половой процесс и развитие плодовых тел у лишайников, особенно базидиальных, изучены недостаточно. Эти процессы имеют много общих черт с аналогичными процессами у свободноживущих грибов, хотя и отличаются целым рядом особенностей. Было замечено, что развитие плодового тела в различных семействах лишайников протекает неодинаково и изменяется от семейства к семейству. На основе онтогенеза плодовых тел сумчатые лишайники делят на две группы: асколокулярные и аскогимениальные. Существуют и промежуточные формы (семейство Arthoniaceae), которые объединяют черты этих двух основных типов развития плодовых тел.
Развитие плодового тела асколокулярных лишайников начинается с возникновения особой ткани - стромы, состоящей из переплетения грибных гиф, а уже затем в строме закладывается архикарп - женский половой орган лишайника. Позднее в строме возникают особые камеры, называемые локулами, внутри которых формируются сумки со спорами. Грибная ткань между локулами частично отмирает, и от нее остаются лишь отдельные вертикально расположенные гифы, срастающиеся своими вершинами, их называют парафизоидами.
Этот тип онтогенеза плодовых тел наиболее характерен для свободно живущих аскомицетных грибов и среди лишайников встречается довольно редко, лишь у представителей двух семейств: артопирениевых (Arthopyreniaceae) и микопоровых (Мусорогасеае).
Подавляющее большинство сумчатых лишайников принадлежит к группе аскогимениальных. Это филогенетически очень древняя группа, представители которой благодаря их симбиотическому образу жизни рано отделились от грибов и развивались далее самостоятельно и независимо от них. У аскогимениальных лишайников развитие плодовых тел начинается с закладки архикарпа непосредственно среди вегетативных гиф в верхней части сердцевинного слоя, на границе с зоной водорослей или, реже, в самой зоне водорослей. В дальнейшем во взрослом плодовом теле аскогимениальных лишайников всегда формируется настоящий гимениальный слой, образованный вертикально стоящими сумками со спорами и развивающимися между ними парафизами - бесплодными вытянутыми гифами, свободными на верхнем конце (рис. 305, 5, 6).
Женский половой орган лишайников - архикарп - состоит из двух частей (рис. 304, 3, 4). Нижняя часть носит название аскогона и представляет собой спирально закрученную гифу, более толстую по сравнению с другими гифами и состоящую из 10-12 одно- или многоядерных клеток. От аскогона вверх отходит трихогина - тоненькая вытянутая гифа, которая проходит через зону водорослей и коровой слой и выходит на поверхность слоевища, возвышаясь над ней своей липкой верхушкой.
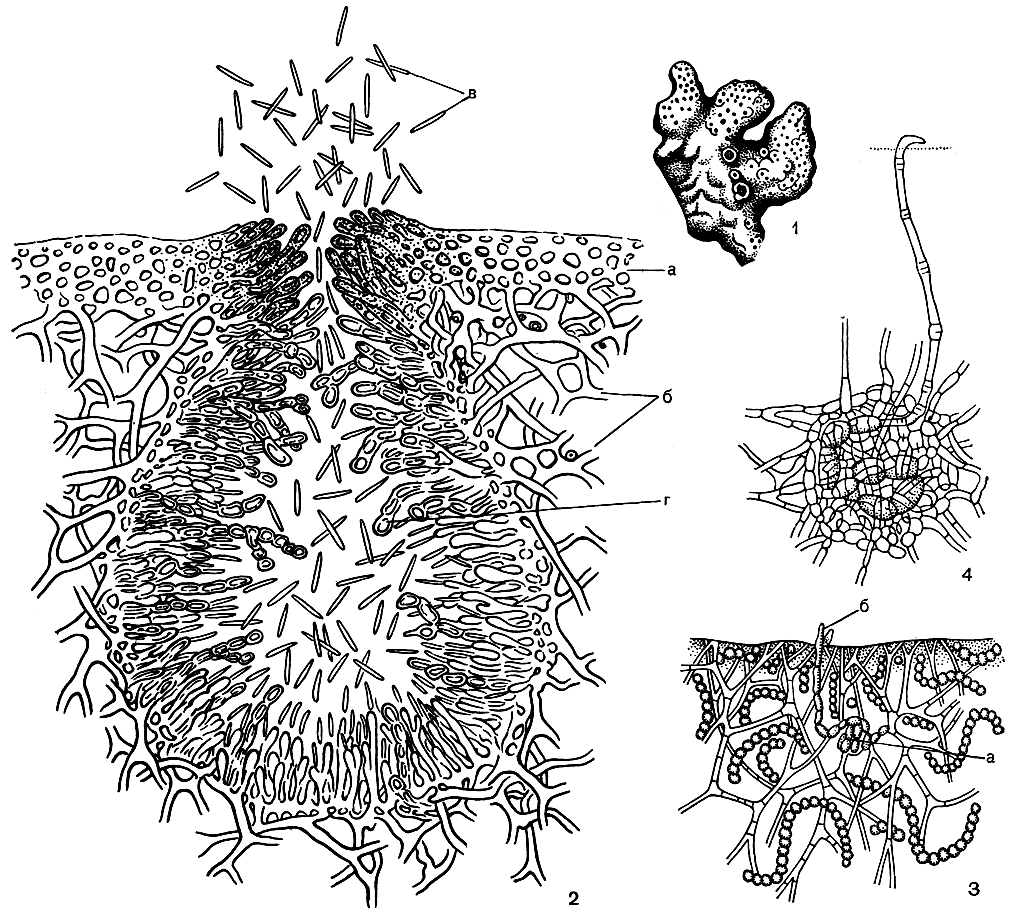
Рис. 304. Пикнидии и архикарп у лишайников: 1 - лопасть листоватого лишайника гипогимнии с пикнидиями, заметными на поверхности в виде черных точек; 2 - поперечный разрез через слоевище с пикнидией (а - коровой слой слоевища, б - сердцевина, в - пикноконидии, г - конидиеносцы); 3 - поперечный разрез через слоевище коллемы, в центре - многоклеточный архикарп (а - аскогон, б - трихогина, высовывающаяся над поверхностью слоевища); 4 - аскогон и трихогина после копуляции с пикноконидией
Как происходит оплодотворение архикарпа и вообще имеет ли место этот процесс у лишайников, до сих пор не выяснено. Правда, некоторым ученым удалось наблюдать, как к клейкой верхушке трихогины прилипали маленькие бесцветные тельца - пикноконидии. Они быстро теряли свое цитоплазматическое содержимое, и на вершине трихогины оставалась лишь их пустая оболочка. Можно было предположить, что содержимое этих телец переходит в трихогину. Это позволило считать пикноконидии мужскими половыми клетками, оплодотворяющими архикарп.
Что же представляют собой пикноконидии? Это бесцветные, очень маленькие клеточки (длиной 2-8 мкм, шириной 0,5-1 мкм) различной формы: палочковидные, овальные, игловидные, прямые и изогнутые. Пикноконидии образуются в пикнидиях - особых замкнутых вместилищах шаровидной или овальной формы с маленьким отверстием наверху (рис. 304, 2, 2). Пикнидии нередко можно наблюдать на верхней поверхности слоевища лишайников или на кончиках и по краям лопастей, где они бывают заметны в виде черных, реже красных точек - это видны лишь выводные отверстия пикнидиев, в то время как сами они целиком погружены в слоевище. Стенки пикнидия состоят из нескольких слоев клеток, и от них перпендикулярно внутрь отходят конидиеносцы - тонкие гифы, поделенные на маленькие клетки. На конидиеносцах образуются многочисленные пикноконидии. При увлажнении слоевища маленькие пикноконидии в огромной массе выплывают на поверхность слоевища лишайника и некоторые из них, по-видимому, прилипают к высунувшимся клейким верхушкам трихогин. Но действительно ли пикноконидии являются мужскими половыми клетками, оплодотворяющими архикарп, пока с уверенностью сказать нельзя. В настоящее время еще нет цитологических данных, показывающих проникновение содержимого пикноконидии в трихогину. Многие ученые решительно отрицают, что пикноконидии - это мужские половые клетки и, наоборот, считают их органами бесполого размножения лишайников. В некоторых случаях удавалось наблюдать, как пикноконидии в благоприятных условиях сами прорастали в мицелий, который давал начало образованию слоевища лишайников.
Считается, что для лишайников характерна редукция, утрата процесса оплодотворения. Возможно, что во многих случаях процесса оплодотворения вовсе не происходит и плодовое тело формируется из неоплодотворенного аскогона. Причем лишь у слизистых лишайников семейства коллемовых развитие плодового тела происходит из одного архикарпа, как и у свободноживущих грибов. У всех остальных лишайников плодовое тело обычно формируется из многих архикарпов и окружающих их гиф. Из нижней части архикарпа - аскогона - развиваются аскогенные гифы, из которых в дальнейшем формируются сумки. Их развитие начинается с разрастания аскогона и попарной группировки ядер в его клетках. В результате этого процесса возникают клетки, каждая из которых содержит по два ядра. Эти двухъядерные клетки - материнские клетки сумок - постепенно начинают разрастаться, и из них формируются сумки. Далее, по мере их созревания, внутри сумок происходит слияние двух ядер в одно диплоидное, копуляционное ядро. Через некоторое время наблюдается последовательное тройное деление этого ядра, в результате чего в сумке образуется уже 8 ядер, каждое из которых является ядром будущей споры. Постепенно вокруг каждого ядра обособляется часть цитоплазмы сумки, а затем эта обособленная часть цитоплазмы с заключенным внутри ее ядром покрывается оболочкой. Так в сумке образуется 8 спор.
Развивающийся аскогон окружается, как футляром, сплетением мелкоклеточных гиф, из которых формируются отдельные части плодового тела. Эти ткани называют генеративными. Гифы генеративных тканей отличаются от гиф остального слоевища тонкими клеточными стенками, формой клеток и их богатым содержимым, причем структура генеративных тканей у различных лишайников сильно варьирует, так же как и процесс дальнейшего формирования плодовых тел.
Развитие и созревание плодового тела у лишайников - очень медленный процесс, который длится 4-10 лет. Сформировавшееся плодовое тело тоже является многолетним, способным в течение ряда лет продуцировать споры.
Большинство лишайников образуют плодовые тела открытого типа в форме апотециев. Эти округлые дисковидные образования, напоминающие по виду маленькие блюдца, очень часто можно наблюдать на верхней поверхности слоевища накипных и листоватых лишайников или на концах лопастей кустистых слоевищ (табл. 43, 44, 53). Обычно их диаметр не превышает 1-2 мм, но у некоторых листоватых и кустистых лишайников он может достигать 1-3 см, а у накипных - всего 0,1-0,5 мм. По форме апотеции чаще всего округлые, реже овальные; когда же несколько апотециев развивается рядом друг с другом, они нередко становятся угловатыми. У семейства артониевых (Arthoniaceae) они имеют форму звезд или неправильных пятен. Обычно апотеции образуются на верхней поверхности слоевища лишайников, плотно прирастая к ней своей нижней частью или возвышаясь над ней на тонкой маленькой или довольно длинной ножке (рис. 305, 3). У некоторых лишайников они бывают целиком погружены в слоевище, например у солорины (табл. 42, 4). Только у двух родов лишайников - нефрома (Nephroma) и нефромопсис (Nephromopsis)- апотеции образуются не на верхней, а на нижней стороне лопастей (табл. 49, 5).
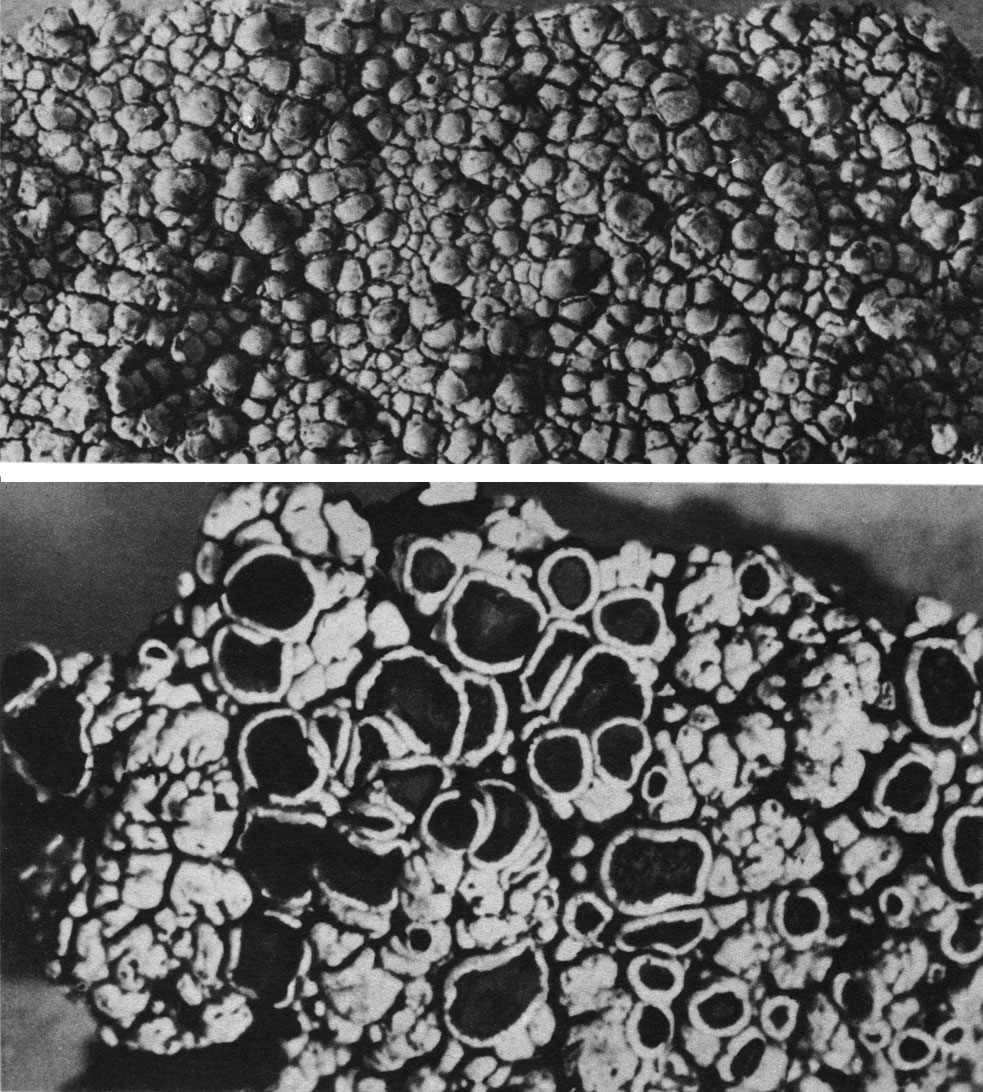
Таблица 43. Наскальные лишайники с ареолированными слоевищами:
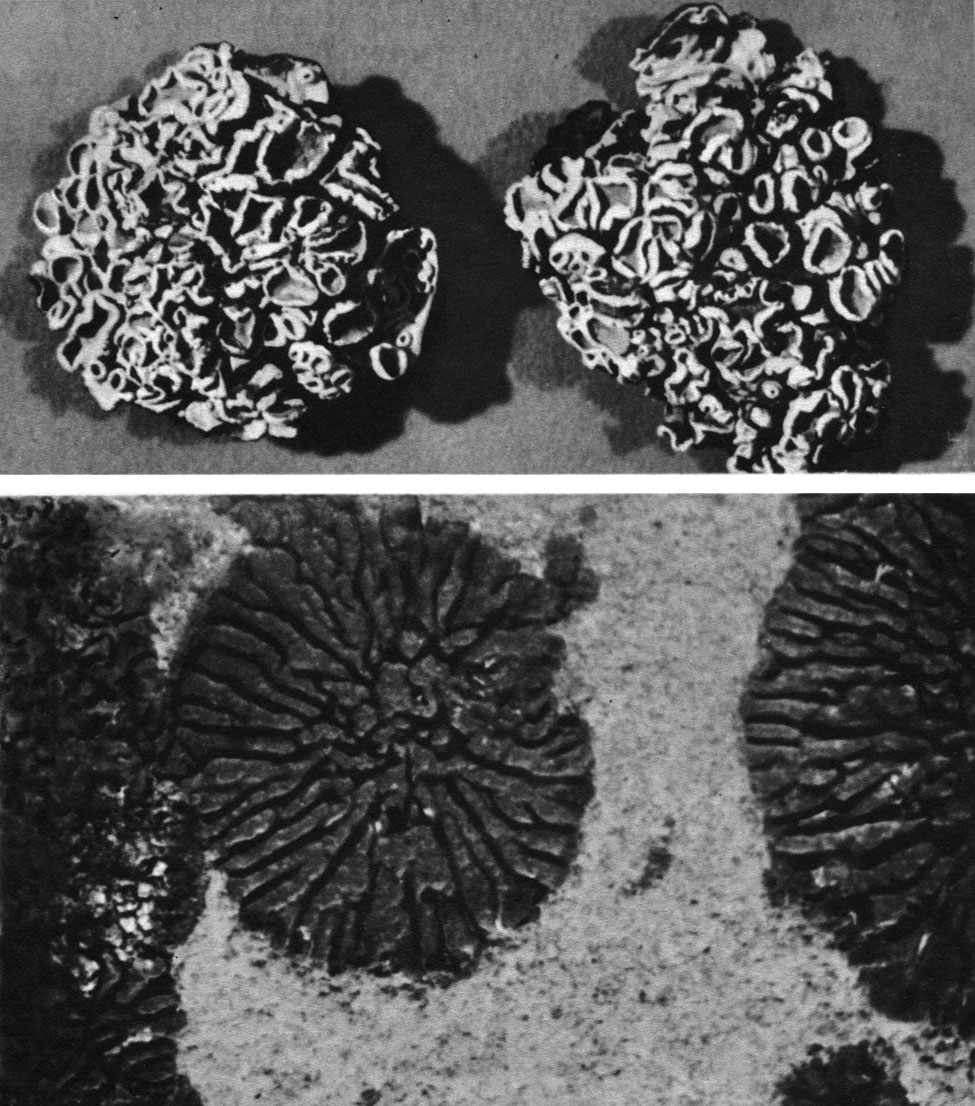
Таблица 44. Розетковые накипные слоевища: вверху - арктоальпийский наскальный лишайник Placolecanora peltata, внешний вид слоевища с апотециями; внизу - арктический наскальный лишайник Acarospora molybdina
В апотециях можно различить центральную часть - плоский диск и периферическую - округлый выпуклый валик, окружающий диск.
Плодущей частью апотеция является диск. На его поверхности развивается плодущий слой, который носит название гимениального. Он образован вертикально стоящими сумками со спорами и расположенными между ними бесплодными гифами - парафизами (рис. 305, 2, 5, 6). Парафизы - тонкие, нитевидные гифы (обычно толщиной 1,5-2 мкм), простые или разветвленные, со свободными верхними концами. Они служат для защиты сумок со спорами. По высоте они обычно несколько превосходят сумки, а их свободный верхний конец бывает булавовидно утолщен и окрашен в зеленоватый, синеватый, коричневатый, желтоватый и другие цвета. Эти окрашенные утолщенные вершинки, плотно прилегая друг к другу, образуют над сумками в верхней части гимениального слоя защитный слой, который называют эпитецием. От окраски эпитеция зависит окраска диска апотеция: он может быть коричневым, черным, желтым, оливковым, красным, оранжевым, розовым и др. (табл. 42, 47, 48, 49). Под гимениальным слоем расположен гипотеций - слой, состоящий из плотно переплетенных гиф. Тонкое цитологическое исследование позволяет различить в нем основания парафиз и аскогенный аппарат, из которого по мере созревания вырастают сумки.
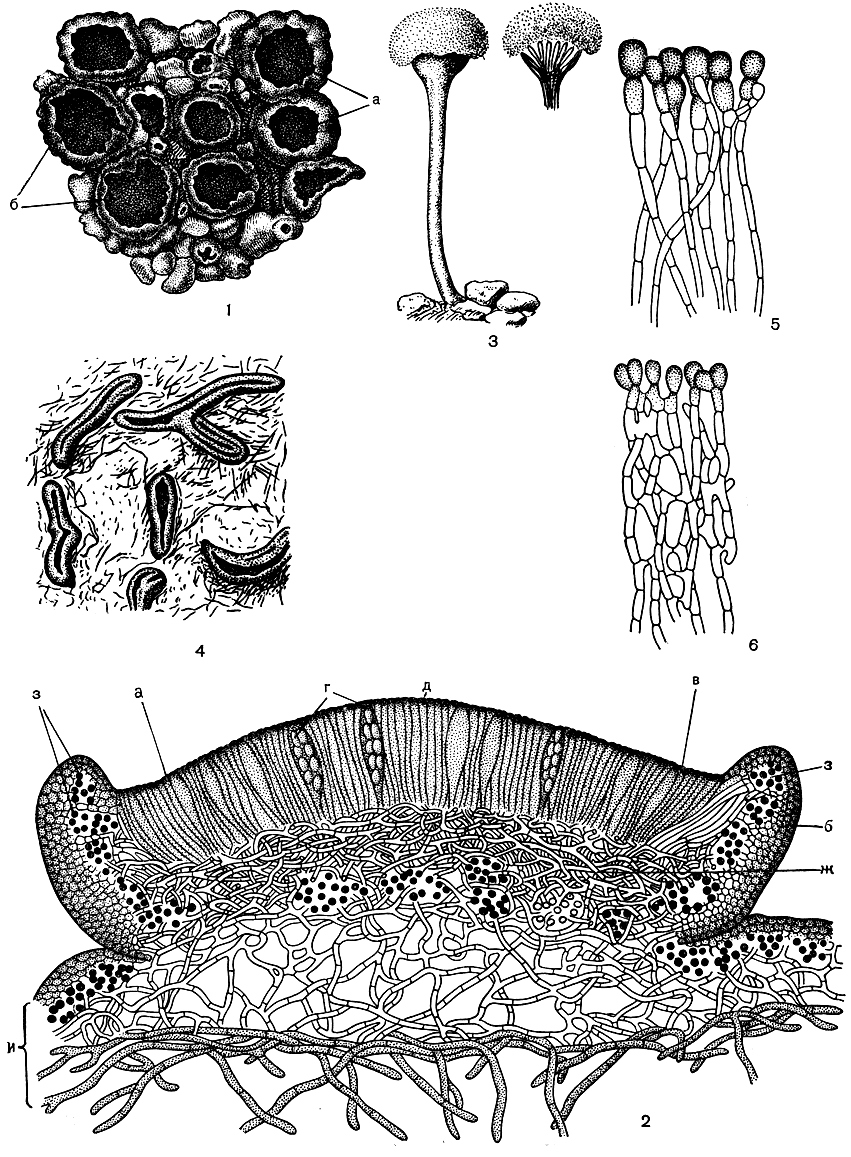
Рис. 305. Леканоровые апотеции и гастеротеции: 1 - внешний вид анотециев на накипном слоевище Lecanora (а - диск, б - край); 2 - поперечный разрез через апотеции (а - гимениальный слой, б - слоевищный край, в - парафизы, г - сумки со спорами, а - эпитещш ж - гипотеций, в - водоросли, и - слоевище); 3 - апотеции на длинной ножке; пылевидная масса наверху - мазедии; 4 - гастеротеции, 5,6 - парафизы с булавовидными окрашенными вершинами
Край апотеция, имеющий вид округлого валика, как защитный футляр, одевает плодущий диск с боков и снизу. В зависимости от его строения различают несколько типов апотециев: леканоровые, лецидеевые и биаторовые. Диск леканоровых апотециев окружен слоевищным краем, который всегда содержит внутри водоросли, а строение и окраска его очень похожи на строение и окраску слоевища. На анатомическом срезе в нем можно различить коровой слой, слой водорослей и слой рыхлой сердцевины (рис. 305, 2). Леканоровые апотеции легко распознаются и по внешнему облику: окраска их края всегда отличается от окраски диска. Если диск может быть коричневым, черным, красным, то край чаще всего бывает более светлым, окрашенным в один цвет со слоевищем,- сероватым, беловатым, серовато-зеленоватым, светло-коричневым и т. д. (табл. 47).
Второй тип апотециев - лецидеевые апотеции - также легко можно распознать по внешнему облику. Обычно они черные и очень твердые, их с трудом можно раздавить на предметном стекле. В отличие от леканоровых край этих апотециев имеет ту же окраску, что и диск: он тоже черный и носит название собственного края. Собственный край никогда не содержит водорослей и отличается по строению от слоевища. У лецидеевых апотециев он состоит из темноокрашенных, плотно прилегающих друг к другу гиф, которые образуют с трех сторон вокруг диска валик, называемый эксципулом (рис. 306).

Рис. 306. Лецидеевые апотеции: 1 - внешний вид темноокрашенных апотециев на накипном слоевище (Lecidea); 2 - поперечный разрез через апотеций (а. - парафизы, б - сумки со спорами, в - эпитеций, г - собственный край, образованный эксципулом, д - слоевище)
Биаторовые апотеции по строению похожи на лецидеевые: у них тоже имеется собственный край, образованный эксципулом. Но, в отличие от лецидеевых апотециев, гифы, образующие эксципул, и сами биаторовые апотеции окрашены в светлые тона - они бывают восковидные, желтовато-коричневатые, красновато-коричневые, розовато- или беловато-желтые и никогда не бывают черные. Биаторовые апотеции отличаются очень мягкой консистенцией, их легко можно раздавить на предметном стекле, как теплый, расплавленный воск.
Гастеротеции, как и апотеции, являются плодовыми телами открытого типа. В них также можно различить диск и край. Но они отличаются от апотециев сильно вытянутой, линейной формой. Обычно гастеротеции имеют вид штрихов, простых или разветвленных линий. Диск у этих плодовых тел очень узкий, напоминающий тоненькую полоску, в то время как край, который может быть и собственным и слоевищным, сильно выступает и возвышается над глубоко погруженным диском (рис. 305, 4). Гастеротеции у лишайников встречаются довольно редко. Они известны лишь у представителей двух родов накипных лишайников - графис (Graphis) и опеграфа (Opegrapha).
Перитеции - плодовые тела закрытого типа, имеющие вид замкнутого кувшина округлой или овальной формы с маленьким отверстием в верхней части (рис. 307). Через это отверстие, которое носит название устьица, происходит выбрасывание зрелых спор. Обычно перитеции почти незаметны на поверхности лишайника: при внимательном рассмотрении можно обнаружить лишь черные точки - устьица перитециев, в то время как само плодовое тело целиком погружено в слоевище. Реже перитеции выступают своими верхушками (рис. 307, 1) или целиком сидят на слоевище. Стенки этого кувшинообразного плодового тела состоят из нескольких слоев (рис. 307,5). Внутреннюю полость перитеция, где образуются сумки со спорами, окружает эксципул. Это темно- или светлоокрашенная оболочка, состоящая из нескольких слоев гиф, поделенных на удлиненные клетки. Нередко эксципул снаружи покрыт еще одной оболочкой - покрывальцем. Покрывальце, как правило, бывает темноокрашенным или черным, может охватывать эксципул со всех сторон или покрывать его лишь до половины; иногда оно развито лишь у устьица, а нередко и вовсе отсутствует. Внутри перитеция развиваются сумки со спорами, которые в нижней и боковой частях плодового тела образуют вместе с парафизами гимениальный слой (рис. 307, 26). Гимениальный слой развивается на гипотеции - узком зернистом слое, прилегающем к внутренней стенке перитеция. Характерно, что у некоторых лишайников в перитециях защитные нити - парафизы - вовсе не образуются или очень рано расплываются в слизь. Это связано с тем, что сумки со спорами надежно защищены самими стенками перитеция и надобность в парафизах исчезает. Особые защитные нитевидные гифы в перитециях развиваются около устьица (рис. 307, 2г). Они называются перифизами и служат для защиты всего внутреннего ядра перитеция от воздействий внешней среды.
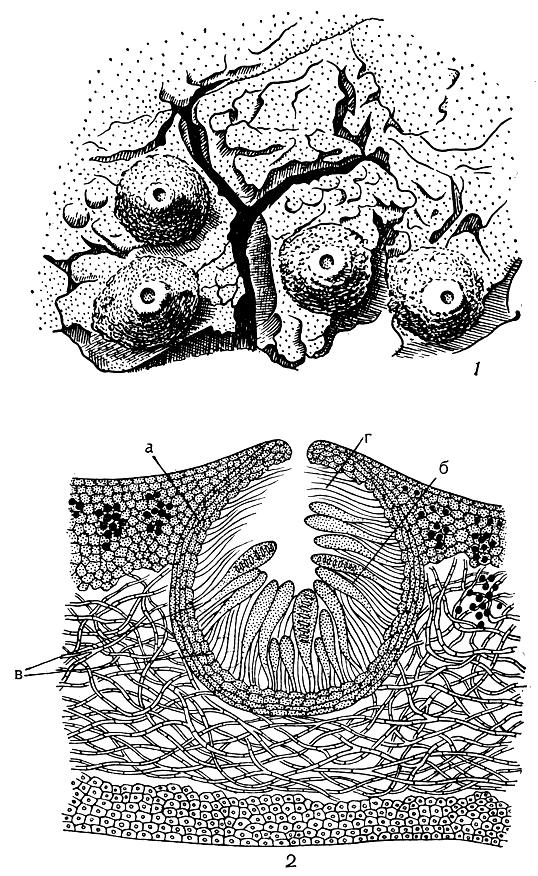
Рис. 307. Перитеции: 1 - внешний вид перитециев; 2 - поперечный разрез через слоевище с погруженным в него перитецием (а - эксципул, б - гимениальный слой, образованный сумками со спорами и парафизами, в - гипотеций, г - перифизы)
Сумки лишайников могут быть самой различной формы: цилиндрической, булавовидной, грушевидной, бутылковидной, бесформенно-мешковидной и др. (рис. 308). Средние их размеры: в длину 40-75 мкм и в ширину 15-25 мкм, но у некоторых родов длина сумок достигает 400 мкм, а ширина - 90 мкм.
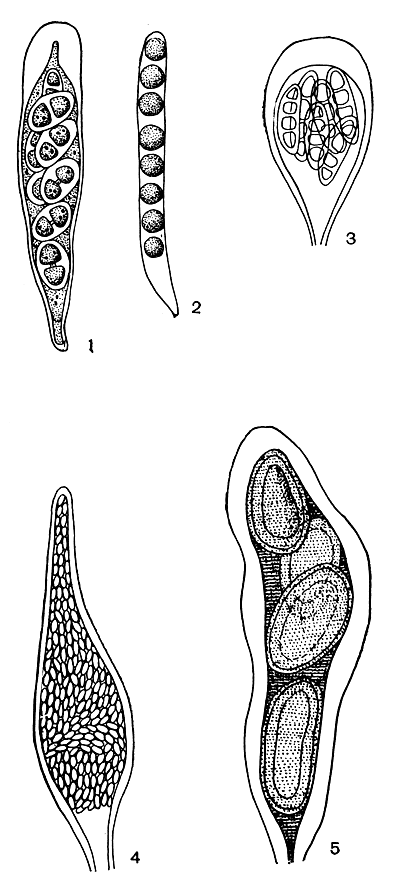
Рис. 308. Форма сумок лишайников: 1 - удлиненно-булавовидная (Buellia); 2 - цилиндрическая (Sphaerophorus); 3 - булавовидная (Arthonia); 4 - бутылковидная (Thelocarpon); 5 - мешковидная (Pertusaria)
У каждого вида лишайника в сумке развивается постоянное количество спор. Чаще всего их бывает 8, но иногда 12-32. У некоторых родов (например, у представителей рода акароспора) в сумке образуется до 200 спор. Однако встречаются лишайники, в сумке которых всегда развивается лишь одна очень крупная (длиной до 300 мкм) спора. Иногда таких спор бывает 2 или 4.
Сумки большинства лишайников имеют одну оболочку и называются унитуникатными. На вершине образуется особый апикальный (верхушечный) аппарат, с помощью которого споры высвобождаются из сумки. Этот аппарат может иметь форму кольца, крышечки и др. У некоторых родов лишайников встречаются сумки с двумя оболочками - битуникатные. У них при выбрасывании спор на некотором расстоянии от вершины происходит поперечный разрыв внешней оболочки и споры выплывают, окруженные тонкой внутренней оболочкой, которая разрывается уже позднее. Битуникатные сумки особенно характерны для асколокулярных лишайников (рис. 309).
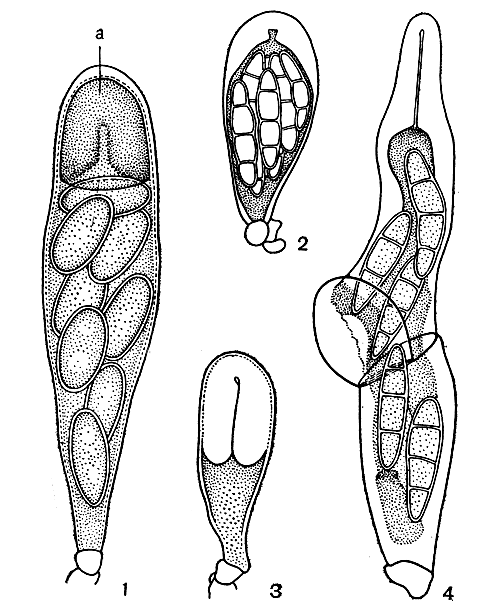
Рис. 309. Сумки лишайников: 1 - унитуникатные сумки с апикальным аппаратом (а) на вершине, с помощью которого споры высвобождаются из сумок; 2,3 - зрелая и молодая битуникатные сумки; 4 - выбрасывание спор из битуникатной сумки
Спора лишайников представляет собой окруженный оболочкой небольшой комочек цитоплазмы с заключенным в ней ядром, ядрышком и иногда несколькими каплями жира. Оболочка споры двухслойная. Внешний слой, толстый и нередко окрашенный, носит название экзоспория, внутренний, более тонкий, называют эндоспорием. Споры чаще всего бывают бесцветными или коричневыми. Обычно их размеры: в длину 1-50 мкм и в ширину 0,5-10 мкм. Самые крупные споры - у накипного лишайника бацидии (Bacidia marginalis), произрастающей на Филиппинских островах и острове Суматра на листьях вечнозеленых растений, они достигают величины 525 мкм.
По форме споры могут быть шарообразными, эллипсоидными, яйцевидными, веретеновидными, игловидными, нитевидными и прямыми, согнутыми, спирально закрученными и др. (рис. 310). Среди них встречаются как одноклеточные, так и двух- и многоклеточные. Помимо поперечных перегородок, в многоклеточных спорах нередко встречаются еще и продольные перегородки. В таких случаях размещение клеток в спорах напоминает кладку кирпичей в стенах и их называют муральными (murus - стена). У лишайников встречается еще один интересный тип спор - биполярные споры, которые специфичны для этих растений и не встречаются у свободноживущих грибов. Биполярные споры двухклеточны и имеют в центральной части толстую перегородку, внутри которой проходит тончайший каналец, соединяющий протопласты обеих клеток (рис. 310, 4).
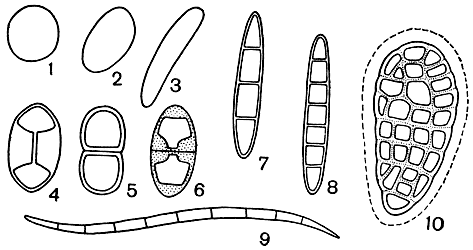
Рис. 310. Споры лишайников: 1 - шарообразные; 2 - эллиптические; 3 - удлиненные; 4 - биполярные; 5 - двухклеточные; 6 - двухклеточные с сильно и неравномерно утолщенными оболочками; 7 - четырехклеточные вытянутые; 8 - восьмиклеточные вытянутые; 9 - игловидные многоклеточные; 10 - муральные
Как уже отмечалось, плодовые тела у лишайников многолетние, образование сумок, их созревание, выход из них спор - сложный процесс, который продолжается не один год. После выхода спор стенки сумки распадаются и вместо пустой сумки снизу, из гипотеция, вырастает новая. Считают, что процесс выхода спор - споруляция приурочена у лишайников к определенной поре года и в различных областях земного шара происходит в разное время. Например, было замечено, что в Западной Европе у многих лишайников споруляция осуществляется особенно активно осенью, зимой и весной, у других происходит круглый год. Но, как показали исследования, у большинства лишайников в условиях эксперимента после увлажнения апотециев споры начинали быстро выбрасываться из сумок. Это привело к мысли, что, возможно, выход спор из сумок связан не столько со временем года, сколько с погодными условиями, прежде всего с влажностью воздуха.
Лишь у немногих лишайников (порядок Caliciales) происходит пассивное освобождение спор в результате постепенного разрушения тонкой и нежной оболочки сумки. Споры некоторое время остаются склеенными по 8 в ряд, а затем рассыпаются, оставаясь, однако, на плодовом теле и образуя пылистую массу, которая позднее разносится ветром. У большинства лишайников споры с силой выбрасываются из сумки. В результате увеличения осмотического давления внутри сумки и с помощью верхушечного аппарата споры могут быть отброшены в сторону на 1-5 см и подброшены вверх на 3 см. При этом их обычно подхватывают воздушные течения и относят порой на довольно значительные расстояния от материнского плодового тела.
Сколько же спор способны продуцировать плодовые тела лишайников? Подсчитано, например, что у лишайника солорина (Solorina saccata) в апотеции диаметром 5 мм образуется 31 тыс. сумок, а в каждой сумке обычно развивается по 4 споры. Следовательно, общее количество спор, продуцируемое одним апотецием, равно 124 000. В течение одного дня из такого апотеция выбрасывается от 1200 до 1700 спор.
Конечно, не все выброшенные из плодового тела споры прорастают. Многие из них, попав в неблагоприятные условия, погибают. Для прорастания споры необходимы прежде всего достаточная влажность и определенная температура. Наблюдения в природе позволили выяснить, что в умеренном климате споры прорастают лучше всего от марта до июня, однако все условия, необходимые для прорастания спор лишайников, пока неизвестны. Например, оказалось, что некоторым нужно, чтобы в непосредственной близости с ними находилась водоросль.
При прорастании одноклеточной споры образуется одна, редко до пяти ростковых трубочек. У муральных спор каждая из их многочисленных клеток может развить одну или несколько ростковых трубочек, но обычно прорастают лишь их конечные клетки. Ростковые трубочки растут, разветвляются и образуют первичный мицелий, который еще не содержит водорослей. Дальнейшее развитие этого мицелия и образование слоевища лишайника происходит лишь в том случае, если его гифы встречают водоросль, соответствующую данному виду лишайника. В поисках этих водорослей гифы мицелия посылают во все стороны длинные тоненькие ищущие гифы. Если на субстрате, где растет первичный мицелий, нужных водорослей нет, этот мицелий обычно скоро погибает. Встретив свойственную данному лишайнику водоросль, гифы первичного мицелия окружают ее со всех сторон, отделяют от субстрата и через некоторое время, воздействуя на нее, вызывают деление водоросли на дочерние клетки, которые тоже, в свою очередь, оплетаются гифами. Так начинается формирование слоевища лишайника.
У некоторых лишайников в результате симбиотической жизни лишайникового гриба с водорослью появилось своеобразное приспособление: споры гриба выбрасываются из плодовых тел вместе с водорослями. Это возможно потому, что в плодовых телах таких лишайников развиваются мелкие гимениальные водоросли. Например, в перитециях чешуйчатого лишайника эндокарпона (Endocarpon) среди сумок, содержащих крупные муральные споры, можно увидеть мелкие водоросли палочковидной, овальной и шарообразной формы. Когда муральные споры этого лишайника выбрасываются из перитеция, вместе с ними бывают выброшены и прилипшие к ним гимениальные водоросли. В дальнейшем эти водоросли увеличиваются в размерах и приобретают нормальную форму, присущую данному лишайниковому фикобионту. В этом случае, следовательно, сразу начинается формирование слоевища лишайника, так как имеются налицо оба компонента сим-биоза. Однако в общей массе лишайников такие случаи единичны, гимениальные водоросли известны лишь у трех родов лишайников: эндокарпона (Endocarpon), стауротеле (Staurothele) и гоногимении (Gonohymenia).
Бесполые спороношения. Помимо спор, образующихся половым путем, у лишайников известны также бесполые спороношения - конидии, пикноконидии и стилоспоры, возникающие экзогенно на поверхности конидиеносцев (рис. 304, 2г). При этом конидии образуются на конидиеносцах, развивающихся непосредственно на поверхности слоевища, а пикноконидии и стилоспоры в особых вместилищах - пикнидиях, строение которых описано выше (рис. 304, 1, 2).
Из бесполых спороношений лишайники чаще всего формируют пикнидии с пикноконидиями. Пикнидии нередко встречаются на слоевищах многих кустистых и листоватых лишайников, реже их можно наблюдать у накипных форм. У таких листоватых лишайников, как пармелия и гипогимния, пикнидии иногда обильно развиваются на верхней поверхности слоевища, у некоторых видов цетрарии (например, у цетрарии исландской) они образуются на верхушках ресничек, окаймляющих лопасти этого лишайника, а у кладоний группы ягелей пикнидии всегда находятся на кончиках их разветвленных подециев.
В каждом из таких пикнидиев образуются в огромном количестве маленькие одноклеточные споры - пикноконидии. Роль этих столь широко распространенных спороношений в жизни лишайника до сих пор не выяснена. Одни ученые, называя эти споры спермациями, а пикнидии - спермагониями, считают их мужскими половыми клетками, хотя до сих пор нет ни экспериментальных, ни цитологических данных, доказывающих, что пикноконидии действительно участвуют в половом процессе лишайников. Сомнение, высказанное по этому поводу, подтверждается тем, что пикнидии встречаются одинаково часто как на слоевищах, формирующих плодовые тела, так и на слоевищах, которые их никогда не образуют или образуют очень редко. Так, например, кладонии группы ягелей почти никогда не развивают плодовых тел, но всегда на кончиках их лопастей присутствуют пикнидии. С другой стороны, нет доказательств, что пикнидии являются спорами бесполого размножения. Пока что удавалось наблюдать прорастание пикноконидий лишь в искусственных условиях, причем эти эксперименты с проращиванием пикноконидий в культуре долго терпели неудачу. Существует предположение, что у одних лишайников пикноконидии выполняют роль половых элементов, в то время как у других превратились в споры бесполого размножения. Действительно, трудно допустить, что эти споры, так часто встречающиеся у лишайников, не участвуют в их размножении.
Иногда в пикнидиях, кроме обычных маленьких одноклеточных пикноконидий, развиваются крупные (длиной до 50-100 мкм) двух- и многоклеточные споры, называемые стилоспорами. Обычно (но не всегда) и сами пикнидии, образующие стилоспоры, отличаются более крупными размерами. Бывают случаи, когда в одной и той же пикнидии развиваются и пикноконидии и стилоспоры. Стилоспоры у лишайников образуются гораздо реже, чем пикноконидии, однако они известны у представителей родов пельтигера, алектория, артония, пармелия и др.
Еще реже у лишайников образуются конидии. До сих пор они были обнаружены на слоевищах лишь двух видов лишайников - у слизистого лишайника лемфолемма (Lempholemma minutulum) и у накипного лишайника калоплака (Caloplaca decipiens). Чаще их образуют изолированные микобионты в условиях культуры. Так, они были обнаружены в чистых культурах микобионтов, выделенных из слоевища некоторых видов родов кладония, буеллия, лецидея и некоторых других. Конидии лишайников очень разнообразны по форме - палочковидные, нитевидные, гантелевидные, грушевидные, эллипсоидные до почти шаровидных. Они могут быть одноклеточными и поперечно-многоклеточными. Как известно, конидии очень часто встречаются у свободноживущих грибов, они широко распространены у многих гифомицетов, а также обычны у аскомицетов. Редкое образование конидий у лишайников, по-видимому, связано с утратой ими своего биологического значения. Если у быстро развивающихся и недолговечных грибов образование конидий - это быстрая форма размножения в течение одного вегетационного периода, то у лишайников с их многолетними слоевищами и плодовыми телами, которые непрерывно, круглый год, в течение ряда лет образуют споры, этот сезонный способ размножения излишен.
Как и в случае с пикноконидиями, до сих пор не ясно, являются ли стилоспоры и конидии спорами бесполого размножения лишайников. Их прорастание также удавалось наблюдать лишь в условиях культур. Вероятно, у гриба лишайников в связи с особенностями жизни в симбиозе с водорослями происходит утрата способа размножения с помощью спор, образованных бесполым путем, столь характерного для свободноживущих грибов. И хотя микобионт образует эти бесполые спороношения, роль их в размножении лишайников, по-видимому, невелика.
Более того, у лишайников, этих симбиотических организмов, в процессе их эволюции наблюдается также постепенная редукция и полового способа размножения, в котором участвует тоже лишь микобионт.
У лишайников спора, как бесполого спороношения, так и отброшенная из плодового тела, должна пройти очень долгий путь, прежде чем из нее разовьется слоевище. Для развития лишайника необходим не только комплекс благоприятных факторов, которые способствовали бы прорастанию споры: даже если спора прорастает и из нее образуется первичный мицелий, это еще вовсе не означает, что из него разовьется слоевище. Развитие лишайника из споры зависит от многочисленных внешних факторов, главный среди которых - встреча с водорослью, свойственной данному лишайнику, по современным представлениям, носит чисто случайный характер. Размножение с помощью спор проис-ходит чрезвычайно медленно, ведь только на формирование плодовых тел и их созревание требуется не менее 4-10 лет. Поэтому в эволюционном развитии лишайников прослеживается тенденция к увеличению в жизни лишайников роли вегетативного размножения, в котором принимают участие сразу и грибной и водорослевый компоненты.
Вегетативное размножение. Если накипные лишайники, как правило, образуют плодовые тела, то среди более высокоорганизованных листоватых и кустистых лишайников имеется немало представителей, которые размножаются исключительно вегетативным путем. Во многих случаях вегетативное размножение этих лишайников осуществляется просто кусочками слоевища, которые отрываютсй ветром или отламываются от чрезвычайно хрупких в сухую погоду слоевищ в результате воздействия животных, человека и других факторов внешней среды. Эти небольшие кусочки слоевища переносятся ветром на новые местообитания и при благоприятных условиях разрастаются в новые слоевища лишайников. Таким образом, например, размножаются очень многие тундровые напочвенные лишайники, представители родов цетрария и кладония, многие из которых почти никогда не образуют плодовых тел.
Вегетативное размножение лишайников осуществляется также соредиями и изидиями - особыми вегетативными образованиями, возникшими в процессе эволюции главным образом у высокоорганизованных форм листоватых и кустистых лишайников. Преимущество размножения лишайников соредиями и изидиями перед половым способом состоит в том, что в нем принимают участие оба компонента лишайника - и гриб и водоросль.
Соредии имеют вид пылинок - микроскопических клубочков, состоящих из одной или нескольких клеточек водоросли, окруженных гифами гриба. Миллионы таких клубочков собираются в мелкозернистую или порошкообразную беловатую, желтоватую или беловато- зеленоватую массу, которая местами покрывает верхнюю поверхность слоевища лишайника или развивается в виде каймы по краям его лопастей (табл. 46, вверху слева). Соредии образуются в слоевище под верхней корой, в зоне водорослей. У разных лишайников процесс образования соредий происходит неодинаково, однако в общих чертах он всегда сводится к следующему. На месте образования соредий каждая клетка водорослей делится на несколько дочерних клеток. В то же время гифы гриба начинают сильно ветвиться, делятся на множество клеток и окружают каждую из дочерних клеток водоросли. В результате этого процесса под коровым слоем слоевища образуется масса водорослей, оплетенная коротковетвящимися многоклеточными гифами. Спустя некоторое время она распадается на отдельные клубочки, каждый из которых представляет собой соредий. Когда клубочков образуется очень много, их масса начинает давить на коровой слой лишайника, тот не выдерживает, разрывается и соредии выступают наружу, на поверхность слоевища (рис. 311, 2). Здесь они образуют порошистую массу, которая легко сдувается и разносится ветром. Попавший в благоприятные условия соредий начинает разрастаться и сразу развивается в новое слоевище лишайника.
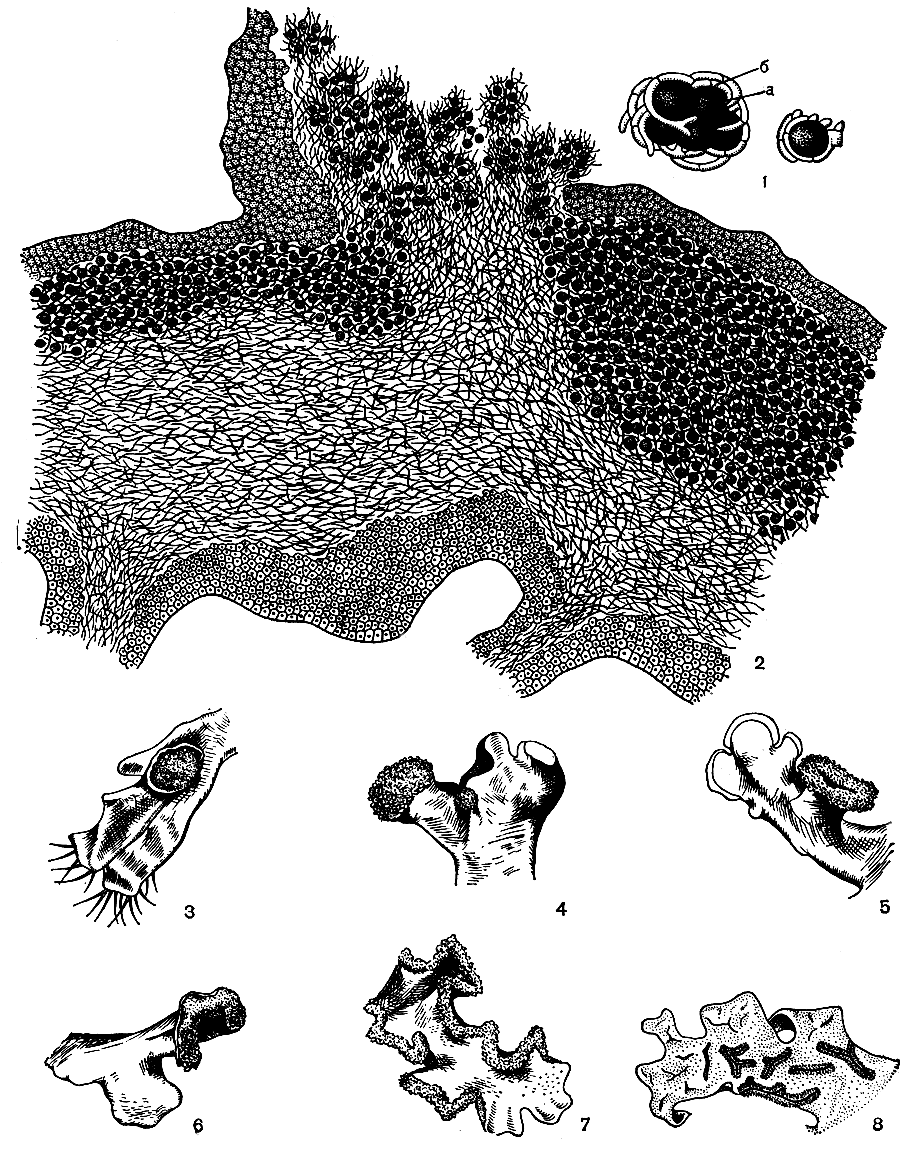
Рис. 311. Вегетативное размножение лишайников с помощью соредий: 1 - соредии (а - клетки водорослей, б - гифы гриба); 2 - схема образования соредий в слоевище; 3-8 - различная форма соралей лишайников: 3 - пятновидная (Physcia orbicularis), 4 - головчатая (Hypogymnia bitteri), 5 - манжетовидная (Меnegazzia pertusa), 6 - губовидная (Physcia tenella), 7 - в виде каймы (Cetraria pinastri), 8 - щелевидная (Parmelia sulcata)

Таблица 46. Листоватые (вверху) и чешуйчатые (внизу) лишайники: вверху слева - Sticta aurata, по краям лопастей кайма ярко-желтых соралей, на нижней поверхности пятнистые ярко-желтые цифеллы; вверху в середине - Parmelia scortea, в центральной части темно-серые изидии; вверху справа - Xanthoria parietina; внизу слева - Psora scalaris; внизу справа - Psora decipiens
Обычно на поверхности слоевища соредии образуют скопления определенной формы, называемые соралями. У каждого вида лишайника, образующего сорали, их форма и расположение на слоевище постоянны. Например, у цетрарии морщинистой (Cetraria pinastri) сорали всегда имеют вид ярко-желтой каймы по краям лопастей (рис. 311,7), у эверниипрунастри (Evernia prunastri) и лобарии легочной (Lobaria pulmonaria) они представляют собой белые округлые пятна (табл. 47, 1). У гипогимнии вздутой (Hypogymnia physodes) сорали образуются на вершине лопастей и имеют губовидную форму, а у менегации продырявленной они бывают манжетовидной формы и т. д. (табл. 52, 2; рис. 311, 5).
Соредии встречаются приблизительно у 30% видов лишайников, главным образом у высокоорганизованных форм. Как правило, виды, образующие соредии, очень редко формируют плодовые тела, и, наоборот, соредии почти не встречаются у лишайников, развивающих апотеции или перитеции. Так, их очень редко можно наблюдать на слоевищах накипных и слизистых лишайников. Соредиеобразование возникло, очевидно, уже в более поздний период существования лишайников.
Изидии - маленькие разнообразной формы выросты верхней поверхности слоевища, одного цвета с ней, а иногда более интенсивной окраски. Они имеют вид бородавочек, зернышек, цилиндрических, булавовидных или коралловидных выростов, иногда маленьких листочков (рис. 312). Если сделать анатомический разрез через изидии, то можно видеть, что внутри их находятся клетки фикобионта лишайника, окруженные грибными гифами (рис. 312, 3). Характерным признаком изидий является то, что они, в отличие от соредий, всегда снаружи покрыты коровым слоем. Если соредии образуются в результате разрыва корового слоя, то изидии - в результате его выпячивания. Форма изидий для каждого вида совершенно определенна и постоянна.
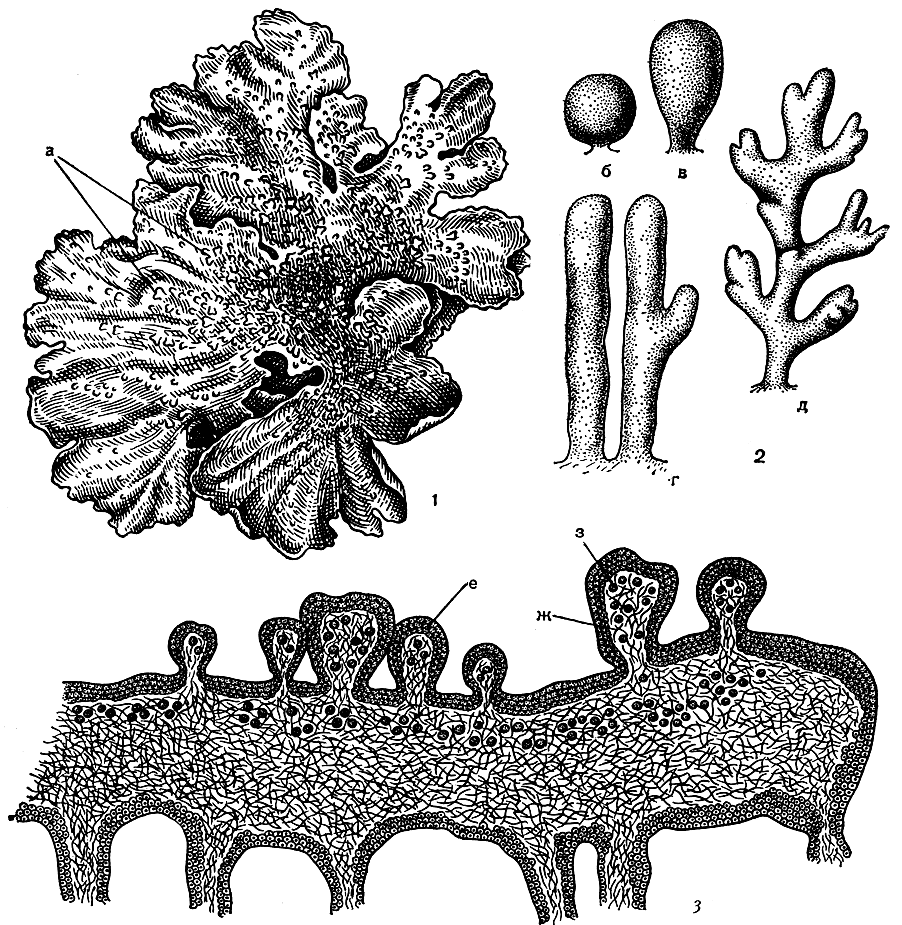
Рис. 312. Вегетативное размножение лишайников с помощью изидий: 1 - слоевище листоватого лишайника Parmelia exasperatula с изидиями (а); 2 - различная форма изидий (б - шаровидная, в - булавовидная, г - цилиндрическая, д - коралловидная); 3 - поперечный разрез через слоевище с изидиями (е - коровой слой, ж - гифы гриба, з - водоросли)
Оторванные от слоевища изидии в благоприятных условиях разрастаются в новые слоевища. Однако значение изидий в размножении лишайников не так велико по сравнению с соредиями. Очевидно, это связано с тем, что обычно изидии не отрываются от слоевища так легко, как соредии, а обламываются лишь животными или человеком. Изидии не только служат для размножения, но и выполняют определенную роль в увеличении ассимиляционной поверхности слоевища лишайника.
Изидии встречаются реже, чем соредии. Они известны приблизительно у 15% видов лишайников. Как и соредии, они характерны главным образом для высокоразвитых форм - кустистых и листоватых лишайников.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'