Порядок десмидиевые (Desmidiales) (Г. М. Мордвинцева)
Десмидиевые водоросли характеризуются удивительным разнообразием очертаний, красотой форм и замечательной симметрией клеток. Водоросли, входящие в состав этого порядка, издавна привлекали к себе внимание не только профессиональных исследователей, но и любителей, благодаря чему сейчас эта группа насчитывает уже около 4000 видов. Уникальные черты организации, своеобразие вегетативного деления и полового воспроизведения, особенности строения оболочки и высокая степень физиологической избирательности среды продолжают и теперь вызывать у исследователей глубокий интерес к десмидиевым.
Общее название "десмидиевые" (греч. "десмос" - связка) было дано этим водорослям в середине прошлого века в связи с тем, что наиболее часто встречаемые и широко распространенные их представители, первыми ставшие известными исследователям, имели клетки, соединенные в длинные нити или цепочки, хотя преобладающее большинство ныне известных форм существуют как одноклеточные организмы. Более того, под этим же названием в течение длительного времени объединяли три различных порядка в классе конъюгат, так как к десмидиевым до недавнего времени относили, кроме порядка собственно десмидиевых, также представителей порядков мезотениевых и гонатозиговых.
Форма клеток. Десмидиевые водоросли в основном одноклеточные организмы. Реже среди них встречаются и колониальные организмы, образующие длинные нити и цепочки или рыхлые колонии. Типичная клетка десмидиевых водорослей (рис. 247) состоит из двух симметричных половинок, так называемых полуклеток, каждая из которых является зеркальным отражением другой. Благодаря своеобразному способу деления клеток у десмидиевых, которое будет описано ниже, одна полуклетка всегда старше другой. Обе полуклетки соединены между собой более узкой частью - перешейком. Угол, возникающий между двумя полуклетками в результате сжатия или сужения клетки, называют синусом.
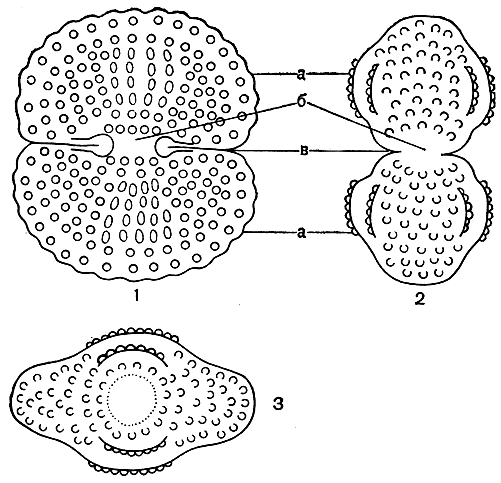
Рис. 247. Внешний вид типичной клетки десмидиевой водоросли в трех разных положениях (схема): 1 - вид спереди; 2 - вид сбоку; 3 - вид сверху, а - полуклетки, б - перешеек, в - синус
Он может быть узким и длинным, закрытым или открытым, с краями, расходящимися наружу от острой или закругленной верхушки.
Внешний вид клеток десмидиевых в большинстве случаев зависит от того, с какой стороны их рассматривают под микроскопом. Различают вид спереди (с широкой, лицевой стороны), вид сбоку и вид сверху (или снизу, рис. 247). Таким образом, клетки имеют три плоскости симметрии. Разнообразие очертаний клетки с различных сторон имеет большое значение в систематике десмидиевых. Несколько видов могут иметь очень сходные очертания при рассматривании их спереди, но при этом резко отличаться друг от друга при рассматривании сбоку или сверху. Однако клетки некоторых видов из родов доцидиум (Docidium) и актинотениум (Actinotaenium) сверху округлые и, следовательно, имеют одинаковые очертания при любых поворотах вокруг длинной оси (рис. 248, 1, 2). Своеобразно строение клетки у видов клостериума (Closterium, рис. 248, 3). Они обычно сужены к концам, не перешнурованы и имеют более или менее серповидную форму, в результате образуются неодинаковые спинной и брюшной края. Спинной край бывает более или менее выпуклым, а брюшной - вогнутым или почти прямым. Однако ясная симметрия полуклеток сохраняется.
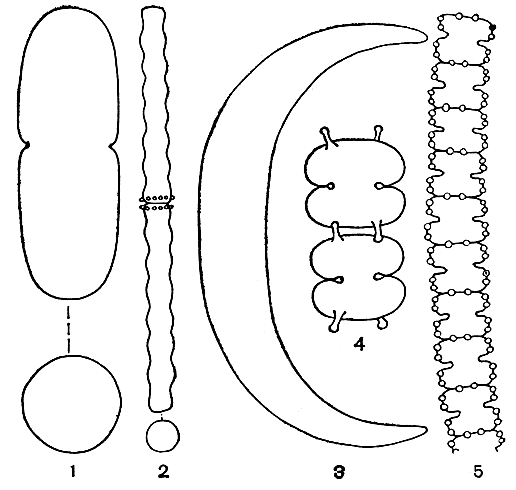
Рис. 248. Разнообразие формы клеток у десмидиевых водорослей: 1 - Actinotaenium cucurbitinum, внизу - вид сверху; 2 - Docidium undulatum, внизу - вид сверху; 3 - Closterium manschuricum; 4 - Sphaerozosma filiformis; 5 - Teilingia granulata
Перешнурованные виды десмидиевых, к которым относится большинство этих водорослей, характеризуются огромным разнообразием формы клеток (рис. 249, 250). У одних клетки сильно вытянутые, цилиндрические, в поперечном сечении округленные или звездчатые; у других - сильно сжатые и плоские. У некоторых представителей развиваются особые отростки и шипы на углах полуклеток. У рыхло-колониальных и нитчатых десмидиевых клетки соединены особыми образованиями (бугорками, гранулами, выступами) или слизью (рис. 248, 4, 5).
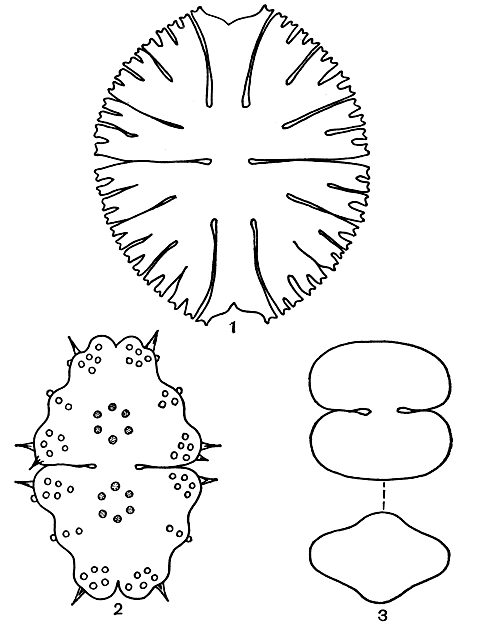
Рис. 249. Разнообразие формы клеток у десмидиевых водорослей: 1 - Micrasterias rotata; 2 - Euastrum divaricatum; 3 - Cosmarium protuberans, внизу - вид сверху
Одним из наиболее характерных признаков десмидиевых является чрезвычайное многообразие очертаний клеток. Края их часто разделены на дольки, или сегменты, или сильно изрезаны. Самые сложные очертания - у клеток из родов микрастериас (Micrasterias), эуаструм (Euastrum) и стаураструм (Staurastrum) (табл. 37, 4, 5; рис. 249, 2, 2; 250). Форма и очертания клеток имеют большое значение при определении отдельных видов.
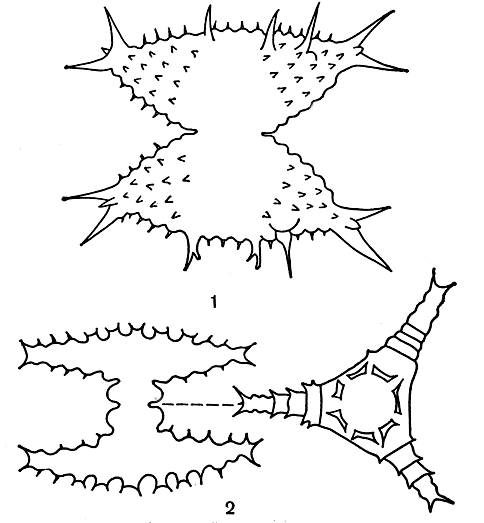
Рис. 250. Разнообразие форм клеток у десмидиевых водорослей: 1 - Staurastrum pelagicum; 2 - Staurastrum cyclacanthum, справа - вид сверху

Таблица 37. Десмидиевые водоросли: 1 - Cosmarium subtumidum ( X 1350); 2 - С. repiforme ( X 900); 3 - С. quinarium ( X 900); 4 - Micrasterias conferta ( X 300); 5 - Euastrum verrucosum
Оболочка клеток. Клетки десмидиевых обладают слоистой оболочкой. Новейшие исследования показали, что она состоит из трех слоев. Самый внешний слой, который до недавних пор не был известен исследователям, кажется под электронным микроскопом то петлеобразным, то гладким или имеет вид пчелиных сот. Химическая природа этого слоя еще неизвестна. Предварительные исследования показывают, что он содержит пектинообразное вещество. У молодых клеток внешний слой оболочки тонкий, у старых достигает значительной толщины. За внешним слоем следует первичная оболочка, затем вторичная оболочка - самый внутренний слой. Первичная и вторичная оболочки при рассматривании клетки в световом микроскопе не отличаются друг от друга и представляются как один целлюлозосодержащий слой. Однако с помощью электронного микроскопа можно хорошо различить их строение. Первичная оболочка состоит из тончайших волоконец - микрофибрилл, имеющих вид пучков соломы, которые перекрещиваются друг с другом. Вторичная оболочка состоит из более широких волоконец - фибриллярных лент, расположенных параллельно микрофибриллам первичной оболочки. Такая оболочка служит хорошей защитой и опорой для содержимого клетки.
Сложность строения оболочки у десмидиевых водорослей усугубляется также наличием в ней особых круглых или овальных отверстий - пор. Они имеют разное значение. Прежде всего поры служат для выделения экскреторной слизи, которая постоянно окружает клетки десмидиевых. Через поры удаляются из клетки ненужные продукты жизнедеятельности. Поры служат и для поступления питательных веществ из окружающей среды в клетку, так как внешний слой оболочки сам по себе непроницаем.
Расположение пор в оболочке у разных видов десмидиевых бывает очень разнообразным. У некоторых представителей поры очень мелкие и многочисленные. Они сосредоточиваются в определенных местах или равномерно распределяются по всей оболочке. У других видов поры бывают крупными и немногочисленными. Они также располагаются в определенных местах, чаще на концах клеток. У третьих видов имеются и мелкие и крупные поры и расположение их также бывает различным.
Поры у десмидиевых представляют собой не просто отверстия в оболочке, а имеют часто сложное строение. Исследования под электронным микроскопом показали, что у этих водорослей имеются два типа строения порового аппарата - тип клостериума и тип космариума.
У представителей рода клостериум поровый аппарат наиболее примитивен. Он состоит из порового канала обратноворонкообразной формы (рис. 251, 1), узкая часть которого, являющаяся собственно порой, расположена во внешнем слое оболочки, а более широкая часть проходит через первичную и вторичную оболочки. Фибриллы этих оболочек пронизывают широкую часть канала, закрывая его своеобразным ситом, через которое может проходить клеточное вещество и выбрасываться наружу через узкую часть. Поровый аппарат типа космариума характеризуется наличием свободного сквозного канала (рис. 251, 2), обычно снабженного различными приспособлениями.
Самое сложное строение порового аппарата встречается, например, в роде микрастериас (рис. 251, 3). Здесь настоящий поровый канал снабжен особыми поровыми нитями. Снизу он начинается на внутренней границе вторичной оболочки поровой луковичкой, проходит через первичную оболочку и внешний слой и заканчивается над внешним слоем конечными запонками. Поровые нити - трубоподобные образования, которые расширяются наружу в бокаловидные конечные запонки. Поровые луковички имеют вид коротеньких гвоздиков с широкой шляпкой, которые погружены своим острием в поровый канал: они сидят в поровом канале, как затвор умывальника в отверстии для стока. Это приспособление в поровом аппарате имеет большое значение. С его помощью поры по мере надобности могут то открываться, то закрываться. В данном случае поровый механизм служит не только для выделения отбросов и поступления питательных веществ, но и предохраняет клетку от нежелательной потери клеточного вещества, а также проникновения в клетку вредных веществ снаружи.
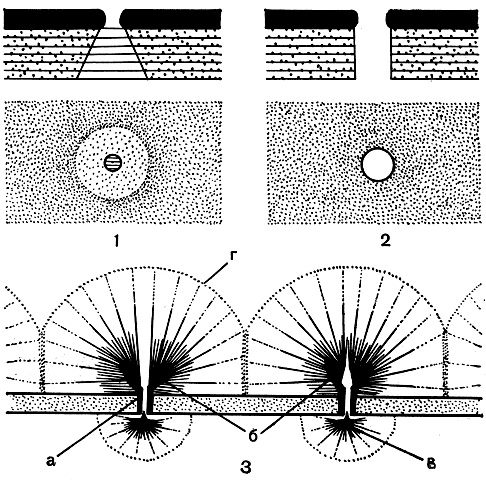
Рис. 251. Строение порового аппарата клеток у десмидиевых водорослей: 1, 2 - общие схемы основных типов порового аппарата в разрезе (в верху ) ис поверхности (внизу ) (1 - тип клостериума, 2 - тип космариума); 3 - схематическое изображение электронно-микроскопической картины порового аппарата у Micrasterias rotata (поперечный разрез): а - поровые нити, б - конечные запонки, в - поровая луковица, г - слизистые призмы
Кроме пор, структура внешней оболочки десмидиевых может быть усложнена наличием особых скульптурных украшений: гранул, ямок, бородавок и шипов, большинство которых расположены в определенном порядке. Они отдельно или в сочетании с особыми выпуклостями, буграми и отростками оболочки создают удивительные узоры, которые придают особую красоту и разнообразие десмидиевым. Эти скульптурные украшения являются важными систематическими признаками при различении таксономических единиц порядка.
Внешний слой оболочки у многих десмидиевых бывает пропитан соединениями железа, отложения которых располагаются у разных видов по-разному, в зависимости от их принадлежности к тому или другому роду. Железо часто откладывается в местах скульптурных украшений и особенно часто в районе перешейка (рис. 252, 1-3). У отдельных видов железа откладывается так много, что оболочка их клеток приобретает желтый, ржавый или темно-коричневый цвет. Способность клеток десмидиевых к накоплению железа считается хорошим систематическим признаком, но все же у некоторых родов оно до сих пор не обнаружено.
Внешний слой оболочки благодаря присутствию пектина и наличию пор часто образует толстый слизистый покров, полностью окутувающий особь или - у нитчатых форм - всю нить (рис. 252, 4, 5). С помощью этого слизистого покрова многие десмидиевые приклеиваются к высшим водным растениям или другим водорослям; другие же увеличивают свою плавучесть для парения в воде.
Рис. 252. Отложение железа на клетках (1-3) и слизистые покровы (4,5) у десмидиевых водорослей: 1 - Cosmarium quadrum; 2 - Xanthidium aculeatum; 3 - Staurastrum gladiosum; 4 - Staurastrum sonthalianum; 5 - Hyalotheca mucosa
У некоторых представителей клетки объединены в нити посредством слизистых тяжей, проходящих между прилегающими верхушками смежных клеток. При этом связь клеток оказывается настолько крепкой, что нити разрываются намного легче в районе перешейка клетки, чем в точках верхушечного прикрепления. Такое склеивание посредством слизи, выделяющейся между прилегающими верхушками клеток, иногда дополняется развитием верхушечных отростков оболочки клеток, как у видов родов сферозосма (Sphaerozosma) и тейлингия (Teilingia) (рис. 248, 4, 5).
У колониального рода космокладиум (Cosmocladium) клетки удерживаются вместе с помощью слизистых стебельков, образуя небольшую, разветвленную, свободноплавающую колонию (рис. 260, 1, 2). У рода оокардиум (Оосаrdium) колония имеет вид полушария, достигающего в диаметре 1-2 мм и прикрепляющегося к известковым скалам. Она состоит из более или менее параллельных, расходящихся лучами слизистых нитей значительной толщины. Каждая нить расширяется по направлению к поверхности колонии и иногда разветвляется. На свободном конце каждой слизистой нити расположена одна клетка. Она размещена так, что ее продольная ось лежит перпендикулярно к оси слизистой нити (рис. 260, 3).
У некоторых видов образуется очень плотная слизь, которая окружает клетку в виде футляра и служит защитой при неблагоприятных условиях: она предохраняет клетку от недостатка влаги и защищает ее от высыхания.
Движение клеток. Десмидиевые водоросли обладают способностью двигаться с помощью особой слизи, которая выделяется через специальные крупные поры, размещенные на концах клеток. Движение свойственно всем представителям десмидиевых, но особенно хорошо оно изучено у видов рода клостериум. Движение у них осуществляется таким образом. Один конец клетки клостериума временно прикрепляется к субстрату, а другой конец выполняет ритмические колебательные движения под углом в 180°. Через некоторое время свободный конец клетки прикрепляется к субстрату, а колебательные движения начинает первый конец. Так, прикрепляясь то одним, то другим концом, клетка клостериума передвигается по субстрату. При температуре воды +33 °С смена позиции клетки происходит через 6-8 мин. У других родов также отмечено неправильное, порой спазматическое движение, но иногда оно имеет плавный характер. Некоторые виды образуют слизистые "ножки", приподнимающие клетку над субстратом.
Все десмидиевые положительно фототаксичны. В ясные солнечные дни они всегда собираются у поверхности воды. Если склянку с живыми десмидиевыми водорослями поставить на окно, то через некоторое время клетки сконцентрируются на освещенной стороне.
Протопласт. У десмидиевых протопласт заполняет всю полость клетки. В цитоплазме находятся хлоропласты, ядро, вакуоли, кристаллики гипса и слизистые тельца.
Симметрия очертаний клеток и украшений оболочки у десмидиевых отражается на симметрии протопласта. Симметрично расположены и хлоропласты, что является отличительным признаком десмидиевых водорослей.
Хлоропласты. У десмидиевых водорослей хлоропласты бывают двух типов: осевые (или центральные) и постенные. У большинства видов встречается осевой хлоропласт, причем почти всегда по одному в каждой полуклетке. Однако у нескольких очень мелких форм, принадлежащих к роду космариум, и у некоторых других в центре каждой клетки имеется только один простой хлоропласт с одним пиреноидом. Хлоропласты имеют обычно густо-зеленый цвет и бывают очень разнообразной формы. Они могут иметь вид звездообразных тел (пениум), осевых конусов с радиальными пластинками (клостериум), стенкоположных лент (плеуротениум), осевых тел с радиальными лучами (космариум) и т. д. (рис. 253).
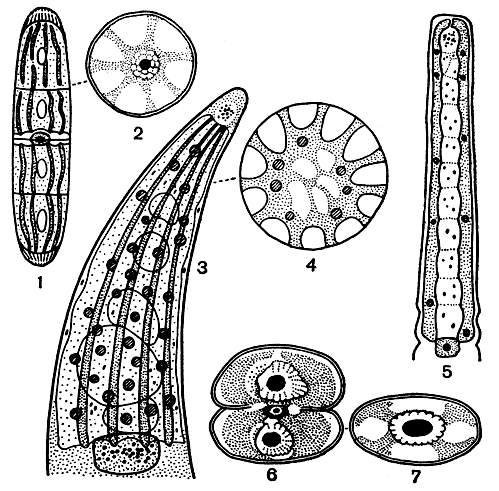
Рис. 253. Хлоропласты десмидиевых водорослей: 1, 2 - Penium spirostriolatum; 3, 4 - Closterium ehrenbergii; 5 - Pleurotaenium trabecula; 6, 7 - Cosmarium depressum (2, 4, 7 - вид на поперечном разрезе)
Постенные хлоропласты встречаются реже, чем осевые, и преимущественно у более крупных форм. У некоторых представителей, особенно из родов эуаструм и микрастериас, имеющих осевые хлоропласты, наблюдается тенденция к развитию и постенных хлоропластов. Центральная часть хлоропласта становится у них более тонкой и исчезает совсем, тогда как периферическая часть развивается очень хорошо. Иногда цельные пластинки хлоропластов в периферической части клеток разделяются на дольки или многочисленные короткие отростки, направленные вперед к оболочке клеток. Благодаря этому площадь хлоропласта значительно увеличивается, что улучшает восприятие света клеткой. Величина и характер расчленения хлоропласта могут быть различными у разных особей одного и того же вида в зависимости от условий их существования. При хорошем питании и освещении клеток хлоропласт хорошо развит и клетки окрашены в темно-зеленый цвет. В неблагоприятных условиях (при недостаточном количестве питательных веществ, при отсутствии или, наоборот, большом избытке какого-нибудь одного вещества) хлоропласты развиваются слабо, преимущественно у стенок клетки. В таком случае клетки имеют бледно-зеленый или желтоватый цвет.
На внешний вид хлоропласта сильно влияет также количество крахмала, содержащегося в нем в виде свободных крахмальных зерен. Их иногда хорошо видно в живых клетках, но еще легче можно обнаружить, если окрасить клетку раствором йода, который вызывает посинение крахмала. Крахмальные зерна образуются и располагаются в основном вокруг пиреноидов. Они могут равномерно покрывать поверхность пиреноида или же располагаться на некотором расстоянии друг от друга. Иногда зерна крахмала возникают в строме хлоропласта. Если крахмала очень мало, то хлоропласт имеет тонкую и плотную структуру и кажется гладким. Иногда крахмала в хлоропласте бывает очень много, и в этих случаях хлоропласт становится массивнее, приобретает более грубую структуру и гранулированный внешний вид. При некоторых условиях крахмальные зерна выбрасываются из пиреноидов в больших количествах и распространяются по хлоропласту. Тогда хлоропласт в результате разбухания крахмальных зерен увеличивается в несколько раз, его структура нарушается и он становится бесформенной массой. Это явление иногда вызывает замечательное разнообразие формы хлоропласта, особенно у видов клостериума, у которых относительная величина центральных осевых и радиальных ребер хлоропласта в большой степени зависит от количества присутствующего крахмала. Количество и положение хлоропластов имеют большое значение для определения некоторых видов и родов десмидиевых.
Количество и положение пиреноидов сильно варьируют у разных представителей десмидиевых и зависят от формы хлоропласта и условий существования клетки. У представителей большинства видов, имеющих крупные постенные хлоропласты, пиреноиды разбросаны без определенного порядка, порой малозаметны. Если виды имеют осевой хлоропласт, пиреноиды обычно находятся в тех частях клетки, в которых хлоропласт наиболее массивен. Количество пиреноидов, внедренных в ось осевого хлоропласта, зависит от его величины. У некоторых видов микрастериаса их может быть более ста, а у мелких видов обычно имеется только один пиреноид. В зависимости от массы накопленных питательных веществ количество пиреноидов изменяется.
Ядро. Десмидиевые водоросли - одноядерные организмы. Только иногда при нарушениях процессов деления клетки образуются урод-ливые формы, которые обладают двумя и большим количеством ядер.
Ядро у большинства видов десмидиевых занимает центральное положение, располагаясь посередине между хлоропластами двух полуклеток у неперешнурованных видов или в перешейке у перешнурованных видов. Величина ядра может колебаться от нескольких до 50 мкм в поперечнике. Ядро может иметь шаровидно-эллипсоидную, квадратную или прямоугольную форму. Ядра содержат одно или несколько ядрышек шаровидной формы. У разных видов число ядрышек и их величина зависят от величины клетки.
Вакуоли. Для протопласта десмидиевых характерны вакуоли. Обычно они содержат кристаллики гипса или слизистые тельца пектиновой природы. У тех форм, которые имеют постенные хлоропласты, эти вакуоли изредка размещены в цитоплазме, но у видов клостериума они могут оказаться в углублениях между продольными радиальными ребрами хлоропласта. Кроме того, здесь имеются также хорошо заметные вакуоли на концах клеток, содержащие, как правило, один или несколько кристалликов гипса. У видов космариума вакуоли размещены между хлоропластами обеих полуклеток в области перешейка.
При нормальных условиях число и расположение вакуолей у одного и того же вида бывают очень постоянными. В большинстве случаев в каждой полуклетке десмидиевых имеются одна крупная (основная) вакуоля и несколько более мелких вакуолей. Величина одних вакуолей может увеличиваться за счет слияния их с другими. При некоторых ненормальных условиях жизни протоплазма клеток десмидиевых может образовывать многочисленные вакуоли, иногда очень значительных размеров.
Вакуольная система десмидиевых послужила предметом многих исследований, но ее полное значение и возможная роль еще до сих пор не совсем ясны. Особенно разнообразные мнения были высказаны по поводу природы и назначения кристаллов гипса в вакуолях на концах клеток. Эти кристаллы находятся в постоянном броуновском движении. Число их в вакуолях зависит от возраста клеток и может изменяться под влиянием различных факторов окружающей среды. Иногда их бывает 1, 2, но не больше 10; в других случаях насчитывали более 40 кристаллов в каждой вакуоле. Во время деления клетки кристаллы гипса могут полностью исчезать.
В вакуолях многих десмидиевых встречаются еще особые слизистые тельца, имеющие вид маленьких подвижных зерен. Эти тельца состоят из органического вещества, так как при нагревании до +64 °С, а также в кислотах и щелочах они растворяются. По-видимому, они представляют собой запасные питательные вещества, связанные каким-то образом с выделением слизи.
Размножение десмидиевых осуществляется вегетативным и половым путем.
Вегетативное размножение. Деление десмидиевых можно хорошо проследить на примере рода космариум. Сначала ядро, лежащее в районе перешейка, делится, образуя два ядра, которые расходятся к центрам обеих полуклеток. После того как ядра передвинулись в полуклетки, в середине перешейка образуется перегородка, тесно примыкающая своими краями к внутреннему слою оболочки (рис. 254, 1). Затем область перешейка вытягивается, в результате чего две старые полуклетки отделяются одна от другой (рис. 254, 2). Верхняя и нижняя части перешейка, лежащие по обеим сторонам поперечной перегородки, начинают раздуваться в виде пузырька, увеличиваться в размерах, после чего наступает расщепление самой перегородки (рис. 254, 3). Образовавшиеся из раздувшихся частей перешейка молодые полуклетки начинают быстро расти и достигают нормального облика, оставаясь соединенными друг с другом своими вершинами (рис. 254, 4, 5). Когда клетки достигают зрелого возраста, их оболочка, являющая собой временное образование, отбрасывается и отпадает, наподобие линяющей кожи. Тогда клетки окончательно разъединяются и начинают самостоятельное существование.
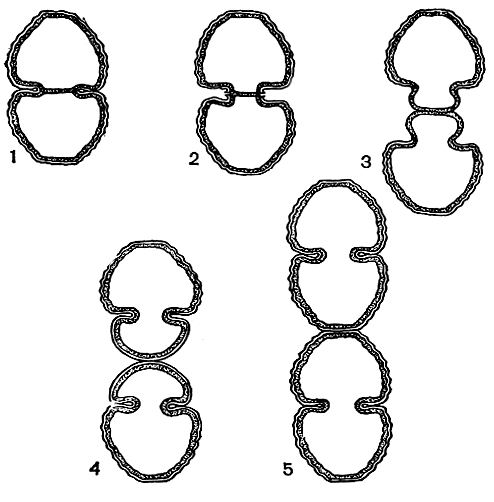
Рис. 254. Схема вегетативного деления клетки десмидиевых водорослей: 1-5 - последовательные стадии деления
Таким образом, как уже упоминалось, в каждой клетке оказываются более старая и более молодая полуклетки. На самых ранних стадиях развития содержимое молодых полуклеток бывает совершенно прозрачным, поэтому клеточные ядра, лежащие в области перешейка, бывают хорошо видны даже без специального окрашивания. Потом протоплазма становится густозернистой, заполняется вакуолями. Наконец, хлоропласты старых полуклеток удлиняются, делятся, и один из них переходит в молодую полуклетку. Иногда клетки после окончательного деления и завершения роста не распадаются, а образуют ложные нити, которые, однако, легко разрываются. В некоторых случаях ложные нити образуются путем сцепления верхушечных отростков и лопастей.
У неперешнурованных форм, как, например, у представителей родов клостериум или пениум, клеточное деление происходит еще более сложным образом.
Половое размножение. Как и у других представителей класса, конъюгация десмидиевых совершается безжгутиковыми апланогаметами, которые выполняют функцию половых клеток. Гаметы двух клеток сливаются, образуя зиготу. Зиготы известны для многих представителей десмидиевых, но подробности полового процесса были зарегистрированы только у нескольких видов. Это связано с тем, что половой процесс у этих водорослей довольно редко встречается в природе, а культивирование их до сих пор было малоуспешным.
У одного из видов космариума половой процесс начинается так: пары клеток с различным половым знаком располагаются рядом перпендикулярно одна к другой и выделяют слизь, которая их полностью окутывает (рис. 255, 1). Некоторое время спаренные клетки лежат рядом, но к началу слияния гамет отодвигаются друг от друга. Первым симптомом наступающего слияния гамет является выработка копулирующими клетками цитоплазматических бугорков. Бугорки увеличиваются, растут навстречу друг другу и наконец соприкасаются, образуя копуляционный канал (рис. 255, 2). Весь процесс длится в течение 1 ч. Как только бугорки соприкоснулись, протопласты обеих клеток начинают отделяться от клеточных стенок и двигаться к общему центру в копуляционный канал (рис. 255, 3, 4). К этому времени в каждой клетке в районе перешейка образуется отверстие, которое расширяется при выходе гаметы. Отделение протопласта от стенок обычно продолжается 13-25 мин. Через 4-7 мин, после того как протопласты соприкоснулись, слияние заканчивается и между пустыми оболочками образуется зигота не правильной формы (рис. 255, 5, 6). Иногда гаметы не отделяются полностью от оболочек клеток, и в таких случаях образовавшаяся зигота бывает прочно прикреплена к пустым оболочкам клеток.
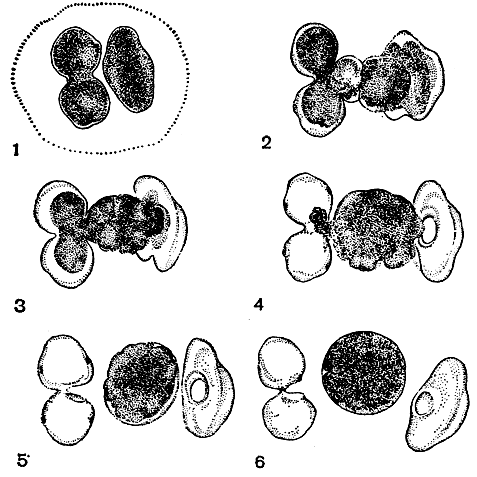
Рис. 255. Различные стадии конъюгации у Cosmarium botrytis: 1 - ранняя стадия (одна клетка лежит боком, другая - в поперечной плоскости, вид сверху); 2 - выработка копуляционного канала и начало копуляции; 3, 4 - слияние содержимого обеих клеток; 5, 6 - формирование зиготы
В первые минуты образования зиготы в ней появляются многочисленные сокращающиеся вакуоли. Маленькие вакуоли сливаются в более крупные. Каждая вакуоля пульсирует в течение 4-7 мин, затем сжимается, и ее содержимое выходит наружу. Действие пульсирующих вакуолей длится от 4 до 32 мин, до тех пор пока прозрачные места зиготы перестают быть видимыми. За этот период (от 1 до 3,5 ч) зигота значительно сокращается. Внутри ее хлоропласты и пиреноиды располагаются в периферическом слое. Затем начинают появляться первые признаки орнаментации зиготы - конусовидные бугорки, разбросанные по ее поверхности. Постепенно они вытягиваются и приобретают вид шипов. За время разрастания шипов, которое продолжается 2-3 ч, зигота опять увеличивается в размерах и становится такой же, как при слиянии гамет.
До окончания роста шипов зигота сохраняет цитоплазматическую оболочку. Когда шипы полностью сформируются, происходит быстрое изменение в их преломляющем индексе, что говорит о том, что в это время образуется новый слой оболочки. В зрелой зиготе оболочка состоит из трех слоев.
Экзоспорий - это самый наружный, целлюлозный слой оболочки зиготы. Во многих случаях он бывает орнаментирован различными скульптурными украшениями. Окончательное формирование экзоспория происходит за несколько дней. Вначале он имеет вид тонкой кожицы, затем на нем образуются выросты различной длины и формы, заполненные протоплазмой. В дальнейшем выросты увеличиваются в размерах, уплотняются и превращаются в крепкие зубцы, шипы или разветвленные отростки. Иногда экзоспорий бывает бородавчатым, ячеистым или пунктированным. У представителей некоторых родов, особенно стаураструм и микрастериас, экзоспорий бывает очень сложно и определенно орнаментирован, что во многих случаях является хорошим признаком при определении вида. Средний слой оболочки зиготы - мезоспорий - также состоит из целлюлозы, пропитанной пробковидным веществом. Он бывает окрашен в коричневый, красный или оливковый цвет. Самый внутренний слой оболочки зиготы называют эндоспорием. Он состоит из целлюлозы и обладает способностью сжиматься. Формирование мезоспория требует более длительного времени, чем формирование экзоспория.
У представителей разных родов десмидиевых существует много мелких, но интересных различий в процессе конъюгации и формирования зигот. Во многих случаях между копулирующими клетками возникает копуляционный канал. Это приводит к значительному разнообразию зигот по форме, строению и положению в клетках. Так, кроме зигот, сразу освобождающихся от оболочек образовавших их клеток (рис. 256, 1, 2), у некоторых десмидиевых возникшие зиготы захватывают часть пустых полуклеток, а их углы оказываются снабженными роговидными выростами (рис. 256, 3). У части представителей нитчатых форм перед началом копуляции происходит распад нити на отдельные клетки. При этом у рода гиалотека (Hyalotheca) копуляционный канал сохраняется до окончательного сформирования зиготы. Он значительно расширяется от образовавшейся в нем зиготы, и на нем остаются прикрепленными четыре пустые оболочки полуклеток (рис. 256, 4). У других нитчатых десмидиевых зиготы располагаются или между двумя рядами пустых полуклеток (рис. 256, 5), или внутри одной из копулирующих клеток (рис. 256, 6). При этом наблюдается половая дифференциация. Одна из двух гамет - "мужская клетка" - постепенно переходит в копуляционный канал, а затем в другую - "женскую клетку". После слияния обеих гамет происходит развитие зиготы в "женской клетке". К ней прикреплены копуляционный канал и пустая оболочка "мужской клетки".
У десмидиевых может происходить также образование двойных зигот. Они развиваются при копуляции четырех клеток, возникших путем последовательного деления одной особи (рис. 256, 7). У видов клостериума способ образования двойных зигот иной: в каждой клетке сначала развиваются две особые гаметы, которые, сливаясь, дают двойные зиготы. Очень редко в образовании зигот десмидиевых принимают участие три или четыре особи.
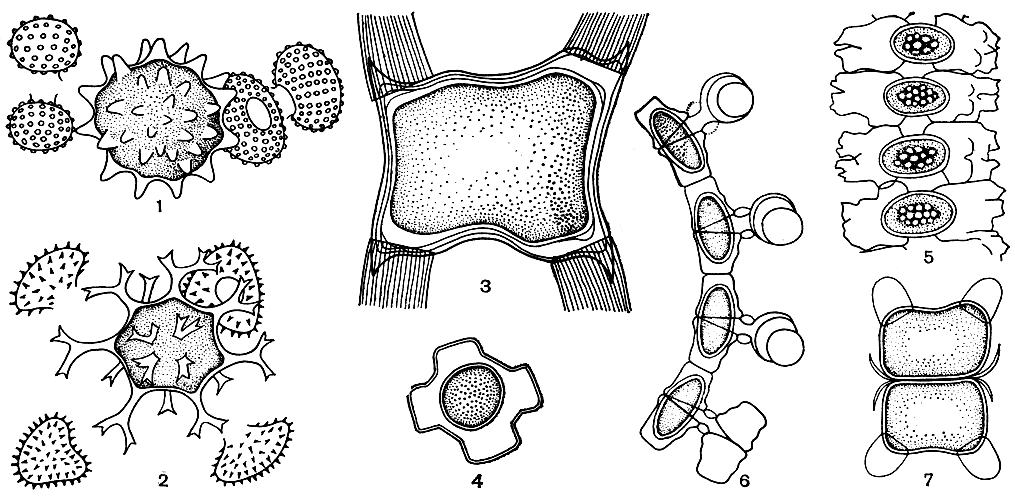
Рис. 256. Зиготы десмидиевых водорослей: 1 - Cosmarium portianum; 2 - Staurastrum hirsutum; 3 - Closterium kuetzingii; 4 - Hyalotheca dissiliens; 5 - Desmidium swartzii; 6 - Bambusina brebissonii; 7 - Penium didymocarpum
Конъюгация наиболее часто встречается у форм, имеющих наибольшие возможности для встречи физиологически разных клеток. Такая возможность чаще всего возникает у нитчатых и колониальных водорослей. Поэтому, например, представители гиалотеки и десмидиума часто встречаются в состоянии конъюгации. Однако у колониального космокладиума, где близость клеток, казалось бы, достаточно обеспечена общей колониальной слизью, конъюгация встречается редко. Вероятно, колонии часто представляют здесь чистые клоны (клон - потомство одной клетки, образовавшееся путем вегетативного размножения), и тогда конъюгация становится невозможной из-за отсутствия физиологически разных клеток.
После слияния гамет и образования зиготы наступает длительный период покоя, который может продолжаться всю зиму и больше. В зиготе два из четырех или один из двух хлоропластов разрушаются (в зависимости от того, сколько их в клетке). Ядра гамет лежат рядом во время периода покоя, а затем сливаются как раз перед прорастанием зиготы. Процесс прорастания зиготы удавалось наблюдать очень редко, и поэтому он еще слабо изучен.
Прорастание зигот у десмидиевых происходит весьма своеобразно. В отличие от других зеленых водорослей оболочка зиготы у них разрывается в самом начале прорастания, высвобождая в окружающую среду еще не разделенное содержимое. Освободившийся протопласт окружен только тонкой гиалиновой оболочкой, за которой следует тонкий слой, образованный липидными телами. Большую часть протопласта занимают светлая вакуоля и два неслившихся хлоропласта. Далее в течение 1,5-2 ч оба хлоропласта постепенно продвигаются к противоположным сторонам протопласта, а его общий объем уменьшается в связи с постепенной потерей воды из вакуоли. Затем происходит деление диплоидного ядра. В результате двух последовательных делений, из которых первое является редукционным, образуются четыре гаплоидных дочерних ядра. Перед этим происходит разделение протопласта на две части. Обе части содержат по одному из первоначальных хлоропластов и по два сестринских ядра. Одно из ядер в каждом вновь сформированном протопласте отмирает, а другое увеличивается в размерах. В результате образуются два проростка. В дальнейшем посередине каждого проростка образуется перетяжка, типичная для большинства десмидиевых водорослей. Оболочка проростка всегда гладкая; он лишен типичных видовых признаков. При первом же вегетативном делении проросток приобретает типовые черты вида, образовавшего зиготу. Два проростка, образованные одной зиготой, имеют, как правило, различные половые знаки. Это говорит о том, что разделение полов у десмидиевых происходит при первом делении ядра. Однако у некоторых представителей число проростков достигает четырех, а иногда развивается всего один проросток.
У десмидиевых довольно часто встречаются межвидовые половые гибриды, особенно у родов микрастериас и эуаструм, и в этом случае две зрелые полуклетки имеют признаки двух различных видов.
В некоторых случаях при определенных условиях начавшийся процесс копуляции прерывается. Тогда содержимое готовой к копуляции клетки округляется и окружается плотной оболочкой. Образуются так называемые партеноспоры, которые по своей форме и строению очень напоминают зиготы. Однако, в отличие от последних, они бывают значительно меньших размеров и имеют только одно ядро. Изредка встречается еще один вид спор - апланоспоры. Они, как и партеноспоры, образуются из всего содержимого клетки и покрываются особой плотной оболочкой, но возникают без всякого участия полового процесса, под воздействием резких колебаний температуры или при внезапном и почти одновременном старении оболочки обеих полуклеток. У десмидиевых имеются еще так называемые хламидоспоры. Они представляют собой бесполые стадии, которые образуются при неблагоприятных для жизни водоросли условиях. При их образовании содержимое более старой полуклетки передвигается к границе между двумя по лук летками, выделяет новую оболочку и отделяется от оболочки старой полуклетки. Развивающаяся хламидоспора покрывается толстой бесцветной оболочкой, состоящей из двух половинок. В благоприятных условиях хламидоспора делится и прорастает, образуя вполне нормальные особи.
В некоторых случаях при прорастании зиготы, а также при вегетативном делении клеток наблюдаются сильные отклонения формы клеток от нормального типа. В результате получаются различные уродливые (тератологические) формы. Наблюдения тератологических форм показало, что они могут возникать от различных причин. Так, при неполном клеточном делении происходит только деление ядра, а разделительная поперечная перегородка между полуклетками не образуется, в результате чего возникают уродливые клетки, состоящие из трех частей. Крайние части представляют собой нормальные полуклетки, а посередине между ними находится уродливая вздутая часть различной формы. Неполное деление встречается у большинства десмидиевых, особенно часто у родов космариум и клостериум. Особенностью некоторых видов является образование аномальных форм с неодинаковыми очертаниями вполне развившихся полуклеток и совершенно нормальной оболочкой. У рода клостериум, например, часто наблюдаются сигмоидные формы, у которых одна полуклетка повернута к другой на 180°.
Особенно большое разнообразие аномальных форм однажды удалось наблюдать у одного вида эуаструма (Euastrum didelta), собранного в высокогорном болоте в Украинских Карпатах. Здесь встречались клетки с двойными и тройными полярными лопастями, а также с резкими различиями в морфологии материнской и дочерней полуклеток, которые заключались в неодинаковом числе боковых извилин или в полном отсутствии извилин и более упрощенной полярной лопасти. Иногда лопасти совершенно отсутствовали и одна из полуклеток имела округленные очертания, напоминающие полуклетку космариума. Некоторые клетки вдобавок срастались между собой попарно в одном участке полуклетки, образуя что-то вроде "сиамских близнецов". Аномальные формы возникают, по-видимому, при неблагоприятных условиях существования клеток и особенно часто встречаются осенью, однако их наблюдали и в летнее время - в июле и августе. Возможно, прямой причиной являются здесь резкие колебания температуры в течение суток, так как аномальные формы особенно обычны в полярных и высокогорных холодных областях.
Местообитание и распространение. Десмидиевые - типично пресноводные организмы, морских видов среди этих водорослей нет. Правда, некоторые из них были найдены в слабосолоноватых водоемах морских побережий или в более соленых континентальных водоемах, однако существует мнение, что десмидиевые попадают туда случайно и через некоторое время погибают.
Десмидиевые предпочитают открытую местность, их редко можно увидеть в тенистых лесах и глубоких канавах. Встречаются они в небольших водоемах и часто обильно населяют пруды, спокойные горные озера со скалистым дном и скалистыми берегами. В быстро текущей воде их, как правило, не обнаруживают. Живут они также в речках среди обрастаний, на влажных скалах, в гипновых и сфагновых болотах. Сфагновые болота - постоянные места обитания десмидиевых, однако здесь численность их зависит от того, к какому типу относится болото. Огромное разнообразие форм десмидиевых встречается в сфагновых низинных болотах (эвтрофных и мезотрофных). Здесь они образуют или слизистые скопления, свободно плавающие в понижениях между кочками, или бурые и зеленые пленки на листьях и стеблях мхов или на отмерших стеблях высших растений. Иногда их можно обнаружить лишь на ощупь - по скользким, влажным стеблям и листьям сфагнума. Лучший способ сбора десмидиевых в таких случаях - это выжимание воды из мхов через планктонную сетку или без нее. Олиготрофные сфагновые болота характеризуются бедным качественным и количественным составом десмидиевых водорослей.
Растут десмидиевые главным образом в мягких водах с низким рН и при наличии гуминовых кислот (в сфагновых низинных болотах), но некоторые виды клостериума и стаураструма обитают в нейтральных и даже щелочных водах. Большинство видов десмидиевых предпочитают чистые воды, но ряд представителей родов клостериум, космариум и стаураструм встречаются и в сильно загрязненных местах, в сточных водах, развиваясь в некоторых случаях в массовом количестве. Некоторые из десмидиевых ведут аэрофитный образ жизни, поселяясь на влажных скалах, на почве среди мхов и нитчаток, в подушках сфагновых мхов на болотах. Имеются сведения о нахождении десмидиевых в почвах.
Большинство десмидиевых - бентосные организмы, однако некоторые виды родов стаураструм, ксантидиум и микрастериас хорошо приспособлены к планктонному существованию благодаря хорошей плавучести за счет развития радиальных полых отростков и шипов и слизистых покровов. Десмидиевые встречаются на всех глубинах, где есть подходящий химический состав воды и достаточное освещение.
Распространены десмидиевые не только на равнинах, но и в горах. Их можно встретить на высоте до 3100 м над уровнем моря. В северных районах СССР десмидиевые распространены на равнинных местах, богатых торфяными болотами и озерами. В степной полосе количество десмидиевых сильно уменьшается, а в пустынях их очень мало.
Десмидиевые весьма чувствительны к окружающим условиям. Главным фактором, определяющим их распространение и количество, является геологическая природа местности. Области распространения гранитов, как правило, отличаются большим разнообразием форм этих водорослей; области распространения известняков, напротив, очень бедны ими, хотя отдельные виды десмидиевых селятся в водоемах с большим количеством извести (например, некоторые виды рода оокардиум). Особенно богаты десмидиевыми те области, где подпочва образована палеозойскими и докембрийскими отложениями. Состав почвы отражается на значении рН среды; наибольшее количество видов встречается при рН ниже 7.
Осадки также играют немаловажную роль в распространении этих водорослей. Обильная флора десмидиевых развивается там, где выпадает большое количество дождей или грунтовые воды подходят близко к поверхности почвы.
Десмидиевые встречаются по всему миру. Существует мнение, что примерно две трети видов этих водорослей являются космополитами или широко распространенными видами. И все же многие виды десмидиевых населяют лишь определенные географические районы и могут быть оценены как эндемики. В тропиках встречаются ряд видов и даже родов, которые до сих пор не обнаружены в других частях земного шара.
Собирать десмидиевые лучше всего в ясную солнечную погоду. В небольших неглубоких водоемах их собирают с поверхности воды при помощи небольшого сачка из плотной шелковой ткани (лучше всего густое мельничное сито, употребляемое для планктонных сеток). Слизь, которая остается на сите, затем осторожно смывают в склянку. Можно просто зачерпывать склянкой или баночкой воду в понижениях между кочками или подушками сфагнума. Иногда десмидиевые развиваются на погруженных в воду растениях в таком количестве, что буроватую слизь, в которую они внедрены, можно собрать просто руками. Если слизь очень нежная, то ее можно извлечь из воды, осторожно опустив в нее руку ладонью кверху так, чтобы листья или стебли растения могли проходить между пальцами и как можно ближе к ладони. Сомкнув пальцы и держа ладонь согнутой, поднимают осторожно руку к поверхности, тогда слизь со стеблей скапливается на ладони, откуда ее нужно немедленно перелить в склянку. Можно также выжимать эти растения над широким горлышком склянки или соскабливать с них слизь с помощью ложки или перочинного ножа. Для сбора планктонного материала из больших озер нужны конусообразные планктонные сетки из мельничного газа, через которые любым способом фильтруют большое количество воды. Со дна водоемов десмидиевых лучше всего собирать специальным стаканчиком или склянкой. Такой материал обычно загрязнен песком и илом, поэтому его полезно помещать в плоскую посуду и оставлять при хорошем освещении. Через некоторое время десмидиевые появятся на поверхности в виде слизистой массы, которую легко удалить с помощью небольшой пипетки.
Систематическое положение десмидиевых среди других водорослей сейчас прочно установлено, хотя по этому поводу существовали различные, иногда противоречивые мнения. Сначала десмидиевых относили к простейшим животным. Позднее, когда они были причислены к растениям, возникли разные мнения по поводу их места в общей системе водорослей. Некоторые исследователи объединяли десмидиевых вместе с диатомовыми, основываясь на сходстве их полового процесса и строения оболочки. Однако это объединение было построено на очень поверхностном суждении, так как структура оболочек и их скульптура, поровый аппарат, хлоропласты и пигменты совершенно различны в обеих группах. Иногда десмидиевых вместе с зигнемовыми считали изолированной группой среди зеленых водорослей и отделяли от них совершенно. Сейчас большинство альгологов относят десмидиевых к конъюгатам, которые являются признанной составной частью зеленых водорослей, так как пигменты и продукты метаболизма у них в основном те же самые.
Десмидиевые входят в состав класса конъюгат как один из порядков. Положение их внутри класса определяется родственными связями с другими его порядками.
Существуют две точки зрения на происхождение десмидиевых. Согласно одной из них, одноклеточные формы произошли от нитчатых предков в результате их распада на отдельные клетки. Если принять это предположение, то, во-первых, следует признать близкие родственные связи десмидиевых с зигнемовыми, а во-вторых, в этом случае приходится считать их более примитивной и вырождающейся группой среди конъюгат, имеющей первоначальную ретрогрессию от нитчатых форм. Однако против сближения десмидиевых с зигнемовыми говорят существенные различия в организации их клеток. Нельзя признать и примитивность десмидиевых, так как структура их клеточной оболочки и строение порового аппарата свидетельствуют об их сложности и высокой специализации.
Другая точка зрения заключается в том, что нитчатые формы произошли от одноклеточных форм, имеющих простую организацию клетки. Это мнение разделяют большинство альгологов. Согласно такому взгляду, наиболее примитивными среди конъюгат являются мезотениевые водоросли. От них берут начало две линии развития. Одна линия привела к гонатозиговым и далее к зигнемовым; другая - к высокоразвитым десмидиевым. Самыми примитивными среди десмидиевых являются те представители, которые характеризуются цилиндрическими, радиально-симметричными клетками с усеченными или округленными верхушками и отсутствием срединной перетяжки. Такие клетки встречаются у представителей рода пениум, отчасти клостериум и плеуротениум. Некоторые виды рода пениум очень близки по своему строению к представителям мезотениевых. Дальнейшая эволюция десмидиевых шла в направлении усложнения формы клеток и структуры их оболочки, а также усложнения и увеличения разнообразия их хлоропластов.
До настоящего времени систематика десмидиевых основывается главным образом на морфологических и в некоторой мере на цитологических признаках. Для разграничения родов десмидиевых широко используют форму клетки, ее симметрию и орнаментацию. Отдельные роды десмидиевых легко отличаются один от другого, но все же границы многих родов очень неопределенны и расплывчаты. Ни в одной другой группе водорослей нет такого большого количества переходных форм, как у десмидиевых. В связи с этим систематика десмидиевых очень запутана и несовершенна. До сих пор в пределах этого класса существуют искусственные полиморфные роды, как, например, космариум (около 2000 видов) и стаураструм (около 1500 видов).
Основываясь на форме и структуре клеток, структуре клеточной оболочки и строении порового аппарата с учетом данных электронно-микроскопического изучения, мы разделяем порядок десмидиевых на три семейства: пениевых (Peniaceae), клостериевых (Closteriaceae) и космариевых (Cosmariaceae).
Семейство пениевые (Peniaceae)
Семейство пениевых содержит только один род пениум (Penium). Клетки у этих водорослей одиночные, прямые, цилиндрические или эллипсоидные, с округленной или срезанной верхушкой, слабо перетянутые или без перетяжки, сверху круглые. Оболочка бывает гладкая, пунктированная, продольно или спирально по штрихованная, иногда гранулированная, бесцветная или окрашенная, без пор. Хлоропласты осевые, с радиальными пластинками, по одному-два в каждой полуклетке. Пиреноиды одиночные, иногда по два в клетке. У некоторых видов имеются терминальные вакуоли с движущимися кристаллами гипса. Зиготы шаровидные, эллипсоидные, четырехугольные или неправильных очертаний, образуются между двумя копулирующими клетками. Для СССР известно 10 видов. Наиболее широко распространены, например, Penium polymorphum, P. spirostriolatum (рис. 257, 2, 2).
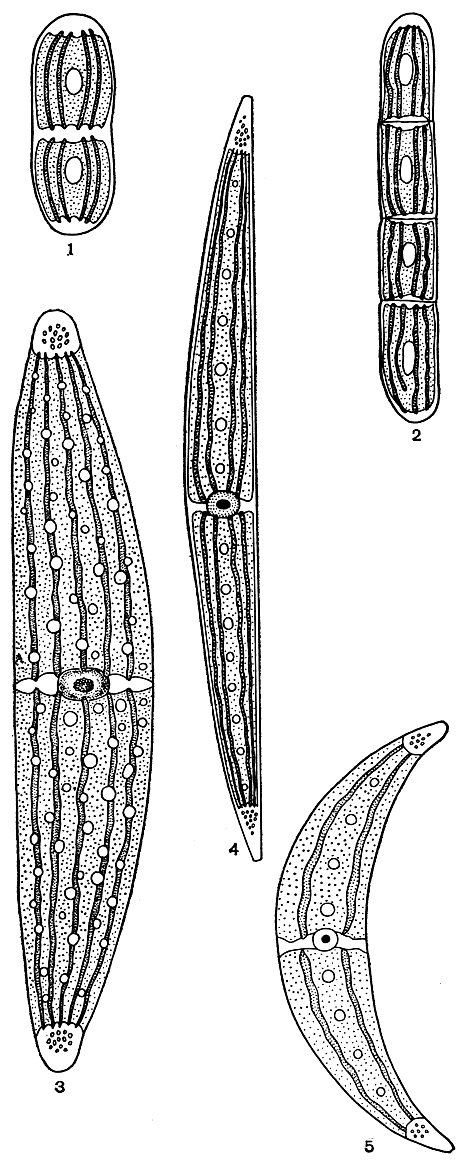
Рис. 257. Виды пениума и клостериума: 1 - Penium polymorphum; 2 - P. spirostriolatum; 3 - Closterium lunula; 4 - G. acerosum; 5 - G. leibleinii
Семейство клостериевые (Closteriaceae)
Эта группа также содержит только один род клостериум (Closterium). Клетки у его представителей одиночные, редко склеивающиеся в пучки, обычно серповидно-согнутые, реже прямые, сильно вытянутые и заметно суживающиеся на концах, без срединной перетяжки, на поперечном сечении круглые. Деление всегда происходит в средней части клетки. Ряд видов образует пояски. Оболочка гладкая или поштрихованная. Поры всегда присутствуют; они бывают разнообразной формы - круглые, овальные или продолговатые. Поровый аппарат состоит из порового канала обратноворонкообразной формы, с ситовидным основанием, расположенным в фибриллярном слое оболочки, и порового отверстия во внешнем слое оболочки. Хлоропласты осевые, с разным количеством продольных ребер, по одному, реже по два в каждой полуклетке. Пиреноиды шаровидные, эллипсоидные, по 1-2-20 в клетке; они расположены в один осевой ряд или разбросаны
беспорядочно по всему хлоропласту. Терминальные вакуоли постоянно присутствуют, по одной, реже по две-три на каждом конце клетки, с одним-двумя или несколькими движущимися кристалликами гипса. Зиготы различной величины и формы - шаровидные, эллипсоидные, почти треугольные или четырехугольные, с гладкой или окрашенной оболочкой. В СССР встречается 77 видов. Среди самых распространенных могут быть названы Closterium асеrosum, Cl. leibleinii, Cl. lunula (рис. 257, 3-5).
Семейство космариевые (Cosmariaceae)
Эта группа включает много родов, имеющих клетки очень разнообразной формы и величины, глубоко или, реже, слабо перетянутые, одиночные или соединенные в колонии или нити. Сверху они также имеют очень разнообразную форму и могут быть эллиптическими, ромбовидными, круглыми, многоугольными, двух- или многорадиальными. Деление клеток происходит в районе перешейка. Оболочка всегда пронизана порами. Поровый аппарат нередко имеет сложное строение и состоит из порового канала, поровых нитей, поровой луковицы и поровых запонок.
Из рода доцидиум (Docidium) в СССР известно несколько видов. Клетки у представителей этого рода вытянутые, прямые, почти цилинд-рические, слабо перетянутые посередине, сверху круглые, с волнистым краем. Полуклетки с ровными или волнистыми боковыми сторонами, у основания несколько вздутые и снабженные венчиком из продольных складочек оболочки, имеющих вид бугорков. Верхушки полуклеток срезанные, без скульптурных украшений. Оболочка гладкая, пунктированная или слегка поштрихованная. Хлоропласты осевые, с несколькими неправильными продольными ребрами. Пиреноиды располагаются вдоль оси; число их варьирует, иногда их бывает более 15. Терминальные вакуоли отсутствуют. Зиготы неизвестны.
Наиболее распространен Docidium baculum, обычен и D. undulatum (рис. 248, 2).
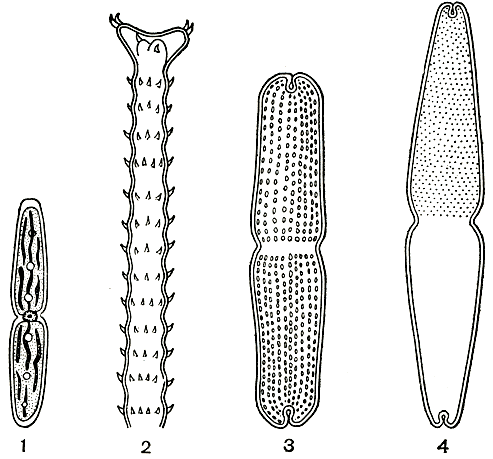
Рис. 258. Десмидиевые: 1 - Pleurotaenium minutum; 2 - Triploceras gracile, часть клетки; 3 - Tetmemorus brebissonii; 4 - T. granulatus
У водорослей из рода плеуротениум (Pleurotaenium) клетки прямые, удлиненно-цилиндрические, слабо перетянутые посередине, в районе перетяжки, у основания полуклетки, они имеют кольцеобразный шов, сверху бывают круглые или звездчато-лопастные. Полуклетки с хорошо заметным базальным вздутием, которое, в отличие от предыдущего рода, никогда не бывает складчатым, и ровными, прямыми или волнистыми боками. Верхушки полуклеток срезанные или притупленно-округленные, гладкие или снабженные венчиком из гранул или шипов. Оболочка клеток гладкая, пунктированная, ямчатая, гранулированная. В каждой полуклетке по одному хлоропласту. Они лентовидно-постенные, с разбросанными пиреноидами. Хорошо заметные терминальные вакуоли содержат кристаллики гипса. Зиготы шаровидные, широкоэллипсоидные, с гладкой или по-крытой коническими шипами оболочкой.
В СССР известно около 10 видов. Одним из наиболее распространенных является Pleurotaenium minutum (рис. 258, 1).
У представителей рода триплоцерас (Triploceras) клетки вытянутые, умеренно перетянутые, прямые, изредка слегка согнутые, с волнистыми или гладкими боковыми сторонами, на концах переходящие в различной длины отростки, которые заканчиваются заостренными шипиками. Оболочка гладкая или несет многочисленные кольца шипов или выростов. Хлоропласты осевые, по одному в каждой полуклетке, с несколькими продольными ребрами и пиреноидами, расположенными вдоль оси. Зиготы неизвестны.
В нашей стране известно всего два вида, из них более распространен Triploceras gracile (рис. 258, 2).
У рода тетмеморус (Tetmemorus) клетки бывают прямые, удлиненные, почти цилиндрические, веретеновидные или эллипсоидные, слабо перетянутые, сверху круглые или широкоэллипсоидные. Полуклетки постепенно суживаются от основания к верхушке, которая обычно округленная и всегда с глубоким узким срединным вырезом. Оболочка пунктированная, поштрихованная или гранулированная, на верхушках утолщенная. Хлоропласты осевые, состоящие из узкой центральной части и боковых продольных ребер или пластинок, по одному в каждой полуклетке. Пиреноиды располагаются вдоль оси или по одному в центре хло-ропласта. Зиготы шаровидные или четырехугольные, развиваются между копулирующими клетками.
В нашей стране встречаются три вида, из них обычен Tetmemorus brebissonii (рис. 258, 3, 4).
У водорослей из рода актипотениум (Actinotaenium) клетки прямые, цилиндрические, веретеновидные или гитаровидные, слабо перетянутые, сверху всегда круглые. Полуклетки с более или менее широкоокругленными, выпуклыми или срезанными верхушками. Оболочка гладкая или ямчатая, иногда утолщенная на верхушке с внутренней стороны или нежно гранулированная на верхушках полуклеток, пронизанная неправильно рассеянными или расположенными в определенном порядке крупными или мелкими порами. Хлоропласты звездообразные, пластинчато-звездообразные или ленточнопостенные, с одним-двумя центральными пиреноидами, реже со многими пиреноидами в каждой ленте постенного хлоропласта. Зиготы шаровидные, эллипсоидные, угловатые, квадратные, реже двойные, с гладкими или бородавчатыми стенками.
В СССР известно около 30 видов. Одним из наиболее распространенных является Actinotaenium cucurbitinum (рис. 248, 1).
У рода эуаструм (Euastrum) клетки относительно короткие, обычно заметно сжатые, разнообразной формы и величины, глубоко перетянутые, с узко линейным закрытым синусом, слегка раскрывающимся в наружной части, эллиптические или узкоэллиптические. Полуклетки трехлопастные, имеющие одну полярную лопасть с более или менее узким срединным вырезом на верхушке и две боковые лопасти, которые бывают цельными или делятся на лопасти второго порядка. В центре каждой полуклетки, а иногда и в углах лопастей имеются симметрично расположенные вздутия различной величины, часто орнаментированные гранулами или бородавками. Сверху клетки узкоэллиптические, со вздутиями на боковых сторонах или полюсах. Оболочка гладкая, пунктированная, гранулированная или мелкобородавчатая.
Хлоропласты осевые, по одному, редко по два в каждой полуклетке, с одним или многочисленными пиреноидами. Зиготы шаровидные, редко эллипсоидные, покрыты крепкими, конусовидными, иногда изогнутыми отростками, простыми или разветвленными, тупыми или заостренными на концах.
В СССР известно 57 видов. Примерами могут служить Euastrum verrucosum (табл. 37, 5) и Е. divaricatum (рис. 249, 2).
К роду микрастериас (Micrasterias) относятся наиболее высоко дифференцированные и красивые виды десмидиевых. Клетки у них часто большие, плоские, в некоторых случаях почти дисковидные, круглые или широкоэллиптические, глубоко перетянутые. Синус узколинейный по всей длине или более или менее сильно открывающийся наружу. Полуклетки разделены глубокими вырезами на три лопасти: полярную и две боковые. Полярная лопасть бывает очень разнообразной формы и величины; обычно она возвышается над боковыми, отделяясь от них широко открытыми вырезами, и имеет узко- или широковыемчатую верхушку, часто снабженную шипами. Боковые лопасти бывают цельные или разделенные глубокими вырезами на лопасти второго, третьего, четвертого, пятого и шестого порядков; конечные лопасти обычно заканчиваются шипиками. Сверху клетки вытянуто-эллиптические, узкоромбовидные, часто со слабыми вздутиями на боковых сторонах.
Оболочка пунктированная, гранулированная или шиповатая. Хлоропласты осевые, по одному в полуклетке, с изрезанными краями и многочисленными пиреноидами. Зиготы шаровидные, эллипсоидные, покрытые крепкими шипами.
В нашей стране встречается 21 вид. Хорошими примерами могут служить Micrasterias conferva (табл. 37, 4), М. rotata (рис. 249, 1).
У представителей рода космариум (Cosmarium) клетки относительно короткие, округленных очертаний или правильно угловатые, удли-ненные или укороченные, более или менее глубоко перетянутые. Синус различной глубины и формы. Полуклетки не разделены на лопасти и не имеют верхушечного выреза, очень разнообразных очертаний: сверху эллиптические, ромбические или круглые, часто с одним-двумя срединными вздутиями. Оболочка гладкая или пунктированная, ямчатая, гранулированная, бородавчатая или покрытая сосочками. Хлоропласты обычно осевые, по одному, редко по два в каждой полуклетке, с одним пиреноидом; реже хлоропласты бывают постенные, состоящие из узких лентовидных полос, в каждой из которых находится по одному пиреноиду. Зиготы шаровидные, эллипсоидные, кубические, покрытые ямками, сосочками, простыми или разветвленными шипами.
В СССР встречается около 500 видов, многие из них широко распространены, например Cosmarium subtumidum, С. reniforme, С. quinarium (табл. 37, 1-3), С. caelatum, С. protuberans (рис. 249, 3).
У рода ксантидиум (Xanthidium) клетки удлиненные, всегда сжатые, глубоко перетянутые. Полуклетки разнообразных очертаний: эллиптические, эллиптически-шестиугольные, трапециевидные или многоугольные, всегда с плоской верхушкай; в центре полуклетки имеется утолщение, часто ямчатое, окрашенное в желтый или коричневый цвет, или выступающий бугор, покрытый гранулами, шипами или зубцами. Сверху клетки эллиптические, с буграми посередине каждого бока. Оболочка гладкая или пунктированная, снабженная на углах полу клеток парными или одиночными шипами, иногда шипы имеются по всей оболочке или в центре полуклеток. Хлоропласты постенные, по четыре в полуклетке, с одним или несколькими пиреноидами; иногда они бывают звездообразные, с одним пиреноидом. Зиготы шаровидные, обычно с простыми или вильчатыми шипами.
В СССР известно более 20 видов. Наиболее широко распространен Xanthidium antilopaeum (рис. 259, 1).
У представителей рода стауродесмус (Staurodesmus) клетки относительно короткие, четырехугольных очертаний, большей частью глубоко перетянутые. Синус выемчатый или треугольный, с острой или тупой верхушкой, открытый. Перешеек часто удлиненный. Полуклетки широкоэллиптические, круглые, веретенообразные, эллипсоидные, но чаще всего имеют вид срезанной пирамиды; верхние или боковые углы их снабжены одиночными длинными или короткими шипами, которые могут быть параллельными, расходящимися или сходящимися, иногда шипы в виде маленьких сосочков на верхних углах. Сверху клетки эллиптические или 3-5-угольные. Оболочка у них гладкая, почти лишенная орнаментации (кроме простых радиальных шипов), пронизанная очень мелкими порами или выступающими порами-органеллами, иногда оболочка бывает ямчатая.
Хлоропласты осевые, по одному в полуклетке, каждый с одним пиреноидом. Зиготы шаровидные, гладкие или снабжены шипами.
В нашей стране известно около 50 видов. Наиболее распространенными являются Staurodesmus incus, S. convergens (рис. 259, 2, 3) и некоторые другие.
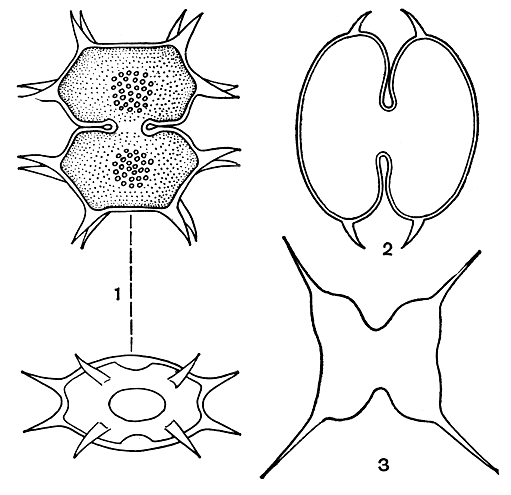
Рис. 259. Десмидиевые: 1 - Xanthidium antilopaeum; 2- Staurodesmus incus; 3 - S. convergens
У водорослей из рода стаураструм (Staurastrum) клетки очень разнообразной формы, со сложной структурой, обычно глубоко перетянутые. Синус чаще широко открытый, с острой верхушкой. Полуклетки полукруглые, эллиптические, продолговатые, чашевидные, треугольные, углы округленные, острые или вытянутые в более или менее длинные отростки или шипы. Сверху клетки трех-, многоугольные, редко веретенообразные. Оболочка гладкая или шиповатая. Хлоропласты осевые, по одному в каждой полуклетке, с одним центральным пиреноидом, иногда постенные. Зиготы шаровидные или угловатые, снабженные простыми или вильчатыми шипами.
В СССР известно более 200 видов. Многие широко распространены, например: Staurastrum pelagicum, S. cyclacanthum (рис. 250, 7, 2).
У представителей рода космокладиум (Cosmocladium) клетки соединены в колонии посредством простых или двойных слизистых не-правильно ветвящихся нитей. Обычно они свободно плавают, но иногда прикрепляются к другим водорослям. Клетки космариевидные, мелкие, обычно сжатые, симметричные в трех направлениях, глубоко перетянутые. Полуклетки почти пирамидальные, эллиптические или почти почковидные, с широким перешейком и узким синусом, сверху эллиптические. Оболочка у них гладкая. Хлоропласты осевые, обычно содержатся по одному в полуклетке. Зиготы более или менее шарообразные, но иногда угловатые, снабженные короткими толстыми и тупыми шипами.
У нас известно 5 видов. Наиболее часто встречаются Cosmocladium saxonicum и С. pusillum (рис. 260, 7, 2).
У рода оокардиум (Oocardium) клетки собраны в шарообразные колонии диаметром 1-2 мм. Колонии состоят из радиальных слизистых тяжей, инкрустированных известью. Каждая нить расширяется к поверхности колонии и иногда разветвляется. На свободном конце каждой нити расположена одна клетка перпен-дикулярно к оси нити. Клетки космариевидные, мелкие, сжатые, значительно укороченные, слабо перетянутые. Оболочка гладкая. Хлоропласты осевые, по одному в полуклетке, каждый хлоропласт содержит один пиреноид. Зиготы угловатые, с несколькими сосочковидными горбиками.
Известен всего один вид - Oocardium stratum (рис. 260, 3).
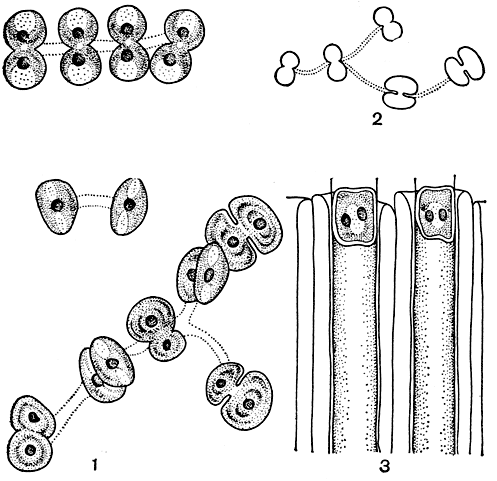
Рис. 260. Десмидиевые: 1 - Cosmocladium saxonicum; 2 -C. pusillum; 3 - Oocardium stratum
У водорослей из рода сферозосма (Sphaerozosma) клетки собраны в однорядные, иногда скрученные слизистые нити. Обычно они очень мелкие, эллиптические или четырехугольные, часто укороченные, более или менее сжатые, всегда перетянутые посередине, с хорошо обозначенным перешейком, с боковыми шипами или без них. Синус различной глубины, закрытый или открытый. На верхушках полуклеток имеется по два небольших, веретенообразных, головчатых отростка, расположенных диагонально и плотно крестообразно прилегающих к соответствующим отросткам соседней клетки. Сверху клетки эллиптические. Оболочка у них гладкая, иногда крупнопористая. Хлоропласты осевые, по одному в полуклетке, с одним центральным пиреноидом.
Зиготы круглые, четырехугольные или продолговатые, гладкие или украшенные простыми шипами.
В СССР известно 5 видов, например Sphaerozosma filiformis (рис. 248, 4).
Род тейлингия (Teilingia) имеет клетки, собранные в однорядные, ровные или слегка скрученные слизистые нити. Часто клетки удлиненные или одинаковые в длину и ширину, более или менее сжатые, эллиптические или четырехугольные. Синус довольно глубокий, выемчатый, открытый, с округленной верхушкой. Верхушка каждой полуклетки снабжена четырьмя гранулами. Сверху клетки эллиптические. Оболочка гладкая или покрыта немногочисленными гранулами. Хлоропласты осевые, с одним центральным пиреноидом. Зиготы шаровидные, эллиптические, кубические, гладкие или шиповатые.
У нас известно 4 вида, в том числе Teilingia granulata (рис. 248, 5).
У водорослей из рода спондилозиум (Spondilosium) клетки соединены верхушками в длинные нити посредством слизистых подушечек. Обычно они небольших размеров, сжатые, всегда более или менее глубоко перетянутые, с узким, закрытым или открытым синусом разнообразной формы. Полуклетки эллиптические, пирамидальные или четырехугольные, часто с широко срезанной или вогнутой верхушкой, сверху эллиптические, треугольные или трехлопастные. Оболочка гладкая или гранулированная. Хлоропласты осевые, по одному в полуклетке, с одним центральным пиреноидом или несколькими пиреноидами, расположенными в один ряд. Зиготы обычно шаровидные, гладкие или шиповатые.
В СССР известно более 10 видов, чаще других встречается Spondilosium pulchellum (рис. 261, 1).
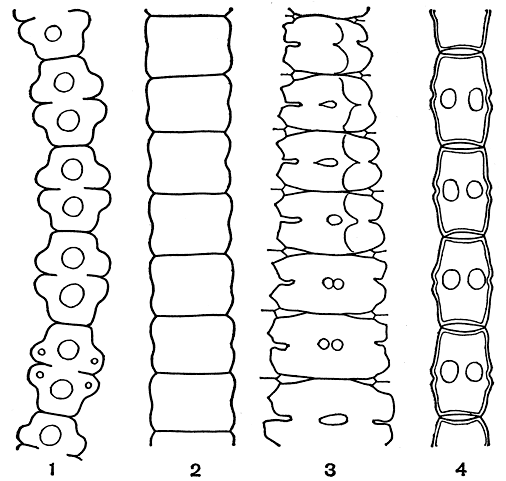
Рис. 261. Десмидиевые: 1 - Spondilosium pulchellum; 2 - Hyalotheca dissiliens; 3 Desmidium swartzii; 4 - Bambuzina brebissonii
У рода гиалотека (Hyalotheca) клетки соединены в однорядные, иногда скрученные нити, заключенные в толстую слизистую обвертку, исчерченную короткими поперечными бороздками. Клетки цилиндрические, очень слабо перетянутые или без перетяжки. По лук летки трапециевидные, почти квадратные или продолговатые, с прямыми или слабовыпуклыми боками. Сверху клетки круглые или почти круглые. Оболочка гладкая или гранулированная. Хлоропласты осевые, звездообразные, с одним центральным пиреноидом. Зиготы шаровидные, гладкие, иногда заключенные в особые образования неправильных очертаний, состоящие из гаметангия, соединенного с широкой конъюгационной трубкой.
В СССР известно более 6 видов. Наиболее широко распространена Hyalotheca dissiliens (рис. 261, 2).
У видов рода десмидиум (Desmidium) клетки соединены в удлиненно-скрученные нити посредством гребневидных утолщений или усеченных отростков, иногда они бывают заключены в толстую слизистую обвертку. Клетки четырехугольные в очертаниях, обычно укороченные, часто сильно сжатые, умеренно перетянутые, с более или менее хорошо заметным синусом. Сверху клетки эллиптические, треугольные или четырехугольные. Оболочка гладкая; в каждой полуклетке под перешейком имеется один поперечный ряд ямок. Хлоропласты осевые, звездообразные, по одному в полуклетке, с массивными, радиально отходящими от центра лопастями и с одним пиреноидом в центре или в каждой лопасти.
Зиготы шаровидные или эллипсоидные, гладкие или с короткими плоскими коническими сосочками.
В нашей стране известно около 10 видов. Наиболее широко распространен Desmidium swartzii (рис. 261, 3).
У водорослей из рода бамбузина (Bambuzina) клетки, соединенные в слабо скрученные нити, имеют цилиндрическую или бочонкообразную форму, они очень слабо перетянутые, часто вздутые и волнистые в основании каждой полуклетки над синусом, сверху круглые, часто с двумя противоположными сосочками. Хлоропласты осевые, с шестью радиальными пластинками, по одному в каждой полуклетке. Зиготы почти шаровидные, овальные, гладкие.
В СССР известно Звида. Широко распространена Bambuzina brebissonii (рис. 261, 4).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'