Отдел эвгленовые водоросли (Euglenophyta) (Т. А. Сафонова)
Эвгленовые водоросли - обычные обитатели небольших пресных стоячих водоемов. В летние месяцы можно наблюдать, как в небольшом пруду или луже внезапно зеленеет вода. Причиной этого позеленения ("цветения") может быть массовое развитие эвгленовых водорослей. Под микроскопом в капле воды, взятой из такой лужи, видны многочисленные зеленые веретеновидные клетки. Легко изгибаясь, меняя форму тела, они быстро движутся во всех направлениях. Обычно это эвглены (Euglena) - центральный род, по имени которого назван весь отдел.
Эвгленовые водоросли - это один из немногих отделов водорослей, организация представителей которого ограничена в основном монадной структурой. Сюда относятся микроскопические (длиной 4-500 мкм) одноклеточные организмы, снабженные одним или двумя жгутами и активно двигающиеся. Движение - характерное свойство эвгленовых водорослей и лишь немногие представители этого отдела, имеющие пальмеллоидную структуру, утратили его.
Форма тела эвгленовых водорослей хорошо приспособлена к движению в воде. У большинства видов она удлиненная, овальная, эллипсоидная и веретеновидная. Последняя среди эвгленовых водорослей встречается особенно часто, и поэтому иногда ее называют эвгленообразной (рис. 193). Имеются виды и с шаровидными клетками, но они сравнительно немногочисленны.
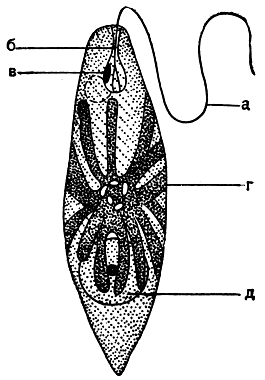
Рис. 193. Эвглена зеленая (Euglena viridis): a - жгут; б - глотка; в - глазок; г - хлоропласт с парамилоновыми зернами в центре; д - ядро с ядрышком
Клетки эвгленовых водорослей на поперечном сечении обычно округлые. В этом случае их можно было бы считать построенными по радиальной симметрии, но последнюю нарушает спиральное расположение штрихов и других элементов орнаментации, а также внутреннее строение клетки. Поэтому более правильно считать клетки эвгленовых водорослей асимметричными. Наиболее ярко асимметрия выражена у высокоорганизованного рода факус (Phacus). Клетки факусов сплющены в виде листообразной пластинки, снабжены килевидными выростами и хвостовым отростком, большей частью изогнутым. Некоторые винтообразно-скрученные клетки факусов при взгляде сверху напоминают гребной винт (рис. 202, 5).
Эвгленовые водоросли не имеют настоящей оболочки, подобной таковой у высших растений. Защитную роль выполняет наружный слой эктоплазмы - перипласт. Те виды, у которых перипласт мягкий, эластичный, обладают способностью менять форму тела. Такие клетки при своем движении то изгибаются, то раздуваются на одном конце и утончаются на другом, как бы переливая свое содержимое из одного конца клетки в другой (рис. 194). Эту способность эвгленовых водорослей называют метаболией.
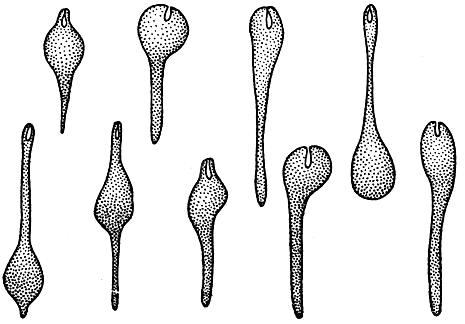
Рис. 194. Изменение клетки Euglena mutabilis с помощью метаболичных движений
Виды с более плотным перипластом сохраняют постоянную форму тела и не способны к метаболии.
Перипласт эвгленовых водорослей редко бывает гладким, обычно он покрыт штрихами или рядами точек, бугорков, проходящих спирально (реже продольно) от переднего конца к заднему. Под перипластом расположены так называемые слизистые тельца, которые выделяют слизь через поры перипласта.
У трахеломопасов (Trachelomonas) и стромбомонасов (Strombomonas) поверх перипласта образуется плотный "домик" - вначале он тонкий и бесцветный, но с возрастом пропитывается солями железа и марганца и от этого окрашивается в различные оттенки бурого, золотистого и коричневого цветов. "Домики" стромбомонасов обычно слегка шероховатые или морщинистые, а у трахеломонасов более богато орнаментированы: покрыты точками, бугорками, мелкими щетинками, крупными шипами различного строения.
Для выхода жгута в передней части "домика" имеется отверстие, часто окруженное выростом стенки "домика" - горлышком (табл. 28, 1, 2; рис. 195, 196).
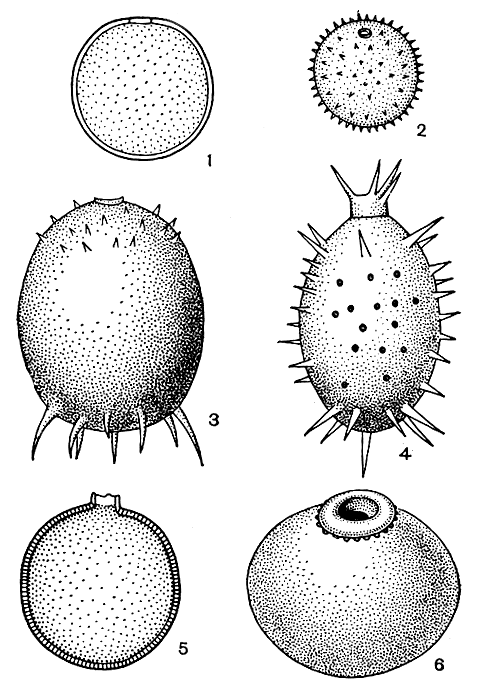
Рис. 195. 'Домики' некоторых видов рода Trachelomonas: 1 - Trachelomonas volvocina; 2 - Tr. globularis; 3 - Tr. armata; 4 - Tr. hystrix; 5 - Tr. planctonica; 6 - Tr. curta
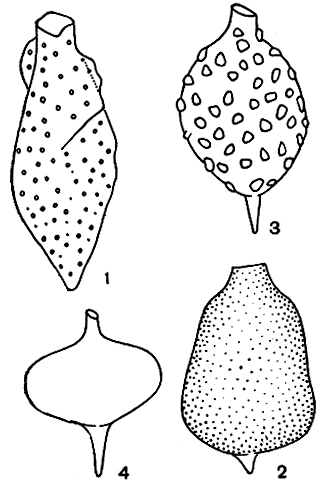
Рис. 196. 'Домики' различных видов рода Strombomonas: 1 - Strombomonas subcurvata; 2 - Str. acuminata; 3 - Str. fluviatilis; 4 - Str. volgensis
На переднем конце клетки эвгленовых водорослей находится углубление, продолжающееся внутрь в виде трубки, - "глотка", которая оканчивается расширением - резервуаром (рис. 197). Глотка - органоид выделительной системы. Сократительные вакуоли, в которых скапливается жидкость с растворенными продуктами обмена веществ, изливают свое содержимое в этот резервуар, откуда через глотку оно выводится наружу.
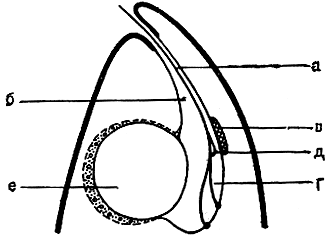
Рис. 197. Схема строения переднего конца клетки эвгленовых водорослей: а - жгут; б - глотка; в - глазок (стигма); г - резервуар; д - фоторецептор; е - вакуоля перед ее опорожнением в резервуар
Процесс излияния происходит ритмично через 20-30 сек.
Движение эвгленовых водорослей совершается с помощью жгута. Жгут прикреплен своим основанием к базальному телу на дне резерву-ара. Через глотку он выходит наружу. У большинства видов жгут один; некоторые эвгленовые снабжены двумя жгутами равной или неравной длины. Но типичным для эвгленовых следует все же считать наличие двух жгутов, так как даже у одножгутиковых форм имеется тонкии и короткий остаток второго жгута, который не выходит из глотки, а соединяется в ее пределах с длинным жгутом (рис. 197).
Движение большинства эвгленовых - это активное плавание. Работая жгутом, клетка одновременно вращается вокруг своей оси и как бы ввинчивается в воду. При этом важную роль играют килевые выросты и конечный отросток - стабилизаторы движения. У некоторых бесцветных представителей эвгленовых, живущих на дне водоемов среди ила, наряду с плаванием (перанема, гетеронема) наблюдается и ползание по дну или другому субстрату (леталомонас, анизонема). Клетка ползает, как бы ощупывая дорогу впереди своим жгутом.
У анизонемы клетка снабжена двумя жгутами. Один из них направлен вперед и выполняет роль двигателя; второй волочится сзади, играя, вероятно, роль руля (табл. 27, 4).
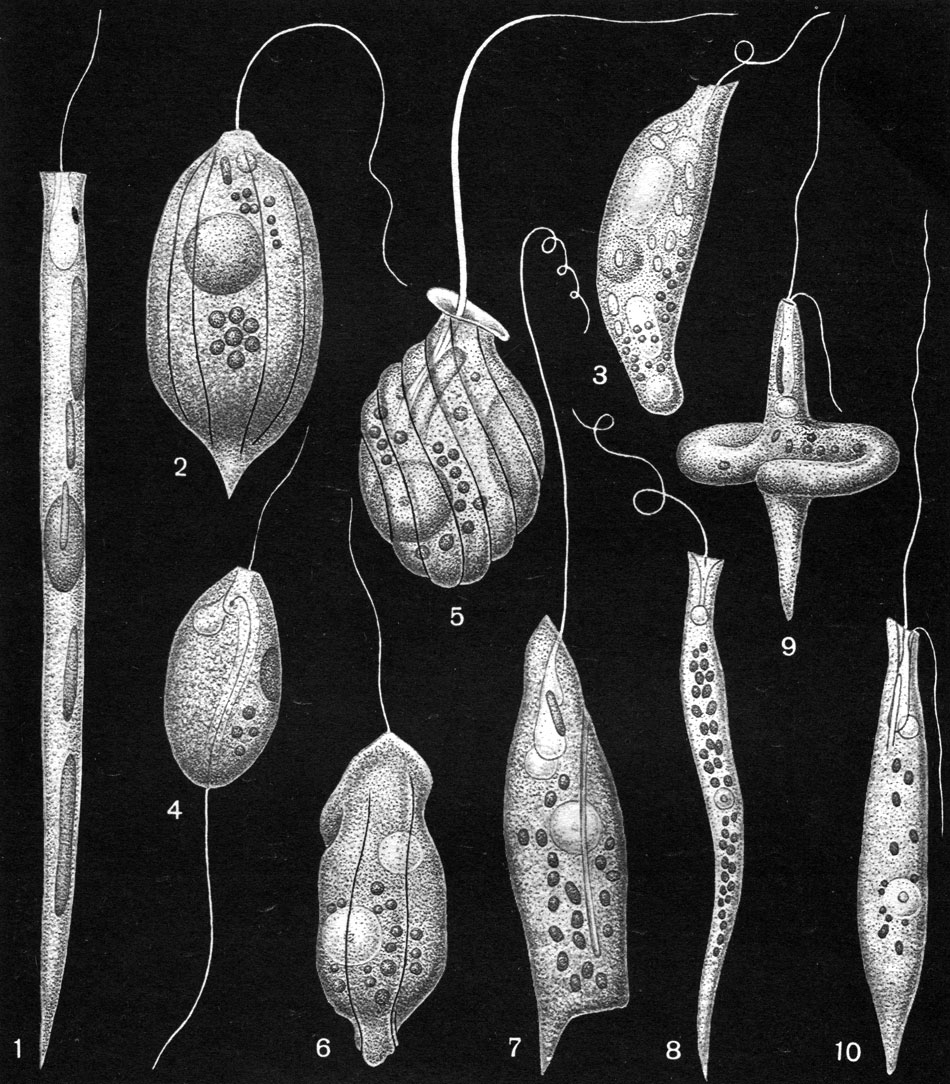
Таблица 27. Бесцветные эвгленовые водоросли: 1 - Cyclidiopsis acus; 2 - Petalomonas praegnans; 3 - Menoidium pellucidum; 4 - Anisonema prosgeobium; 5 - Urceolus cyclostomus; 6 - Urceolus platyrhynchus; 7 - Peranema pleururum; 8 - Menoidium tortuosum; 9, 10 - Heteronema acus, в двух состояниях клетки
Среди эвглен существует группа так называемых "безжгутиковых" форм. Жгут у них отсутствует в наружной своей части или значительно укорочен. Движение у этих форм ползающее, оно осуществляется за счет метаболичных изгибов тела.
Зеленый цвет эвгленовых водорослей объясняется наличием хлорофилла, который содержится у них в виде двух форм - а и b. Это те же самые формы хлорофилла, которые имеются и у высших растений. Хлорофилл - преобладающий пигмент у эвгленовых водорослей, но, кроме него, присутствуют и другие: α-, β- и ε-каротины и несколько ксантофиллов. Все эти пигменты сосредоточены в хлоропластах.
Кроме пигментов, участвующих в фотосинтезе, у некоторых эвглен (в частности, у Euglena sanguinea) имеется в большом количестве пигмент красного цвета - астаксантин. Раствор его в масле (гематохром) выполняет роль световой ширмы: регулирует количество света, попадающего на хлоропласты. В условиях интенсивного освещения пигмент скапливается в периферической части клетки и затеняет собой хлоропласты. Клетка при этом окрашивается в различные оттенки красного цвета. При ослаблении освещенности пигмент перемещается к центру клетки, и водоросли становятся ярко-зелеными.
Хлоропласты эвгленовых имеют разнообразное строение, особенно у представителей рода эвглена. Они могут быть звездчатыми (рис. 198,3), лентовидными (рис. 198, 5), крупнопластинчатыми (рис. 198, 1, 2). Звездчатый хлоропласт основной своей частью лежит в центре клетки, откуда к периферии расходятся его лучи. Но более распространены у эвгленовых мелкие дисковидные хлоропласты, расположенные в клетках постенно, т. е. такие же, что и у высших растений (рис. 198, 6).

Рис. 198. Форма хлоропластов, пиреноидов и парамилоновых зерен эвгленовых водорослей: 1 - пластинчатый хлоропласт с двускорлупчатым пиреноидом (сбоку); 2 - пластинчатый хлоропласт с пиреноидом в центре (сверху); 3 - звездчатый хлоропласт с парамилоновыми зернами в центре; 4 - хлоропласт с внутренним пиреноидом, покрытым скорлупкой парамилона; 5 - лентовидные хлоропласты; 6 - дисковидные хлоропласты; 7-12 - парамилоновые зерна
В хлоропластах у некоторых видов эвгленовых водорослей хорошо заметны более плотные образования - пиреноиды (рис. 198, 1, 2), центры образования запасного питательного вещества парамилона, производного глюкозы. Пиреноиды особенно хорошо заметны благодаря покрывающим их скорлупкам парамилона. Пиреноиды могут откладывать две или одну скорлупку парамилона в форме часовых стекол. У некоторых трахеломонад и стромбомонад известен так называемый внутренний пиреноид: он вытянут к центру клетки и покрыт тонкой наперстковидной скорлупкой парамилона (рис. 198,4). У ряда эвглен встречается "голый" пиреноид, не образующий парамилона. У форм с мелкими дисковидными хлоропластами пиреноиды обычно отсутствуют.
У видов, не имеющих пиреноидов, парамилон образуется непосредственно в цитоплазме. В водоемах с большим количеством органических веществ клетки эвгленовых обычно переполнены массой мелких округлых или удлиненных зерен парамилона. В более "голодных" условиях клетки накапливают меньше парамилона. Кроме неопределенного количества мелких зерен, в клетках эвгленовых нередко имеются одно- два более крупных парамилоновых зерна, называемых парамилиями. Парамилии свойственны обычно малометаболирующим видам. Форма парамилий, их размеры и положение в клетке постоянны для определенных видов (рис. 198, 7-12). По-видимому, парамилии повышают остойчивость организмов при движении.
У живых эвгленовых водорослей хорошо заметно красное пятно в передней части клетки - стигма (табл. 28). Стигма прилегает к спинной стороне резервуара; она состоит из мелких оранжево-красных липоидных телец, окрашенных каротином и астаксантином. Тельца плотно прилегают друг к другу, образуя пластинку. Вместе с фоторецептором - утолщенным узелком на жгуте - стигма выполняет роль светочувствительного органа. При движении организма стигма помогает регулировать положение клетки таким образом, чтобы луч света падал на хлоропласт под определенным углом. Зеленые эвгленовые водоросли обладают положительным фототаксисом, но при очень сильном освещении они удаляются в более затененные места, т. е. фототаксис становится отрицательным.
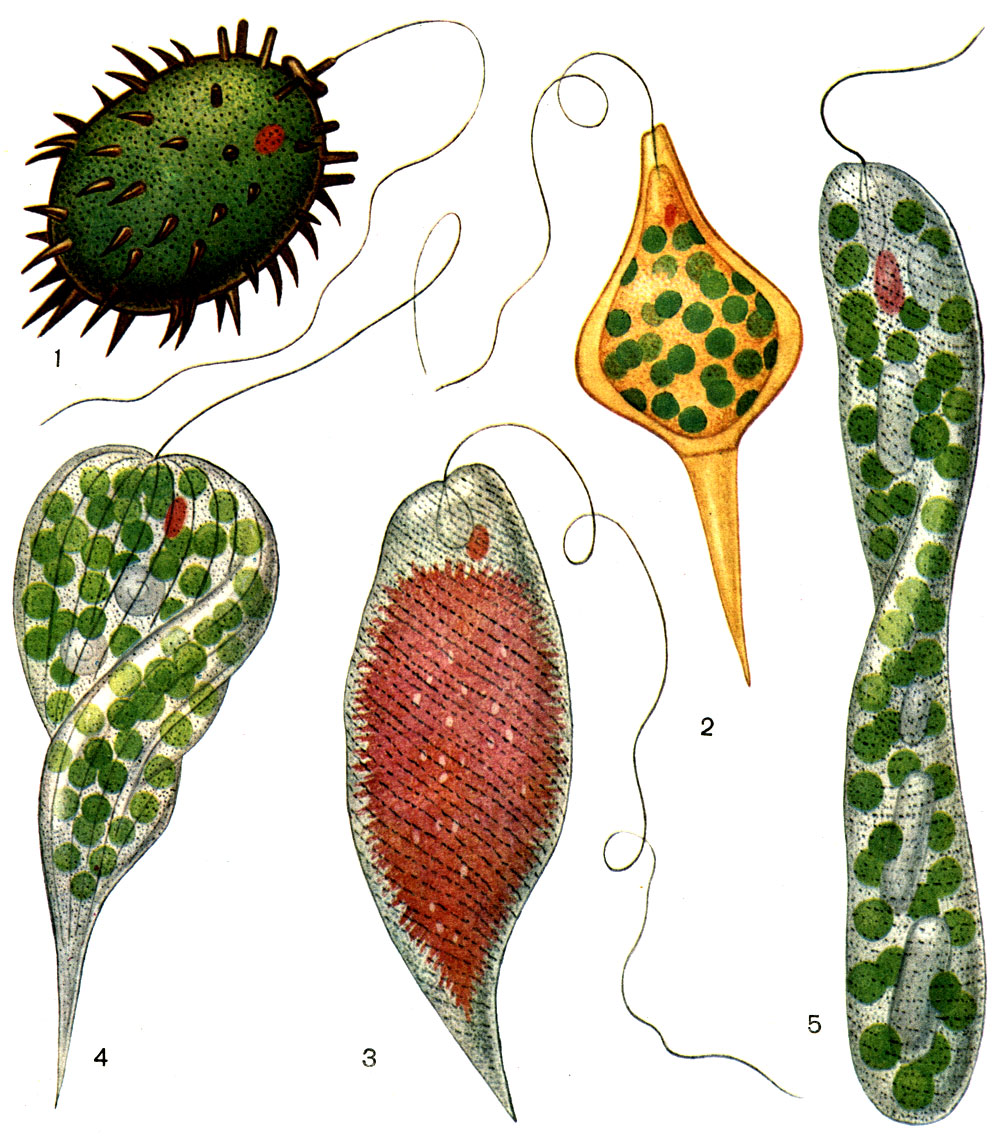
Таблица. 28. Окрашенные эвгленовые водоросли: 1 - Trachelomonas bituricensis; 2 - Strombomonas ensifera; 3 - Euglena sanguinea, заполненная гематохромом; 4 - Phacus longicauda; 5 - Euglena ehrenbergii
Клеточное ядро эвгленовых водорослей крупное, шаровидное или эллипсоидное. Оно расположено в центре клетки или сдвинуто к задне-му концу (рис. 193). У сильно вытянутых клеток ядро также вытянуто. Ядро состоит из нуклеоплазмы, содержащей хроматин, и кариосомы. В отличие от ядрышка высших растений кариосома эвгленовых не исчезает при митозе, а делится самостоятельно.
Эвгленовым водорослям свойственны все три основных типа питания живых организмов: фототрофный, сапрофитный и голозойный (анимальный). Фототрофный тип питания, т. е. усвоение углекислоты с помощью солнечной энергии, характерен для большей части представителей порядка Euglenales: эвглен, трахеломонасов, стромбомонасов, лепоцинклисов и других зеленых форм. Сапрофитный тип питания, т. е. осмотическое потребление растворенных в окружающей среде органических веществ, преобладает у бесцветных представителей этого же порядка: астазий, дистием, меноидиумов и других форм, не имеющих хлоропластов. Наконец, для родов перанема, гетеронема, энтосифон и других, объединяемых в порядок перапемовых, характерен голозойный (анимальный) тип питания: заглатывание оформленных частиц органического вещества или других более мелких организмов. Голозойное питание встречается иногда и у астазиевых, а некоторые перанемовые могут питаться целиком сапрофитно.
Многие зеленые эвгленовые водоросли, усваивая углекислоту с помощью солнечной энергии, обладают способностью одновременно поглощать и готовые органические вещества, имеющиеся в растворенном состоянии в окружающей среде, т. е. склонны к миксотрофному (смешанному) питанию. Отмечена способность эвгленовых положительно реагировать на имеющийся в растворе витамин В12 и другие ростовые вещества.
Процесс размножения эвгленовых водорослей начинается обычно вечером или рано утром. Заключается он в продольном делении особи надвое (рис. 199). Перед делением эвгленовые сбрасывают наружную часть жгута. Происходит деление ядра, жгутикового аппарата, стигмы и хлоропластов. На переднем конце клетки появляется разделительная щель, которая постепенно увеличивается. Образующиеся при этом дочерние клетки совершают активные метаболичные движения. К концу деления особи связаны лишь своими задними концами. Затем и эта связь прерывается, клетки полностью освобождаются друг от друга, вырабатывают жгуты и начинают свое активное движение.
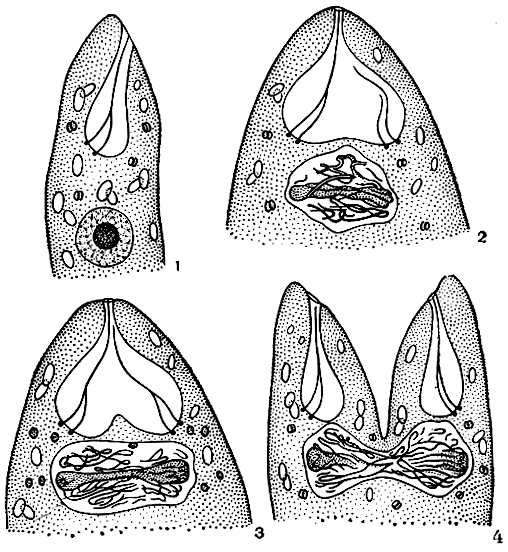
Рис. 199. Схема деления клетки эвглены: 1 - клетка перед делением; 2 - метафаза; 3 - анафаза; 4 начало продольного разделения клетки
У представителей рода колациум (Colacium), ведущих прикрепленный образ жизни, разделительная щель появляется не на переднем конце, которым они прикрепляются к субстрату, а на свободном морфологически заднем конце клетки.
Деление клетки может происходить и в так называемом пальмеллевидном состоянии (рис. 200). В этом случае оно сопровождается выделением большого количества слизи. Клетки, не образуя жгутов, вновь и вновь продолжают делиться, и в результате получается пальмелла - большое скопление неподвижных клеток, погруженных в слизь.
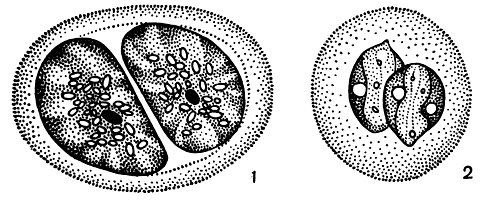
Рис. 200. Деление эвгленовых водорослей в состоянии пальмеллы: 1 - Euglena geniculata; 2 - Е. pisciformis
Процесс деления может продолжаться от 2-4 ч (у эвглен) до двух суток (у факусов).
В своем цикле развития большинство эвгленовых водорослей сохраняет подвижный образ жизни. Лишь немногие виды способны временно переходить в пальмеллевидное состояние. Преимущественно неподвижный образ жизни ведут колациум и эвгленокапса (Euglenocapsa).
У эвгленовых водорослей известны и покоящиеся стадии. У метаболичных видов это округлые клетки. У видов с постоянной формой тела покоящиеся стадии сохраняют эту же форму, лишь сбрасывают жгут и утолщают перипласт.
Эвгленовые водоросли широко распространены по всей территории земного шара. Нет такой области, нет такого водоема, где они не были бы найдены в том или ином количестве. Это, конечно, не означает, что эвгленовые безразличны к условиям внешней среды. Эвгленовые водоросли - это обитатели континентальных внутренних водоемов. В морях и океанах находки их единичны и случайны. В незначительном количестве встречаются эвгленовые в текучих водоемах и в планктоне центральной части крупных озер. Небогат состав этой группы в водоемах Арктики и пустынных областей. Излюбленными местообитаниями эвгленовых являются пресные, стоячие, мелководные, хорошо прогреваемые, заросшие водными растениями и богатые органическими веществами водоемы лесной и лесостепной зон: заболоченные лесные лужи, канавы, пруды, в особенности удобряемые рыбоводные и биологические, служащие для очистки сточных вод, прибрежные мелководные участки озер. В подобных водоемах эвгленовые водоросли играют большую роль, нередко выступая в качестве доминирующих форм и вызывая "цветение" этих водоемов.
Практическое значение эвгленовых водорослей связано прежде всего с их физиологическими свойствами. Способность большинства видов к миксотрофному или полностью сапрофитному питанию позволяет им активно участвовать в самоочищении водоемов, загрязненных органическими веществами. Среди эвгленовых водорослей существует ряд видов (Euglena pisciformis, Е. viridis, Lepocinclis ovum и др.), которые являются хорошими индикаторами степени загрязнения водоема. Большое содержание эвгленовых водорослей в водоеме всегда указывает на повышенную эвтрофность водоема. Некоторые эвгленовые водоросли, в частности Euglena gracilis, служат объектом экспериментального изучения.
Неприхотливость этого вида, способность интенсивно размножаться в условиях культуры позволили использовать его в качестве эталона для выяснения действий на живой организм физиологических, химических и других факторов: различной температуры, антибиотиков, гербицидов, витаминов, ростовых веществ и др.
С помощью эвглен, положительно реагирующих на наличие витамина В12, можно определить его содержание в различных растворах.
Отдел эвгленовых водорослей состоит из двух порядков: эвгленовых (Euglenales) и перанемовых (Peranematales). Первый порядок объединяет три семейства: эвгленовых (Euglenaceae), колациевых (Colaciaceae) и астазиевых (Astasiaceae).
Семейство эвгленовых наиболее богато представителями. Это зеленые свободноживущие формы. Сюда относится прежде всего центральный род - эвглена (Euglena, табл. 28, рис. 201, 1-4). Этот род наиболее известен, так как ряд видов эвглен обладает способностью развиваться в массе, давая упомянутое выше "цветение" воды.
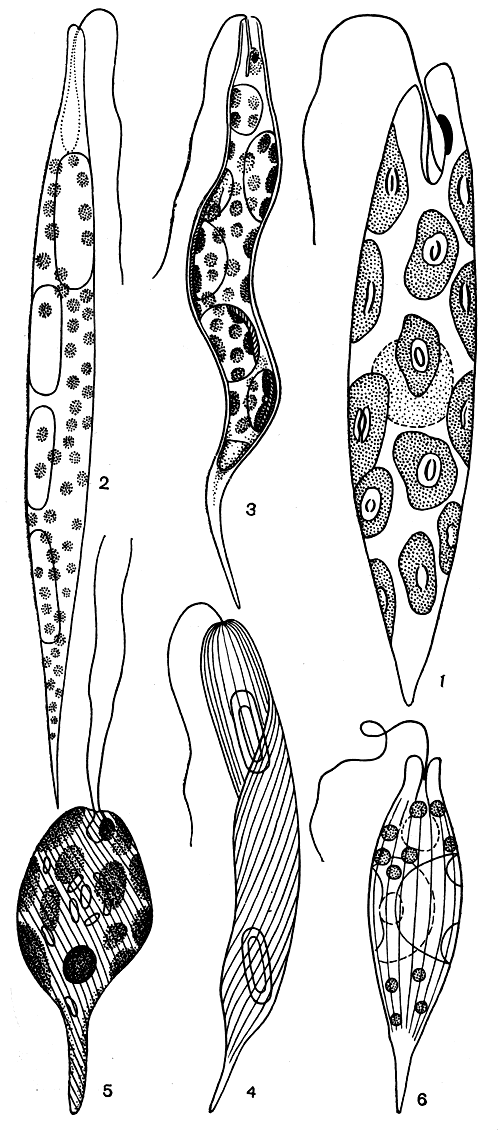
Рис. 201. Зеленые эвгленовые водоросли: 1 - Euglena gracilis; 2 - Е. acus; 3 - Е. convoluta; 4 - Е. oxyuris; 5 - Eutreptia pyrenoidifera; 6 - Lepocinclis marssonii
Близка к эвгленам эвтрепция (Eutreptia), отличающаяся от них наличием двух жгутов равной длины и более сильной метаболией. Как исключение среди эвгленовых водорослей, эвтрепция предпочитает морские или солоноватые водоемы.
Удивительно красивы и своеобразны при рассматривании под микроскопом коричневатые "домики" трахеломонасов (Trachelomonas, табл. 28, 2), стенки которых пропитаны гидроокислами железа. Строение "домика" служит характерным признаком вида. Трахеломонас хорошо развивается в водоемах с темно-бурой гумифицированной водой, богатой соединениями железа, тогда как на многие другие водоросли большое количество соединений железа производит угнетающее действие. Представители рода трахеломонас встречаются в СССР в водоемах Украинского Полесья и в южной тайге Западной Сибири, где они селятся в основном в малых лесных заболоченных водоемах.
Род лепоцинклис (Lepocinclis, рис. 201, 6) объединяет одноклеточные подвижные одножгутиковые организмы, совершенно неспособные к метаболии. По бокам клеток лепоцинклиса под микроскопом обычно очень хорошо заметны два сильно преломляющих свет пятна, благодаря которым этот род отличается от некоторых близких видов эвглен. Это парамилоновые зерна, которые у лепоцинклиса кольчатые и постенно облегают клетку изнутри. Лепоцинклис распространен южнее, чем трахеломонас, и часто встречается в прудах и других водоемах лесостепи и степи, хотя некоторые виды (например, Lepocinclis ovum) имеют повсеместное распространение.
Род факус (Phacus, табл. 28, 4; рис. 202, 1-5) морфологически наиболее сложен. Его асимметричные клетки всегда скручены и снабжены килевидными выростами на спинной стороне и длинным рулевым отростком. Факусы обычно встречаются в небольшом числе в зарослях высших водных растений, случаи массового развития их редки.
Семейство колациевых содержит только один род колациум (Colacium, рис. 202, 6). В цикле развития этих водорослей, в отличие от остальных эвгленовых, преобладает неподвижная стадия. Колациум - эпипланктонный организм, поселяющийся на мелких планктонных животных: циклопах, дафниях, коловратках. К телу животных колациум прикрепляется с помощью слизистой ножки или подушечки.
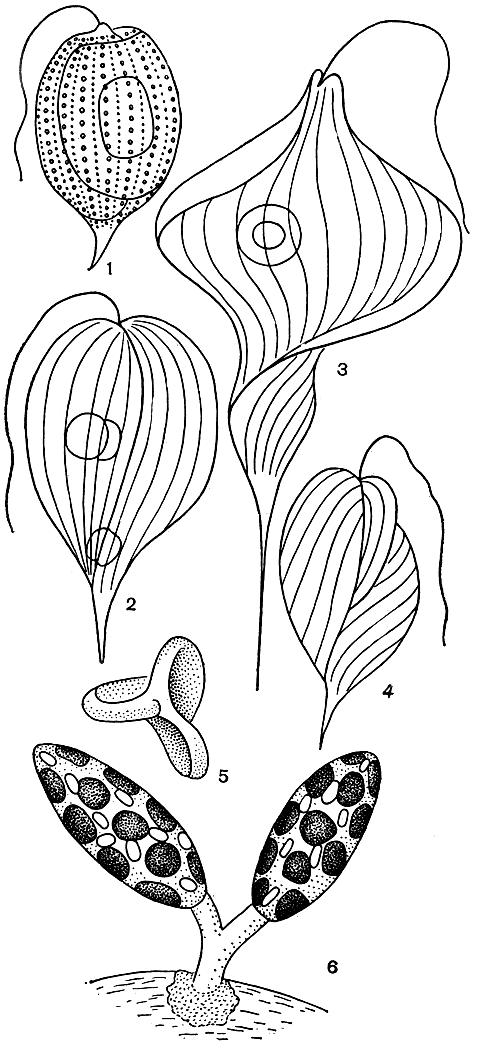
Рис. 202. Зеленые эвгленовые водоросли: 1 - Phacus monilatus; 2 - Ph. orbicularis; 3 - Ph. longicauda; 4 - Ph. arnoldii; 5 - та же клетка сверху; 6 - Colacium arbuscula
К семейству астазиевых относятся бесхлорофилльные организмы, питающиеся сапрофитно органическими веществами, растворенными в окружающей воде. Питательные вещества осмотически поступают в клетку. Сюда относятся сильно метаболирующие дистигма (Distigma, рис. 203, 1) и астазия (Astasia, рис. 203, 2), а также роды меноидиум (Menoidium, табл. 27, 3, 8), гиропаигне (Gyropaigne, рис. 203, 3), рабдомонас (Rhabdomonas, рис. 203, 4) и циклидиопсис (Cyclidiopsis, табл. 27, 1), имеющие постоянную форму тела. Все эти водоросли часто встречаются в поверхностных слоях ила в различных малых водоемах.
Порядок перанемовых включает виды, у которых процесс питания протекает по анимальному типу. Относящиеся сюда петаломонас (Petalomonas, рис. 203, 5; табл. 27, 2), энтосифон (Entosiphon, рис. 203, 6), перанема (Реrаnema, табл. 27, 7; рис. 203, 7), гетеронема (Heteronema, табл. 27, 9, 10; рис. 203, 8), анизонема (Anisonema, табл. 27, 4) и урцеолус (Urceolus, табл. 27, 5, 6) - хищники. Они заглатывают мелкие оформленные частицы органических веществ и мелкие клетки других водорослей. Однако они могут питаться и сапрофитно.
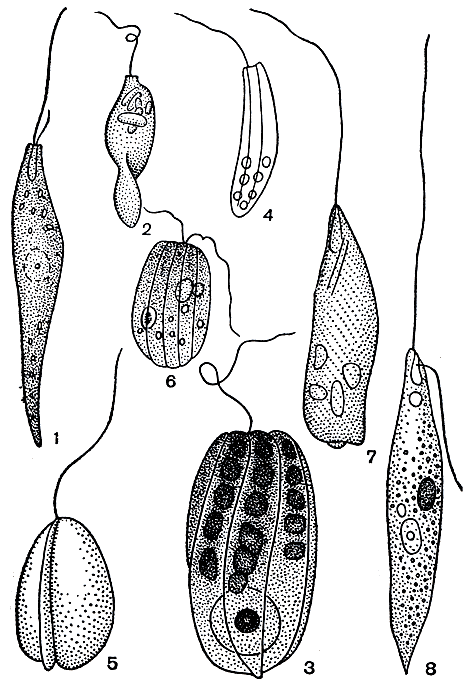
Рис. 203. Бесцветные эвгленовые водоросли: 1 - Distigma proteus; 2 - Astasia inflata; 3 - Gyropaigne cosmos; 4 - Rhabdomonas costata; 5 - Petalomonas steinii; 6 - Entosiphon sulcatum; 7 - Peranema trichophorum; 8 - Heteronema acus
В настоящее время известно около 1000 видов эвгленовых водорослей, из них для водоемов Советского Союза отмечено около половины.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'