Строение слоевища красных водорослей
При всем многообразии внешней формы красные водоросли отличаются единым планом строения слоевища - в основе его у всех многоклеточных багрянок лежит клеточная разветвленная нить. Паренхимный тип организации здесь фактически отсутствует. Единственный пример паренхимного слоевища мы находим в классе бангиевых - у порфиры и бангии.
Слоевище бангиевых. Наиболее просто устроенным слоевищем обладают представители класса бангиевых. Только здесь встречаются одноклеточные формы. Они растут в виде одиночных клеток или колониями. Самые простые колонии - это бесформенные скопления клеток, соединенные общей слизью, подобные тем, какие образует порфиридиум (Porphyridium, рис. 149, 1). Постепенно у колониальных форм намечается тенденция к нитевидному росту: клетки, заключенные в общую слизистую трубку, располагаются в один или несколько рядов.
Нитчатые колонии астероцитиса (Asterocytis), гониотрихума (Goniotrichum) бывают неразветвленными или ветвятся по типу ложного ветвления (рис. 149, 5, 4). Ложным его называют потому, что возникновение ветвей не связано с боковым ростом клеток нитей.
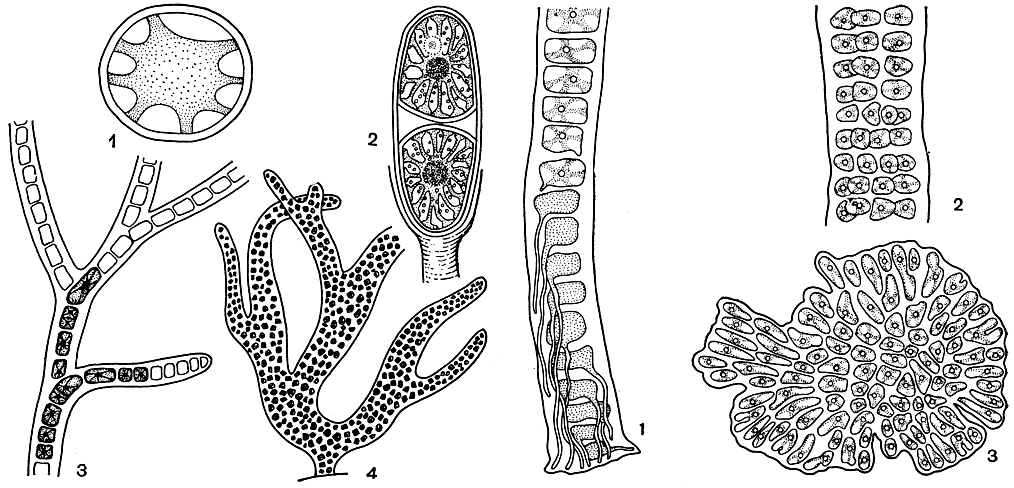
Рис. 149. Бангиевые (слева): 1 - Porphyridium; 2 - Chroothece; 3-4- однорядные и многорядные нитчатые колонии Goniotrichum
Слоевище нитчатых бангиевых может быть вертикальным или стелющимся. Стелющиеся нити обычно ветвятся и образуют на субстрате рыхлую или более или менее плотную пластину неправильных очертаний, как у эритрокладии (рис. 150, 3). Если нити расположены плотно, то такая пластина напоминает паренхимную.
Большинство бангиевых относится к прямостоячим организмам. Усложнение в строении вертикальной нити заключается в том, что клетки приобретают способность делиться в нескольких плоскостях, в результате чего однорядная нить преобразуется в многорядную, как, например, у бангии (рис. 150, 1, 2).
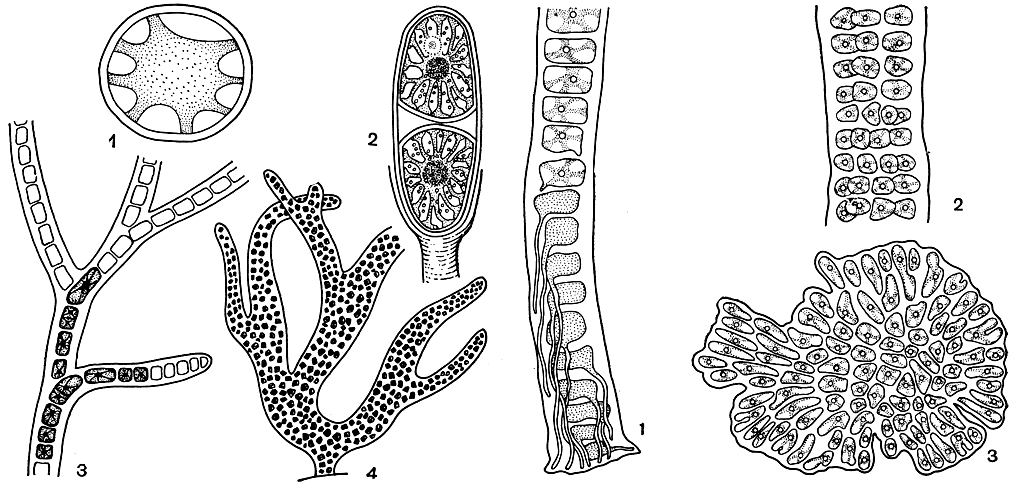
Рис. 150. Строение бангиевых (cсправа): 1,2 - Bangia (1 -основание растения с ризоидами, 2 - часть многорядной нити); 3 - Erythrocladia
Кроме нитчатых форм, среди бангиевых имеется группа водорослей пластинчатого строения. Пластинчатые формы могли образоваться в результате усложнения нитчатых. Приобретение клетками способности делиться в двух плоскостях - поперечной и продольной - привело к преобразованию однорядной нити в однослойную пластину паренхимного строения. Если деление происходит в трех плоскостях, образуется двухслойная пластина. Такое строение мы находим у порфиры, наиболее высокоорганизованной водоросли в классе бангиевых (табл. 20, 1). Но наряду с этим пластинчатое вертикальное слоевище могло возникнуть и из стелющейся структуры, подобной эритрокладии: вначале пластина становится многослойной, затем верхний клетчатый слой в ней отделяется с образованием мешковидной структуры, которая, разрываясь, растет дальше как однослойная пластина. Так формируется слоевище порфиропсиса.
Вместе с усложнением самого слоевища у вертикальных форм усложняется и строение органа прикрепления. Вначале базальная клетка почти не меняет формы и не отличается от остальных клеток, затем у нее появляются выросты, которые создают уже специальный орган прикрепления.
У наиболее высокоорганизованных форм ризоидные выросты отходят не только от базальной клетки, но и от соседних клеток слоевища (рис. 150, 1). Нередко их образуется так много, что вместе они складываются в массивную подошву, как, например, у порфиры.
Рост у бангиевых диффузный, т. е. происходит в результате деления всех клеток слоевища.
Слоевище флоридеевых. В отличие от бангиевых, все представители класса флоридеевых - многоклеточные организмы. Тело их представляет собой более или менее сложную систему разветвленных нитей. Паренхимный тип организации у них отсутствует. В наиболее простом случае слоевище состоит из свободных однорядных клеточных нитей. Эта структура свойственна немногим флоридеевым. Ею обладают примитивные представители порядка немалиевых (рис. 151, 1) и некоторые просто организованные церамиевые (рис. 182, 1-3), которые произошли, вероятно, непосредственно от первых. Водоросли, слоевище которых построено из свободных однорядных нитей,- преимущественно мелкие организмы, живущие на других водорослях и животных. Многим из них свойственно разнонитчатое строение. Стелющиеся по субстрату нити обычно отличаются по строению от вертикальных и имеют вид свободно расположенных нитей или собраны в плотные пластины. У некоторых из нитчатых форм стелющаяся часть не развивается, и они прикрепляются к грунту одной базальной клеткой.
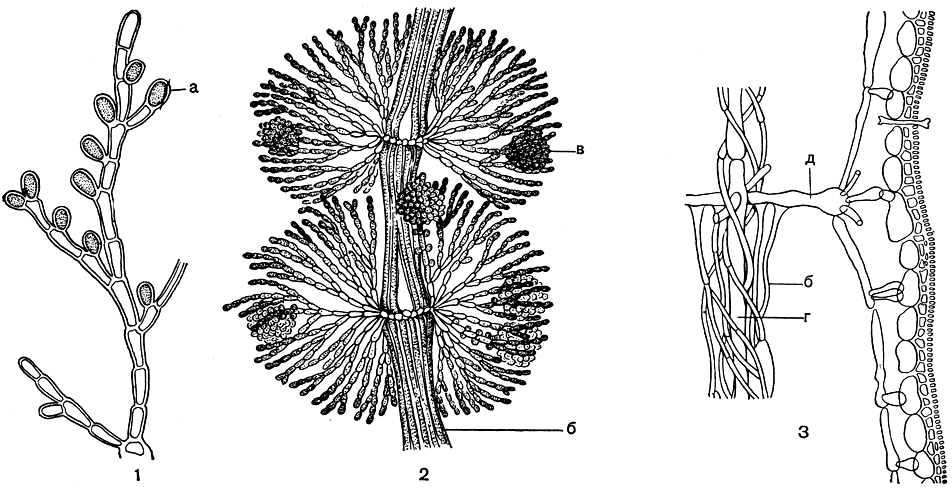
Рис. 151. Эволюция структуры флоридеевых: l - Kylinia; 2- часть слоевища Batrachospermum; 3 - продольный срез через слоевище Lemanea: а - моноспоры, б - ризоиды, в - гонимобласт, г - центральные клетки, д - перицентральная клетка
В процессе эволюции свободно-нитчатое строение претерпело ряд изменений, в результате которых сформировалась сложная анатомическая структура большинства багрянок. Первым шагом в усложнении однорядного нитчатого слоевища было возникновение большого числа коротких и обильно разветвленных боковых веточек, подобно тому, как это происходит у батрахоспермума. Нередко эти боковые веточки отходят супротивно или мутовчато, по нескольку от каждой клетки. В отличие от основных ветвей они растут в длину только до определенного предела и поэтому являются ветвями ограниченного роста. У своей вершины веточки ограниченного роста разветвляются более густо, чем в нижней части, в месте своего отхождения (рис. 151, 2). У многих багрянок с таким строением слоевище сильно пропитывается студенистым веществом, которое скрепляет все многочисленные веточки, в результате чего формируется компактное слоевище с довольно рыхлым строением, имеющее вид слизистых шнуров. В дальнейшем по мере усложнения строения веточки ограниченного роста усиленно ветвятся и разрастаются таким образом, что смыкаются друг с другом, образуя вокруг центральной части плотный периферический слой. При этом студенистое вещество теряет значение скрепляющего ветви материала и развивается в незначительной степени (рис. 151, 3). Таким образом формируется кора багрянок. При плотном строении только клетки, расположенные у поверхности слоевища, способны воспринимать свет и участвовать в фотосинтезе и ассимиляции. В отличие от внутренних клеток слоевища, они интенсивно окрашены и имеют более мелкие размеры, так как представляют собой конечные разветвления ветвей ограниченного роста. Этот ассимиляционный слой часто называют внешней корой, тогда как глубже расположенные части веточек ограниченного роста называют внутренним коровым слоем.
Кора всех багрянок имеет одно происхождение, но в процессе эволюции у разных форм она приобрела свои морфологические особенности, которые складываются из размеров клеток, слагающих ветви ограниченного роста, степени их разветвленности, плотного или рыхлого расположения ветвей. У более высокоорганизованных форм коровые нити укорачиваются и плотнее прилегают друг к другу, так что общее строение корового слоя упрощается и становится похожим на паренхимное. Вследствие этого первоначальная нитчатая структура различается с большим трудом. И только у вершины слоевища, вблизи точки роста, можно увидеть, как центральные осевые клетки отчленяют от себя боковые клеточные нити, не сообщающиеся с соседними, а только близко примыкающие к ним (рис. 152, 1, 2).
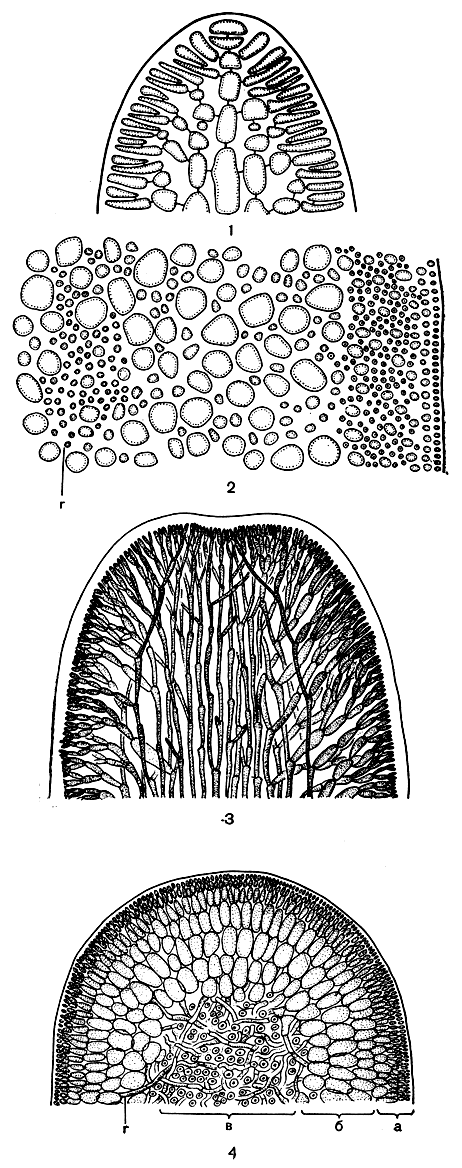
Рис. 152. Строение Gelidium (2, и Furcellaria (5, 4): 1 - продольный срез через вершину веточки Gelidium; 2 - поперечный срез слоевища Gelidium; 3 - продольный срез через вершину Furcellaria; 4 - поперечный срез слоевища Furcellaria: а - внешняя кора, б - внутренняя кора, в - пучок центральных нитей, г - ризоиды
Сложная анатомическая структура многих красных водорослей создается в значительной степени за счет особых ризоидных нитей. Уже у форм со свободным нитчатым строением можно наблюдать, как от нижних клеток ветвей ограниченного роста при их возникновении отходят ризоидные нити, или гифы, имеющие вид узких клеток с толстой оболочкой, которые растут не в бок, а тянутся вдоль осевых нитей по их поверхности (рис. 151, 2, 3). В дальнейшем по мере усложнения организации слоевища степень развития ризоидов увеличивается. Это можно проследить у багрянок и с рыхлым и с плотным строением. Нередко они развиваются настолько, что заполняют всю внутреннюю часть слоевища вокруг основных осевых нитей и клеток внутренней коры (рис. 152, 2; 153, 2).
В анатомическом строении флоридеевых различают два основных типа - одноосевой и многоосевой. При одноосевом типе, или, как его еще называют, центральнонитчатом, в основе строения слоевища лежит единственная однорядная клеточная нить с боковыми веточками ограниченного роста (рис. 151; 152, 1, 2). При многоосевом типе центральная ось слоевища состоит из пучка параллельно идущих нитей. Их расположение напоминает струи воды в фонтане, поэтому этот тип организации называют также фонтанным (рис. 152, 3, 4). Нити пучка могут располагаться плотно в центре слоевища или кольцом, и тогда в центре остается полость, как, например, у родимениевых. Каждая из нитей центрального пучка растет посредством апикальной клетки и отчленяет в бок веточки ограниченного роста.
Тот или иной тип организации характерен для определенных групп водорослей. В одних случаях одинаковое строение имеют представители целых порядков. Все родимениевые многоосевые, тогда как порядки церамиевых и гелидиевых построены по одноосевому типу. В остальных порядках представлены оба типа строения, и здесь характер строения выступает уже как признак семейства. Иногда, как показали наблюдения за развитием некоторых водорослей, растения одного и того же вида формируются как по одноосевому, так и по многоосевому типу.
В основе строения многих флоридеевых лежит разнонитчатая организация. Более всего она выражена у нема лиевых и криптонемиевых. При прорастании спор у водорослей этих порядков вначале образуется стелющаяся структура, от которой позднее возникают более или менее многочисленные вертикальные нити. Позднее из отдельных вертикальных нитей при одноосевом строении и из группы нитей при многоосевом формируется слоевище этих багрянок.
Разнонитчатую организацию можно различить, хотя и в несколько измененной форме, также в порядках гигартиновых и родимениевых. Только у церамиевых стелющаяся часть полностью отсутствует. По-видимому, разнонитчатая структура представляет собой самый примитивный тип организации, и в процессе эволюции наибольшее развитие получает система вертикальных нитей, тогда как стелющаяся часть постепенно редуцируется. Такое же направление морфологической эволюции показывают и бурые водоросли.
В отличие от бангиевых, у которых, как уже говорилось, в росте в длину участвуют все клетки, флоридеевые характеризуются апикальным ростом. Осевые нити, из которых слагается слоевище флоридеевых, растут в длину в результате деятельности верхушечной, или апикальной, клетки. У многих багрянок верхушечную клетку можно отличить от остальных по куполообразной форме и более крупным размерам, тогда как у некоторых других она не отличается по внешнему виду от нижележащих.
У багрянок, построенных по одноосевому типу, имеется одна апикальная клетка, при фонтанном типе организации их несколько. Апикальные клетки делятся поперечными перегородками и отчленяют вниз один ряд клеток, которые по своему положению в слоевище являются центральными. Апикальные клетки некоторых высокоорганизованных багрянок имеют две или три отчленяющиеся поверхности. В этом случае при делении клеточные перегородки располагаются косо по отношению к продольной оси, и от одной апикальной клетки отчленяется уже не один, а два-три ряда сегментов. Образовавшиеся центральные сегменты отделяют вбок две или четыре клетки, которые располагаются вокруг них или двусторонне. Это так называемые перицентральные клетки, которые у большинства флоридеевых функционируют как апикальные клетки боковых ветвей ограниченного и неограниченного роста. Отчленив от себя перицентральные клетки, центральные в дальнейшем уже не подвергаются делению ни поперечными, ни продольными перегородками и их рост осуществляется только за счет вытягивания в длину (рис. 152, 1). Исключение из общего правила составляют некоторые кораллиновые и делессериевые, у которых встречаются интеркалярные клеточные деления. В результате такого способа роста внутренние клетки слоевища большинства багрянок, даже с плотным псевдопаренхимным строением, обычно длинные и крупные, в то время как клетки, расположенные ближе к периферии, становятся короче и мельче.
Часто бывает весьма легко определить, по какому типу - одноосевому или многоосевому - построено слоевище тех или иных багрянок. Но иногда это сделать почти невозможно, так как в построении слоевища принимает участие особая ткань - меристема. Иными словами, кроме апикальных, в росте слоевища участвуют многие клетки. У форм с плотным слоевищем в качестве меристемы функционирует обычно поверхностный слой внешней коры, образованный апикальными клетками веточек ограниченного роста. У видов с цилиндрическим слоевищем действует верхушечная меристема, у уплощенных форм - краевая. Участие в росте меристемы, по-видимому, следует считать вторичным явлением, возникшим на основе одноосевого и многоосевого типов строения. Кроме того, первоначальная структура может сильно затемняться благодаря развитию внутри слоевища многочисленных ризоидных нитей (рис. 153, 1, 2).
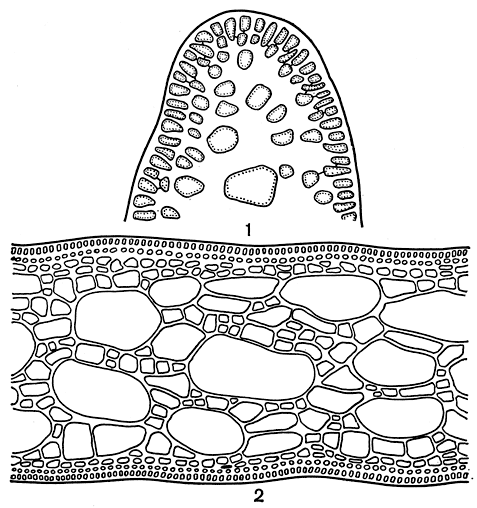
Рис. 153. Строение Callophyllis: 1 - вершина веточки; 2 - поперечный срез слоевища
Процесс формирования слоевища - порядок отчленения клеток, количество перицентральных клеток и т. д.- отличается большой сложностью, но у разных групп багрянок характеризуется постоянством и специфичностью, поэтому принцип организации слоевища в каждом порядке целесообразно рассмотреть отдельно.
Говоря о строении органов прикрепления флоридеевых, следует различать их первичную и вторичную структуру. Первичный орган прикрепления формируется на начальных этапах прорастания, тогда как вторичный характеризует уже взрослое, сформированное растение. В его создании участвуют многие клетки нижних частей слоевища, и он может иметь совсем другое строение по сравнению с первичным. У форм разнонитчатого строения роль органа прикрепления играет стелющаяся структура, состоящая, как известно, из рыхлорасположенных нитей или из плотной клеточной пластины. Когда стелющаяся часть не развивается, базальная клетка проростка образует вниз выросты, или первичные ризоиды. Из них развиваются многообразные структуры, служащие для прикрепления взрослых растений. Многие флоридеевые, особенно среди церамиевых, прикрепляются одноклеточными, короткими или длинными, ризоидами, расширяющимися при соприкосновении с субстратом в присоску. У других базальная часть состоит из многоклеточных побегов. Очень часто орган прикрепления представляет собой дисковидную подошву, имеющую клеточное или ризоидное строение. Весьма распространено у флоридеевых образование стелющихся побегов, напоминающих столоны высших растений, с многочисленными выростами-присосками, которыми растение прикрепляется к субстрату.
Дополнительные ризоиды могут развиваться и на вертикальных ветвях, особенно в условиях, где требуется усиленное прикрепление, например на прибойных скалах.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'