Размножение и цикл развития бурых водорослей
У бурых водорослей встречаются все формы размножения: вегетативное, бесполое и половое. Вегетативное размножение происходит при случайном отделении ветвей от слоевища, и только у видов рода сфацелярия для этого существуют специальные почки (рис. 131, 2-4). Слоевища бурых водорослей, оторванные от грунта, или их части не могут прикрепиться к твердому грунту: у них не образуются органы прикрепления. Такие слоевища сносятся течениями в спокойные места с песчаным или илистым дном, и там они растут в неприкрепленном состоянии. Органы бесполого и полового размножения образуются на них лишь в том случае, если их развитие к моменту отрыва слоевища от грунта уже началось. В результате такие слоевища размножаются только вегетативным путем. Старые нижние части отмирают и разрушаются, а более молодые ветви становятся самостоятельными растениями.
Бесполое размножение бурых водорослей осуществляется зооспорами, только у диктиотовых имеются неподвижные тетраспоры, а у тилоптеридовых существуют моноспоры.
Половой процесс у бурых водорослей представлен изогамией, гетерогамией и оогамией, гетерогамия встречается реже двух других форм полового размножения.
Зооспоры и подвижные гаметы бурых водорослей содержат по одному хлоропласту, который в мужских гаметах при оогамии может быть бесцветным. Глазок у подвижных клеток бурых обнаруживается не всегда.
Образование зооспор и гамет у бурых водорослей происходит во вместилищах двух основных типов: одногнездных (рис. 121, 2) и многогнездных (рис. 128, 1 а). Многогнездные вместилища могут функционировать как спорангии и как гаметангии. Внешне они при этом не отличаются так же, как зооспоры и гаметы. Одногнездные вместилища чаще бывают спорангиями. Мейоз у бурых водорослей происходит при образовании спор в одногнездных спорангиях, лишь у циклоспоровых он приходится на момент образования гамет.
Одногнездные и многогнездные вместилища часто неправильно называют соответственно одноклеточными и многоклеточными. И те и другие могут развиваться из одной и из многих клеток. Когда в однорядных слоевищах группа клеток превращается в одногнездные спорангии, то говорят о цепочке одногнездных спорангиев. Каждый из них при созревании открывается самостоятельным отверстием. В случае многогнездных вместилищ цепочка исходных клеток, каждая из которых увеличивается в размерах, дает единое многоклеточное многогнездное вместилище (рис. 122, 1, 2). После образования в нем камер можно заметить, что некоторые поперечные перегородки толще других - это перегородки материнских клеток. При созревании содержимое такого вместилища выходит через одно отверстие на вершине. У некоторых бурых водорослей одноклеточные многогнездные вместилища узкие и камеры расположены в них в один ряд. Такие образования называют однорядными многогнездными спорангиями (гаметангиями, рис. 123). Многогнездные вместилища с гнездами, расположенными в несколько рядов, считаются многорядными.
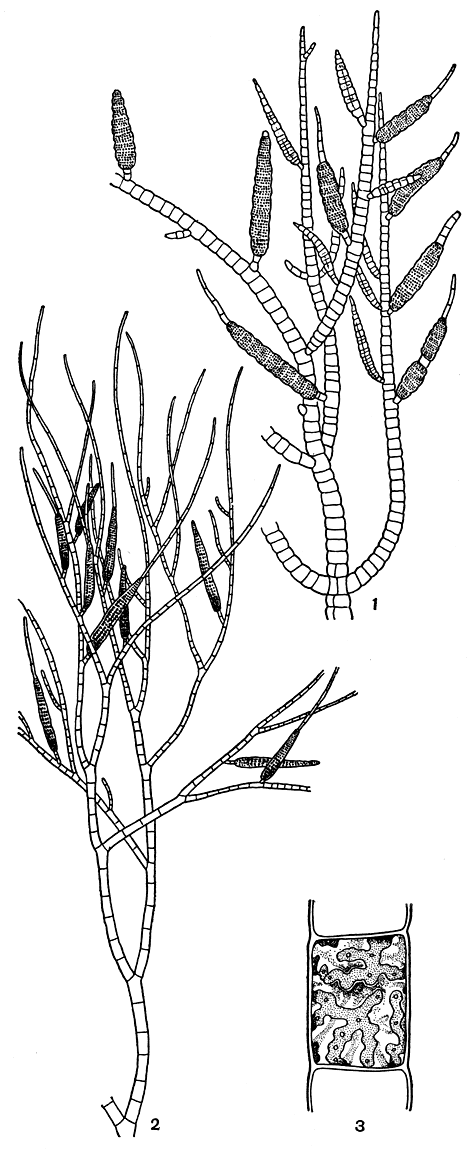
Рис. 122. Пилайелла прибрежная (Pylaiella litoralis) с многогнездными спорангиями (1); эктокарпус стручковатый (Ectocarpus siliculosus) с многогнездными спорангиями (2); клетка эктокарпуса с лентовидными хлоропластами (3)
Большинство бурых водорослей встречается в виде двух самостоятельных форм развития - спорофита и гаметофита или гаметоспорофита, которые могут быть как сходного размера и строения, так и разного. Другими словами, у бурых водорослей существуют изоморфная и гетероморфная смены форм развития. При гетероморфной смене форм развития одна из них бывает микроскопической, чаще всего это гаметофит. Развитию макроскопических слоевищ бурых водорослей нередко предшествует образование стелющихся нитей или дисков, на которых потом начинает расти вертикальное макроскопическое слоевище. Подобные образования называют протонемой. Если такое стелющееся образование несет органы размножения и со временем производит вертикальное слоевище, его называют плетизмоталлюсом.
У наиболее примитивных бурых (пунктариевые, хордариевые, эктокарповые) строгое чередование форм развития отсутствует. Из спор, производимых спорофитом, могут развиваться как слоевища гаметофитов или гаметоспорофитов, так и слоевища спорофитов.
У бурых водорослей с половым процессом в виде изогамии подвижные клетки (зооиды), которые образуются на гаметофитах, функционируют или как гаметы, сливаясь попарно и воспроизводя спорофиты, или как зооспоры, производя снова слоевища гаметофитов. Такое бесполое размножение гаметофитов у представителей вышеперечисленных порядков бурых водорослей распространено крайне широко. Иногда в лабораторных культурах наблюдается подряд несколько поколений "гаметофитов", не приступающих к половому размножению. Таким образом, они проявляют себя как спорофиты и как гаметофиты, и их лучше называть гаметоспорофитами, чтобы не путать с другими растениями, гаметофиты которых не способны к бесполому размножению спорами.
У ряда других бурых водорослей спорофит и гаметофит строго чередуются, особенно у десмарестиевых, ламинариевых и циклоспоровых. Зиготы десмарестиевых и ламинариевых развиваются в слоевища спорофитов, оставаясь прикрепленными к оболочкам оогониев. Это исключает напрасную гибель женских гамет во время нахождения их в толще воды и поиска места для прикрепления. Для класса циклоспоровых характерно развитие гаметофитов в слоевищах спорофитов. Некоторые циклоспоровые, например саргассовые, размножаются проростками спорофитов, которые образуются в результате оплодотворения и прорастания женских гамет на материнском растении.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'