Биологические особенности диатомовых водорослей
Образование колоний. Колонии у диатомовых водорослей бывают различных размеров и формы, это зависит как от количества общей слизи, так и от способа соединения клеток друг с другом (рис. 95). Образуются колонии всегда из одной клетки в результате последовательных и многократных делений ее самой и всех остальных возникающих при этом клеток. Все клетки остаются самостоятельными, и распад колонии не приводит к их гибели. Соединяются клетки при помощи слизи, выростов, шипиков, щетинок, рогов и пр. Колониальные формы обитают как в планктоне, так и в бентосе на различных субстратах - на растениях и животных, на каменистых, песчаных и илистых грунтах, на технических сооружениях и других предметах, введенных в воду человеком.
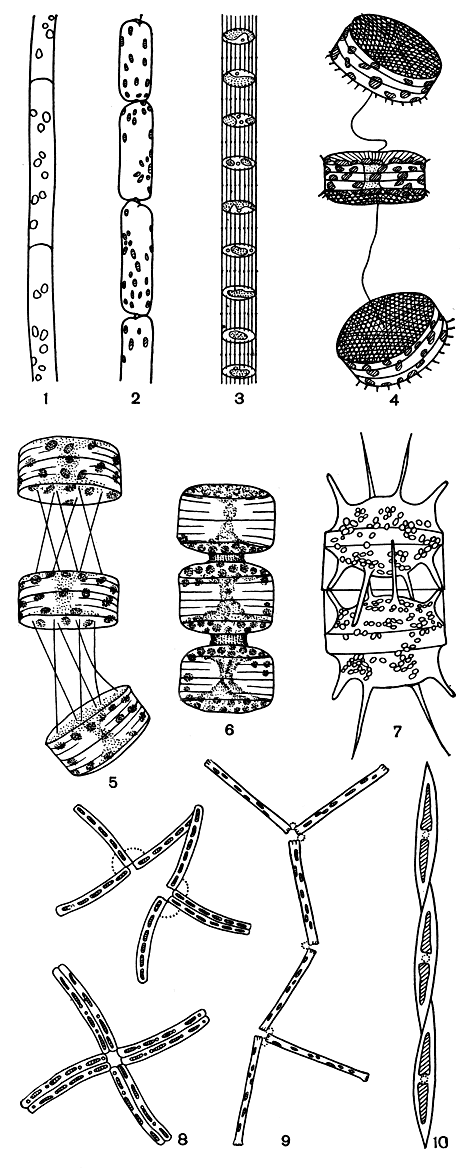
Рис. 95. Типы колоний: 1 - нитевидная колония (Leptocylindrus danicus); 2 - цепочковидная колония (Rhizosolenia fragilisslma); 3 - цепочковидная колония (Sceletonema costaturn); 4 - гибкая цепочка (Thalassiosira excentrica); 5 - гибкая цепочка (Thalassiosira polychorda); 6 - прямая цепочка (Porosira glacialis); 7 - прямая цепочка, образованная при помощи выростов (Biddulphia mobiliensis); 8 - зигзаговидная цепочка (Thalassionema nitzschioides); 9 - зигзаговидная цепочка (Diatoma elongatum); 10 - нитевидная колония (Nitzschia seriata)
В слизистых колониях клетки целиком погружены в выделяемую ими самими слизь. Эти колонии часто представлены бесформенными комочками или пленками однородной слизи, в которой клетки расположены беспорядочно. Клетки здесь могут быть подвижными, под микроскопом это создает впечатление движения всей колонии. Есть также вполне оформленные слизистые колонии, имеющие вид простых ветвистых трубок или листовидных пластинок, которые обычно прикрепляются к субстрату, реже свободно плавают. Они могут достигать макроскопических размеров. Слизь в подобного рода колониях дифференцирована на наружный плотный слой и внутренний слой жидкой консистенции, в котором живут и двигаются клетки.
Форма колоний, не имеющих общей слизи, зависит от способа соединения клеток и очертаний створок. Нитевидные колонии образуются в том случае, если смежные клетки цилиндрические, а створки их круглые, причем клетки соединены поверхностью створок (род Leptocylindrus, рис. 95, 2), мелкими шипиками (род Melosira) или специальными трубковидными выростами (роды Detonula, Stephanodiscus, Cyclotella). В лентовидных колониях клетки уплощенные, со створками линейной формы; соединяются поверхностью створок посредством слизи или мелких шипиков, расположенных по краю створок (роды Fragilaria, Rhabdonema, Achnanthes, Cymatosira, Dimerogramma). Особенно интересны колонии бациллярии парадоксальной (Bacillaria paradoxa): благодаря скользящему движению отдельных клеток по продольной оси створки в обе стороны, ее колония меняет форму от широкой ленты до длинной ступенчатой нити. Если клетки клиновидные и имеют булавовидные створки, то колония приобретает вееровидную форму (роды Licmophora, Meridion, Opephora, табл. 10, 8).
Особенно распространены цепочковидные колонии разнообразной формы. В такие колонии клетки объединяются при помощи слизистых подушечек или тяжей, коротких или длинных трубковидных выростов, рогов, щетинок и шипиков (табл. 10, 7; рис. 95, 2-7). Клетки, соединенные одним или несколькими довольно длинными слизистыми тяжами, образуют гибкие и непрочные цепочки (род Thalassiosira, некоторые виды Cyclotella). Прямые, часто очень длинные и прочные цепочки образуются путем соединения клеток посредством трубковидных выростов - длинных (род Sceletonema) или коротких (роды Bacterosira, Lauderia), а также щетинок (род Chaetoceros), длинных шипов (род Stephanopyxis), мелких шипиков (Rhizosolenia fragilissima) или рогов (роды Hemiaulus, Biddulphia). В таких цепочках соседние клетки обычно расположены на некотором расстоянии друг от друга, что повышает плавучесть колонии. При свойственном пеннатным диатомеям соединении клеток в колонии попеременно разными углами прямоугольного панциря образуются зигзаговидные цепочки (роды Tabellaria, Grammatophora, Thalassionema, Diatoma и др., рис. 95, 8, 9), а если соединение происходит с помощью двух углов одного конца створки, то получается звездчатая колония (роды Asterionella, Thalassiothrix). Клетки в подобных колониях скрепляются слизью, выделяемой специальными слизевыми порами, расположенными на полюсах створки.
Прикрепленные формы часто образуют своеобразные пучковидные или кустиковидные колонии. При этом исходная клетка сначала прочно прикрепляется одним концом к субстрату при помощи слизи, которая выделяется слизевой порой на базальном конце створки и образует слизистую подошву. В процессе последующих делений клеток общая слизистая подошва разрастается и разветвляется, благодаря чему возникает кустиковидная колония с клетками, сидящими на концах студенистых "веточек" (роды Licmophora, Rhoicosphenia, Gomphonema, Cymbella, Didymosphenia, табл. 10, 8). Если такого разрастания слизи не происходит, то клетки сидят пучками непосредственно на слизистой подошве (роды Synedra, Licmosphenia). Иногда эти колонии достигают макроскопических размеров.
Совершенно уникальна колония Coenobiodiscus muriformis, состоящая из одного слоя в 200-500 клеток, соединенных в поясковой зоне перегородками органической природы. Эти колонии воспроизводятся без прохождения одноклеточной стадии.
Приспособления к планктонному образу жизни. Планктонные диатомеи - это пассивно плавающие организмы, не связанные в своем жизненном цикле с каким-либо субстратом. Основной особенностью их являются различного рода приспособления для уменьшения массы клеток и увеличения поверхности панциря, способствующие свободному парению в толще воды. Небольшая плотность клеток достигается здесь благодаря малому объему цитоплазмы, мелким хлоропластам и наличию в протопласте значительного количества капелек масла. Интересно, что содержание масла в клетках диатомей во много раз выше, чем в клетках других планктонных водорослей. С другой стороны, панцирь планктонных диатомей, как правило, тоньше и легче, чему бентосных форм. Он пронизан густо расположенными мельчайшими отверстиями, а форма его обеспечивает большую поверхность на единицу массы. Так, у многих видов клетки приобрели форму тонких и длинных палочек (роды Rhizosolenia, Attheya, Thalassiothrix); у других - форму низкого цилиндра большого диаметра, часто с выростами и шипиками по краю створки (роды Thalassiosira, Bacterosira, Lauderia, Planktoniella). Многие планктонные виды, особенно морские пелагические, несут на своем панцире тонкие и длинные щетинки (роды Chaetoceros, Bacteriastrum, Corethron), способствующие повышению плавучести клетки (табл. 13).

Таблица 13. Диатомовые водоросли планктона: 1 - Stephanodiscus astraea; 2 - Coscinodiscus perforatus var. celulosus; 3 - Asteromphalus robustus; 4 - Thalassiosira pseudonana; 5, 6 - Thalassiosira nordenskioldii (5 - клетка, 6 - створка); 7 - Sceletonema costatum; 8 - Actinocyclus ehrenbergii; 9 - Biddulphia mobiliensis. Микрофотографии: 1 - С. И. Генкала, 2, 3, 9 - А. И. Прошкиной-Лавренко, 4-8 - И. В. Макаровой
Наиболее эффективное приспособление к парению - это увеличение поверхности путем соединения клеток в колонию. Чем больше поверхность колонии, тем выше ее плавучесть. Кроме того, слизь, выделяемая клетками планктонных видов, легче воды; поэтому она, обволакивая клетки, также препятствует погружению отдельных клеток и колонии в целом.
Кроме морфологических приспособлений к планктонному образу жизни, диатомеи обладают еще физиологической адаптацией, связанной с процессами ассимиляции. Мертвые клетки и покоящиеся споры, когда ассимиляция отсутствует, теряют плавучесть, вследствие чего опускаются на дно.
Способы передвижения. Многие пеннатные диатомовые водоросли способны к активному движению. Они передвигаются по-разному: толчками вперед, назад или слегка в сторону, поворотами с боку на бок и медленным вращением вокруг своей оси. Особый тип движения наблюдается у бациллярии парадоксальной (Ваcillaria paradoxa), палочковидные клетки которой, образующие лентовидную колонию, способны перемещаться скользящими движениями одна относительно другой в направлении продольной оси панциря.
Большинство диатомовых водорослей передвигается по субстрату, хотя некоторые движутся и в толще воды. Подвижные диатомеи, как правило, снабжены швом, причем чем сложнее устройство шва, тем совершеннее их движение. Но все же механизм движения до настоящего времени окончательно не объяснен.
Существует мнение, что клетки диатомовых водорослей скользят по субстрату благодаря плазматическому потоку в щелевидном шве панциря. Однако движение наблюдалось и у видов, имеющих очень короткий шов на конце створок без центрального узелка, а также в тех случаях, когда клетки были обращены к субстрату поясковой стороной или не соприкасались с опорой, как, например, при вращательном движении.
Одна из причин, вызывающих движение диатомей,- фототаксис. Некоторые виды обладают положительным фототаксисом, другие - отрицательным. Бентосные диатомеи, обитающие на дне, при сильном освещении погружаются в иловую пленку, а при благоприятном для них освещении перемещаются на ее поверхность. Некоторые из них по-разному реагируют на различные цвета спектра. Так, например, Navicula radiosa относится положительно к красным лучам спектра и отрицательно к голубым. Кроме того, на движение диатомовых водорослей отрицательно действуют различные вещества, способные растворять слизь.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'