Размножение диатомовых водорослей
Деление. Чаще всего диатомеи размножаются вегетативным делением клетки на две половины; этот процесс обычно происходит ночью или на рассвете. Темпы деления различны у разных видов и могут меняться даже у одного вида в зависимости от сезона или условий окружающей среды. Весной и в начале лета наблюдается максимальное развитие диатомовых в результате их интенсивного деления. Наличие в воде биогенных веществ способствует делению и росту диатомей.
Опыты показали, что в культуральной среде некоторые планктонные виды могут делиться до 3-8 раз в сутки. Бентосные виды делятся гораздо реже - один раз в 4 дня. Известны случаи еще более редкого деления - один раз за 25 дней. Но эти сведения не абсолютны, и темпы деления могут меняться в зависимости от широтного расположения водоема, его физико-химического режима и, конечно, от особенностей вида.
Процесс деления клетки у диатомей своеобразен из-за наличия твердого панциря. Сначала в протопласте начинают скапливаться капельки масла, а сам протопласт значительно увеличивается в объеме, вследствие чего эпитека и гипотека панциря расходятся, оставаясь соединенными только краями своих поясковых ободков. Протопласт делится на две равные части, а вместе с ним и хлоропласты. Если хлоропласт один, то он делится пополам; если их много, то в дочерние клетки сначала попадает половина их, а затем они делятся, в результате в дочерних клетках образуется исходное количество хлоропластов. Ядро делится митотически, часто с хорошо заметными хромосомами и центросомой на каждом из образовавшихся полюсов. После окончательного разделения клетки на две каждая из дочерних клеток, получившая лишь половину материнского панциря, сразу же "достраивает" недостающую половину, но обязательно внутреннюю, т. е. гипотеку. Дополнительные образования панциря - вставочные ободки, септы и другие структурные элементы - возникают вскоре после сформирования новой гипотеки. Таким образом, возникшие в результате деления две дочерние клетки оказываются по размерам несходными: одна клетка, получившая эпитеку, сохраняет размеры материнской клетки, а другая, получившая материнскую гипотеку, ставшую в новой клетке эпитекой, приобретает меньшие размеры. В результате после многократных делений происходит постепенное уменьшение размеров клеток у половины каждой данной популяции: у центрических диатомей уменьшается диаметр клеток, а у пеннатных - длина и отчасти ширина клеток. Установлено, что у некоторых видов в процессе делений размеры клеток уменьшаются почти в 3 раза по сравнению с первоначальными.
Микроспоры. У многих планктонных диатомей обнаружены так называемые микроспоры - мелкие тельца, возникающие в клетках в количестве от 8 до 16 и более, а у некоторых видов их бывает и более 100. Наблюдались микроспоры со жгутиками и без них, с хлоропластами и бесцветные. Наиболее часто микроспоры развиваются у видов рода хетоцерос (Chaetoceros), и наблюдалось даже их прорастание (рис. 90).
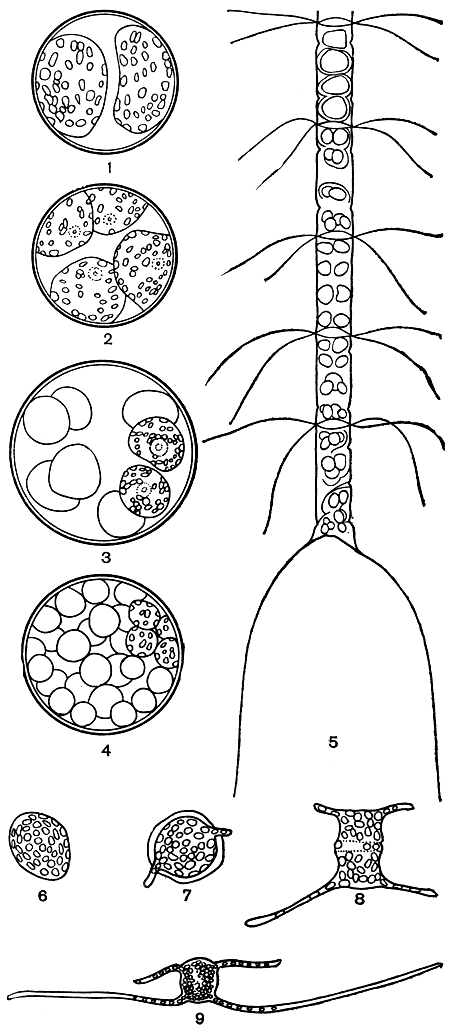
Рис. 90. Образование и прорастание микроспор: 1-4 - последовательные стадии образования микроспор у Coscinodiscus jonesianus; 5 - различные стадии образования микроспор у Chaetoceros curvisetus; 6-9 - последовательные стадии прорастания микроспор у Ch. danicus
Процесс образования микроспор цитологически не изучен, и природа их точно не установлена.
Половой процесс и образование ауксоспор. Ауксоспорами, т. е. "растущими спорами", называют такие, которые при своем образовании сильно разрастаются и затем прорастают в клетки, резко отличающиеся по размерам от исходных. Способность образовывать ауксоспоры свойственна только диатомовым водорослям, но до сих пор не удалось еще полностью объяснить этот процесс и порождающие его причины. Образование ауксоспор, по всей вероятности, вызывается различными причинами. Согласна наиболее распространенному мнению, оно наступает вследствие многократных делений, приводящих, как это описано выше, к мельчанию клеток. Достигнув минимальных размеров, клетки развивают ауксоспоры, что приводит к восстановлению их размеров. Однако другие исследователи считают, что ауксоспорообразование связано просто со старением клеток, так как его нередко удавалось наблюдать и тогда, когда клетки еще не достигли своих минимальных размеров. С этих позиций ауксоспорообразование рассматривается как процесс "омоложения" клетки. Кроме того, есть наблюдения, свидетельствующие о развитии ауксоспор при изменении условий окружающей среды, например при резком понижении температуры воздуха или воды.
Каковы бы ни были причины, способствующие возникновению ауксоспор, установлено главное: ауксоспорообразование всегда связано с половым процессом. У диатомовых водорослей встречаются все три типа полового процесса, вообще известные у водорослей,- изогамный, анизогамный и оогамный, а также некоторые формы редуцированного полового процесса (рис. 91). У пеннатных диатомей половой процесс во всех случаях состоит в сближении двух клеток, в каждой из которых створки раздвигаются и происходит редукционное деление ядра, после чего гаплоидные ядра попарно сливаются и образуется одна или две ауксоспоры. У центрических диатомей попарное сближение клеток отсутствует и ауксоспора образуется из одной клетки, в которой сначала происходит деление материнского диплоидного ядра на четыре гаплоидных ядра, два из них затем редуцируются, а два сливаются в одно диплоидное ядро и образуется ауксоспора.
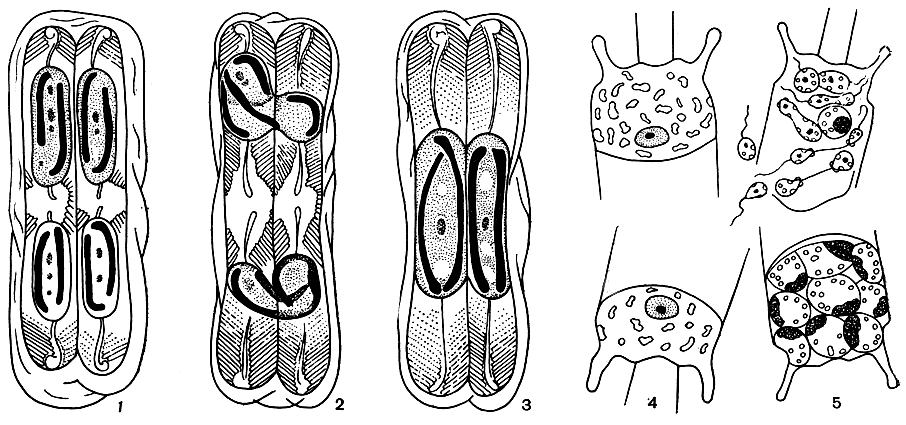
Рис. 91. Половой процесс у диастомей: 1-3 - анизогамный половой процесс у Pinnularia sp.; 4-5 - оогамный половой процесс у Biddulphia mobiliensis
Все диатомовые водоросли - диплоидные организмы, а гаплоидная фаза у них бывает только перед слиянием ядер в ауксоспоре. Как в первом, так и во втором случае после слияния ядер образуется зигота, которая сразу, без стадии покоя, резко увеличивается в размерах и развивает ауксоспору. По положению и связи с материнской клеткой ауксоспоры бывают разных типов: свободная ауксоспора, конечная, боковая, интеркалярная и полуинтеркалярная (рис. 92).
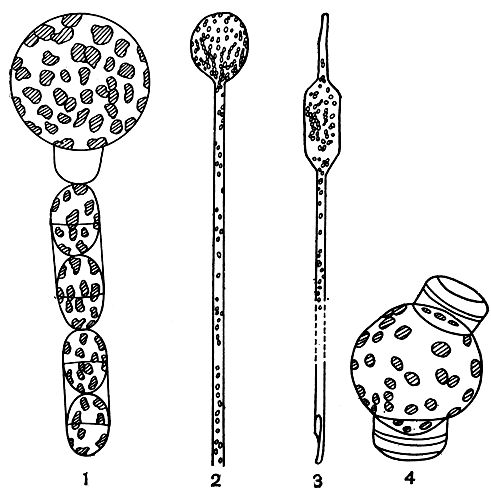
Рис. 92. Ауксоспоры: 1 - Melosira moniliformis, цепочка с конечной ауксоспорой; 2,3 - Rhizosolenia alata (2 - начало образования ауксоспоры, 3 - зрелая конечная ауксоспора); 4 - Thalassiosira excentrica, интеркалярная ауксоспора
После созревания ауксоспоры в ней развивается новая клетка, у которой сначала образуется эпитека, а затем гипотека. Первую клетку, возникшую из ауксоспоры, называют инициальной. По размерам она значительно превышает исходную.
Покоящиеся споры. Образованию покоящихся спор обычно предшествует или обильная вегетация вида, или наступление неблагоприятных условий. Протопласт клетки сжимается, округляется, и на его поверхности появляется сначала первичная створка споры, а затем вторичная, обе плотно соединяются краями (поясок у них отсутствует). Створки часто отличаются структурными элементами, они покрыты шипиками, выростами и некоторыми другими образованиями (рис. 93, 94). Обычно у диатомовых водорослей в клетке развивается только одна спора. Спустя определенное время покоящаяся спора, подобно ауксоспоре, увеличивается в объеме и дает начало новой клетке, вдвое большей по сравнению с исходной.
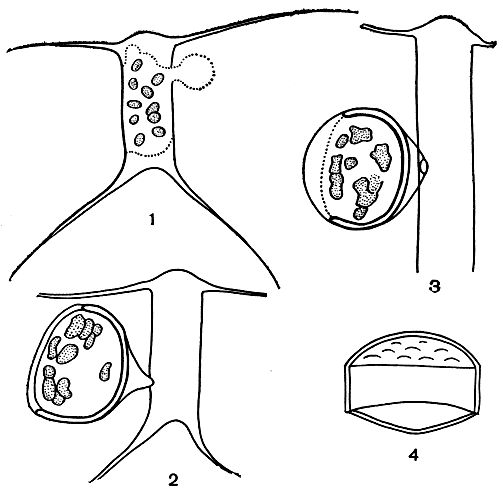
Рис. 93. Образование покоящихся спор: 1-4 - последовательные стадии развития споры у Chaetoceros heterovalvatus
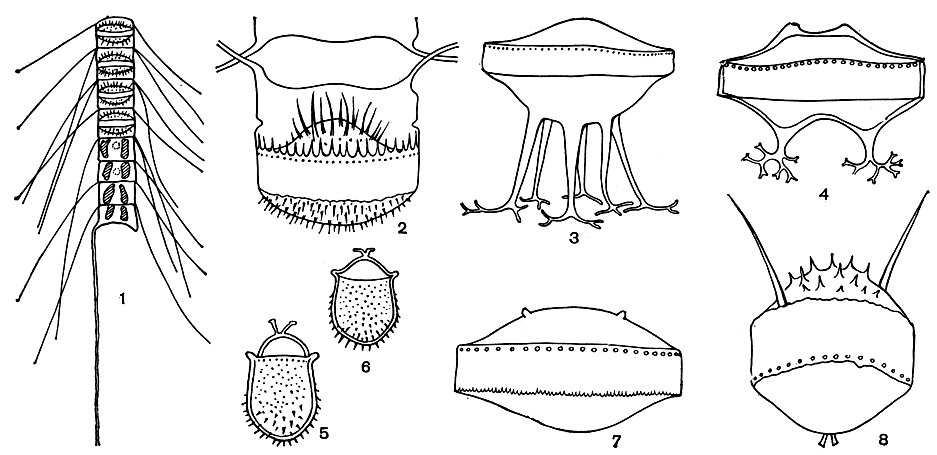
Рис. 94. Различные споры у видов рода Chaetoceros: 1 - Chaetoceros abnormis, цепочка с молодыми спорами; 2 - Ch. seiracanthus; 3 - Ch. subsecundus; 4 - Ch. lorenzianus; 5-6 - Ch. paulsenii; 7 - Ch. compressus; 8 - Ch. rigidus
Покоящиеся споры обычно образуют многие морские неритовые диатомеи, а также некоторые пресноводные виды. У представителей многих родов они возникают периодически как обычное явление в жизненном цикле.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'