Мохообразные
Систематика, строение и размножение мохообразных
Мхи в широком смысле этого слова, или мохообразные (Вrуоphyta), представляют собой довольно крупный, насчитывающий около 20 тысяч видов, отдел растительного царства.
Мохообразные - представители высших, или побеговых, растений (Cormophyla). В отличие от низших растений - водорослей и лишайников - тело большинства мохообразных представлено побегом, состоящим из стебля и листьев; только у части мохообразных тело представлено слоевищем (талломом). От низших растений мохообразные отличаются также многочисленными микроскопическими особенностями, в том числе наличием своеобразно устроенных гаметангиев (половых органов); мужских - антеридиев и женских - архегониев. Другой отличительный признак мохообразных - правильное чередование в нормальном цикле развития растения двух различных по своей морфологии поколений. Одно из поколений называется гаметофитом (растение, производящее половые элементы - гаметы), другое - спорофитом (растение, производящее элементы бесполого размножения - споры).
Образующийся на слоевищном или листостебельном гаметофите антеридий имеет вид многоклеточного мешочка, внутри которого образуются мужские гаметы - сперматозоиды. Архегоний имеет вид многоклеточной колбочки, в расширенной части которой - брюшке архегония - образуется женская гамета, или яйцеклетка. Если антеридии и архегоний располагаются на одном гаметофите, то такие растения называются однодомными. Если на одном растении (мужском) располагаются антеридии, а на другом (женском) - архегоний, то такие виды называются двудомными. Есть и многодомные мохообразные, у которых антеридии и архегоний могут располагаться на одном и на разных растениях того же вида.
При наличии капельножидкой воды сперматозоид достигает яйцеклетки и оплодотворяет ее. Из возникающей в результате оплодотворения зиготы вырастает спорофит, который у мохообразных называется спорогонием и который может состоять из стопы, ножки и коробочки. Спорогоний первоначально развивается в брюшке архегония. которое, разрастаясь, превращается в колпачок. С помощью стопы спорогоний высасывает из гаметофита воду с минеральными солями и органическими веществами. В коробочке спорогония образуется споровый мешок, или спорангий. Созревшая коробочка вскрывается, и споры попадают во внешнюю среду. При наличии благоприятных условий споры прорастают и дают начало новому гаметофиту. При этом первоначально образуется предросток, или протонема, имеющая вид многоклеточной нити, пластиночки, шаровидного тела и др., а затем уже вырастает гаметофор - собственно слоевищный или листостебельный гаметофит, несущий гаметангии, в которых вновь возникают сперматозоиды и яйцеклетки, и т. д. Таким образом происходит чередование поколений в жизненном цикле мохообразных.
Отличаясь рядом особенностей от низших растений, мохообразные стоят особняком и среди высших растений. В то время как у мохообразных спорофит растет и развивается, оставаясь все время прикрепленным к гаметофиту и паразитируя на нем. у других групп высших растений - плаунообразных, хвощеобразных, папоротникообразных и семенных - спорофит, напротив, большую часть своей жизни существует независимо от гаметофита и превышает его по своим размерам и степени морфологической дифференцировки. Это преобладание в цикле развития спорофита или гаметофита находит свое отражение в том, что у мохообразных растением мы обычно называем слоевищный или листостебельный гаметофит, а у остальных высших растений - листостебельный спорофит.
От большинства остальных высших растений мохообразные отличаются также отсутствием корней и некоторыми микроскопическими особенностями.
Мохообразные можно разбить на три класса: антоцеротовые (Anthocerotae), печеночники (Hepaticae) и мхи (Musci). Все три класса возникли на Земле очень давно, около 300 миллионов лет назад, и с тех пор развивались независимо один от другого, а потому наряду с общими признаками, указывающими на происхождение их от общего предка, эти классы обладают и рядом специфических, присущих только им особенностей.
Класс Антоцеротовые - Anthocerotae
Самый малочисленный класс мохообразных, насчитывающий около 300 видов, объединяемых в 3-5 родов. Гаметофит антоцеротовых представляет собой распростертое по поверхности субстрата (почвы, реже коры деревьев) вильчато ветвящееся слоевище, имеющее обычно вид многолопастной темно-зеленой (табл. 41), иногда курчавой по краям розетки. Листьев у растения нет, но иногда верхняя поверхность слоевища покрыта пластинчатыми или волосковидными придатками, защищающими его от высыхания. На нижней поверхности слоевища имеются одноклеточные волоски, или ризоиды, с помощью которых растение прикрепляется к почве и всасывает из нее воду с растворенными в ней минеральными солями. Слоевище сложено из однородных клеток, в которых протекает фотосинтез, и нарастает с помощью особых меристематических клеток, находящихся в углублениях между лопастями. На нижней (а иногда и на верхней) поверхности слоевища некоторых антоцеротовых имеются устьица, которые ведут в полости, заполненные слизью и колониями сине-зеленых водорослей; полости заметны иногда в виде темных точек. У некоторых видов отдельные лопасти слоевища углубляются в почву и образуют клубеньки - органы вегетативного размножения, с помощью которых растение способно переносить неблагоприятные (засушливые) сезоны года; растение может также размножаться и с помощью очень мелких выводковых почек.
На верхней стороне слоевища в особых, хорошо заметных с помощью лупы камерах образуются антеридии и архегонии. Во время дождя или обильной росы созревшие в антеридиях сперматозоиды выплывают из антеридиальных камер и один из них оплодотворяет находящуюся в брюшке архегония яйцеклетку, в результате чего возникает зигота, из которой вырастает спорогоний.
Спорогоний антоцеротовых состоит из стопы и коробочки; ножки обычно нет, или она очень короткая. С помощью погруженной в ткань слоевища стопы спорогоний всасывает из него воду с питательными веществами. В основании коробочки у большинства видов сохраняется меристематическая ткань, с помощью которой спорогоний может долго нарастать в длину, достигая иногда 15 см. Коробочка в молодом состоянии зеленая, в стенках ее имеются устьица, а в клетках идет фотосинтез. В коробочке образуются споры и особые стерильные клетки. У большинства антоцеротовых в центре коробочки формируется колонка и по созревании спорогония стенка коробочки растрескивается сверху вниз на две створки, между которыми в виде щетинки заметна колонка (рис. 17,1). Попадая на влажный субстрат, споры прорастают, и при благоприятных условиях из них возникает новое слоевище.
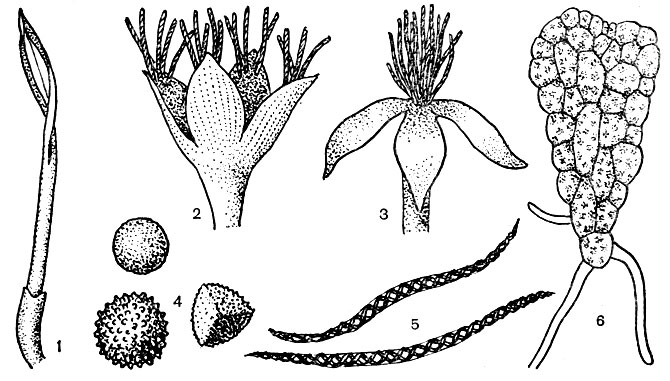
Рис. 17.1. (1 - коробочка антоцеротовых, вскрывающаяся двумя створками, между которыми сохраняется колонка, 2 - коробочка юнгерманниевых с кисточками элатер на концах створок, 3 - коробочка юнгерманниевых с кисточкой элатер на две коробочки, 4 - споры печеночников, 5 - элатеры печеночников, 6 - молодая пластинчатая протонема печеночников)
Класс Печеночники - Hepaticae
Второй по величине класс мохообразных, насчитывающий около 6 тысяч видов и около 300 родов, объединяемых примерно в полсотни семейств. Печеночники делят на два подкласса - маршанциевых (Marchantiidae) и юнгерманниевых (Jungermanniidae), каждому из которых присущи наряду с общими для обоих подклассов признаками и свои специфические признаки.
Все представители подкласса маршанциевых - слоевищные растения. Слоевища маршанциевых (рис. 13) стелются по поверхности субстрата, нарастают в результате деления клеток, расположенных в выемках на концах слоевищ, и вильчато ветвятся, образуя более или менее рыхлые коврики либо округлые розетки. Длина отдельных ветвей слоевища у разных видов измеряется несколькими миллиметрами или сантиметрами, а толщина их не превышает 1-3 мм. У слоевища иногда можно выделить срединное ребро, представляющее собой более темный и толстый участок слоевища по сравнению с более светлыми и тонкими крыльями. На нижней, или брюшной, поверхности слоевища обычно находятся многочисленные ризоиды, посредством которых слоевище прикрепляется к субстрату и снабжается водой и растворенными в ней минеральными солями. На брюшной же поверхности слоевища обычно располагаются в один или несколько рядов по обе стороны срединного ребра прозрачные красноватые или зеленоватые небольшие брюшные чешуйки, или амфигастрии, разнообразной, нередко более или менее серповидной либо полукруглой формы (рис. 14). Чешуйки защищают точки роста слоевища от высыхания и прижимают тяжи ризоидов к брюшной поверхности слоевища.
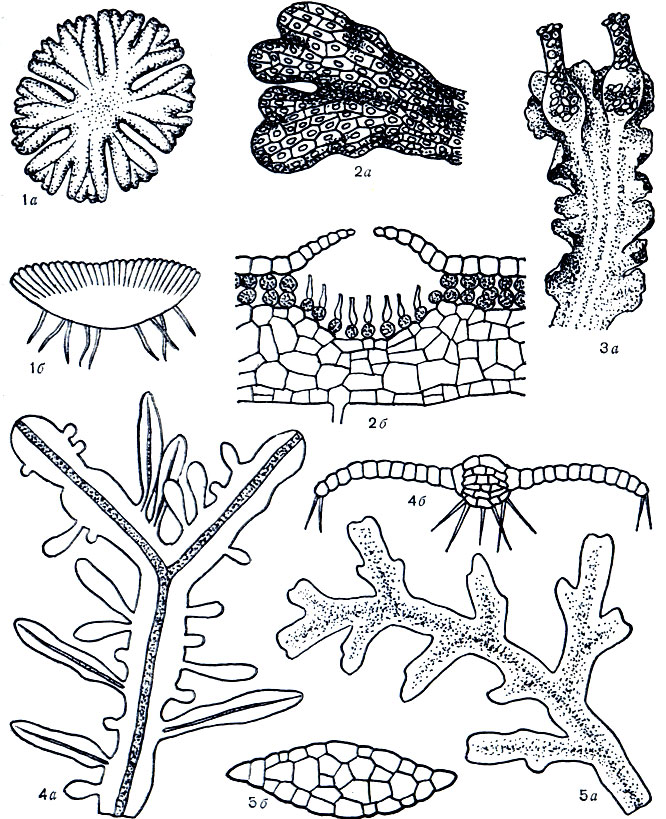
Рис. 13. Слоевища представителей подклассов маршанциевых (1 - 2) и юнгерманниевых (3 - 5): 1 - розетковидное многослойное слоевище с ассимиляторами на верхней и ризоидами на нижней поверхности слоевища, 2 - многослойное слоевище с воздушными камерами и ассимиляторами, 3 - многослойное слоевище с листовидными лопастями и выводковыми колбочками, 4 - слоевище со срединным ребром, однослойнными крыльями и с ризоидами на нижней поверхности, 5 - многослойное слоевище без ребра (а - вид сверху, б - разрез)

Рис. 14. Амфигастрии (1-6) и их расположение (7,8): 1 - серповидный , 2 - округло- квадратный многолопастной, 3 - ланцетный, 4 - округлый цельнокрайний, 5 - овальный двулопастной. 6 - двураздельный с ланцетными лопастями, 7 - расположение амфигастриев у юнгерманниевых, 8 - расположение амфмгастриев у маршанциевых (а - амфигастрий)
Слоевище маршанциевых многослойное и обычно состоит из разнородных тканей: прежде всего из основной, или запасающей, ткани и ассимиляционной ткани, которая располагается на верхней стороне слоевища. Ассимиляционная ткань у некоторых маршанциевых представлена ассимиляторами - клетками с хлоропластами, в которых идет фотосинтез. Ассимиляторы часто находятся в воздушных камерах, а сами воздушные камеры (с ассимиляторами или без них) нередко видны сверху в виде многоугольников, в центре которых с помощью лупы можно заметить отверстие - устьице, ведущее в камеру (рис. 13, 2).
Отмирание старых или поврежденных участков слоевища приводит к распадению его на отдельные части, каждая из которых обособляется в качестве нового растения. Кроме этого простейшего способа вегетативного размножения некоторые маршанциевые обладают способностью развивать выводковые побеги, клубеньки или тела, причем выводковые тела располагаются в особых вместилищах, или выводковых корзиночках (табл. 41).
Половые органы у маршанциевых возникают поодиночке или группами в полостях на спинной поверхности слоевища или на особых видоизмененных ветвях слоевища - мужских и женских подставках, сидящих непосредственно на слоевище или обладающих ножками, которые у женских подставок сильно вытягиваются к моменту созревания спорогониев (рис. 15, 4, 5).
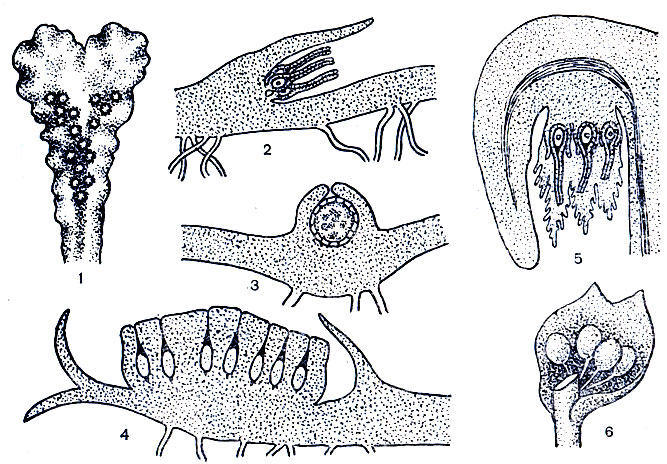
Рис. 15. Расположение гаметангиев у печеночников: 1 - антеридиальные камеры на поверхности слоевища у пеллии, 2 - собрание архегониев у пеллии, 3 - антередиальная камера у пеллии, 4 - мужская подставка у коноцефалка, 5 - женская подставка у маршанции, 6 - антерадии в пазухе листа у мареупеллы (2-5 - разрезрезы слоевищ и подставок)
Группы архегониев окружены нередко общей оберткой, или покрывалом (перихецием); при основании каждого архегония может возникать и частная обертка - перианций (или псевдоперианций), достигающий окончательных размеров лишь при созревании спорогония (рис. 16, 6).

Рис. 16. Спорогонии и защитные образования вокруг них у печеночников: 1 - шаровидный спорогоний риччии, погруженный в ткань слоевища (стенка коробочки растворилась при созревании спор), 2 - спорогоний блазии с оберткой при основании , 3 - спорогоний риккардии мясистым цилиндрическим колпачком в основании ножки, 4 - спорогоний пеллии с кармашковидной оберткой и колпачком у основания ножки, 5 - спорогоний пеллии с грушевидной оберткой и колпачком у основания ножки, 6 - женская подставка маршанции в разрезе (а - перихеций, б - переанций), 7 - спорогоний маршаций (а - коробочка, б - ножка, в - колпачок, г - перианций), 8 - переанций ребристый, суженный в носик, 9 - перианций складчатый, зубчатый по краю, 10 - перианций не складчатый. зубчатый по краю, 11 - перианций сплющенный, с широким двуполостным устьем, 12 - побег со стеблевым перигинием, 13 - цеокаул трихоколеи (слева - с поверхности, справа - в разрезе)
Возникающий из зиготы спорогоний маршанциевых может состоять из стопы, ножки и коробочки. Молодой спорогоний находится внутри разросшегося брюшка архегония, или колпачка, который разрывается при удлинении ножки (рис. 16,7). Посредством стопы спорогоний получает из слоевища воду и необходимые питательные вещества. В коробочке образуются споры, а у большинства родов - особые клетки со спиральными утолщениями - элатеры (рис. 17), играющие большую роль в распространении спор. При созревании спорогония ножка его несколько удлиняется и выносит коробочку за пределы колпачка и обертки (или оберток); у некоторых маршанциевых ножка недоразвита (рис. 16, 1).
Созревшая коробочка вскрывается путем отделения верхней крышечкоподобной части и разрывания оставшейся урночкоподобной части на неправильные лопасти; стенка коробочки может также распадаться на куски или разрываться на лопасти (рис. 16, 7). У некоторых маршанциевых (например, у риччии - Riccia) стенка коробочки после созревания спор растворяется (рис. 16,7).
Среди представителей подкласса юнгерманниевых встречаются как слоевищные, так и листостебельные растения. Слоевище юнгерманниевых (рис. 13) по внешнему виду часто похоже на слоевище маршанциевых, но отличается от последнего простым внутренним строением (не имеет специализированной ассимиляционной ткани). Слоевище юнгерманниевых может обладать срединным ребром, или ребро у слоевища не выражено. На брюшной поверхности слоевища развиваются ризоиды, а по краю иногда располагаются листовидные лопасти-придатки. Слоевища могут образовывать специализированные органы вегетативного размножения - особые укороченные ветви и выводковые тела, иногда сидящие в своеобразных вместилищах. Половые органы у слоевищных юнгерманниевых сидят на спинной поверхности слоевища, располагаясь иногда в особых камерах, но никаких подставок у юнгерманниевых нет (рис. 15, 1-3). Вокруг группы архегониев может возникать общая обертка.
Листостебельные юнгерманниевые обладают стелющимися по поверхности субстрата или приподнимающимися (редко прямостоячими) стеблями, от которых у большинства видов отходят два ряда боковых однослойных листьев без жилки. Форма боковых листьев очень разнообразна (рис.18). Наряду с плоскими листьями у юнгерманниевых встречаются и сложенные листья, у которых верхняя и нижняя лопасти, доли или сегменты иногда резко различаются между собой по форме и выполняют даже различные функции. Например, у фруллании (Frullania) нижний сегмент листа служит для запасания воды. От нижней стороны стебля отходят ризоиды. На ней же часто развиваются амфигастрии; они представляют собой обычно сильно видоизмененные листья третьего ряда, и форма их довольно разнообразна (рис. 14).
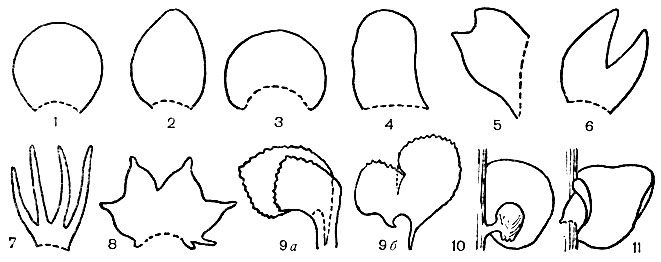
Рис. 18. Форма и расчленение листьев представителей подкласса юнгерманниевых: 1 -
При определении родов листостебельных юнгерманниевых большое значение придается способу прикрепления листьев к стеблю. У двух распространенных в СССР видов листья прикрепляются поперечно, у остальных - косо. Если передний край листа находится над задним краем соседнего листа, расположенного на той же стороне стебля, но ближе к верхушке побега, то листья называют набегающими (покрывающими); если же передний край листа находится под задним краем соседнего листа, расположенного ближе к верхушке побега, то листья называют сбегающими (прикрытыми). В зависимости от того, под каким углом каждый лист прикрепляется к стеблю, различают листья почти
вдоль прикрепленные, косо прикрепленные и почти поперечно прикрепленные (рис. 19).
Листостебельные юнгерманниевые, как и слоевищные, могут образовывать специализированные органы вегетативного размножения - выводковые почки, которые сидят по краям листьев на обычных или видоизмененных побегах или непосредственно на верхушках стеблей и заметны иногда невооруженным глазом в виде налета из мельчайших крупинок.
Антеридии у листостебельных юнгерманниевых возникают обычно в пазухах листьев (рис. 15, 6), нередко видоизмененных; у двудомных и однодомных раздельнополых видов веточки с антеридиями называются антеридиальными колосками (рис. 29). Архегонии у листостебельных юнгерманниевых образуются на концах побегов под защитой покровных листьев. Часто два или три самых верхних листа, срастаясь друг с другом, образуют покров, или перианций (рис. 16, 8-11), соответствующий общей обертке маршанциевых или слоевищных юнгерманниевых в том смысле, что он окружает несколько архегониев.
Развивающийся из зиготы спорогоний юнгерманниевых, как и спорогоний маршанциевых, состоит из стопы, ножки и коробочки. В коробочке образуются споры и элатеры. Спорогоний вплоть до момента созревания спор защищен от неблагоприятных воздействий внешней среды колпачком и различными структурными образованиями гаметофита. У слоевищных юнгерманниевых спорогоний защищен сильно разрастающимся колпачком или общей оберткой (рис. 16, 2-5), хорошо заметными при основании ножки зрелого спорогония. У листостебельных юнгерманниевых спорогоний чаще всего находится под защитой разросшегося перианция разнообразной формы (рис. 16, 8-11). Иногда спорогоний находится внутри стеблевого перигиния, представляющего собой трубку, к верхнему краю которой прикреплены покровные листья и редуцированный перианций (рис. 16, 12). У некоторых листостебельных юнгерманниевых верхушка ветви с перигинием углубляется в почву и образует плодовый мешок, или марсупий, на дне которого сидит молодой спорогоний. Изредка колпачок и перианций не развиваются, и спорогоний находится внутри облиственной верхушки стебля - целокаула (рис. 16, 13).
Ножка спорогония к моменту созревания спор быстро удлиняется; очень длинной бесцветной ножкой спорогоний юнгерманниевых хорошо отличается от спорогония маршанциевых. Коробочка у юнгерманниевых чаще всего вскрывается четырьмя створками; на их концах или между ними, на дне коробочки, иногда имеются кисточки из прикрепленных элатер, играющие большую роль в рассеивании спор (рис. 17).
Протонема печеночников обычно имеет вид пластиночки либо шаровидного или эллипсоидального тела (рис. 17,6). Из протонемы затем вырастает слоевище или листостебельный побег.
Класс мхи - Musci
Самый крупный класс мохообразных, насчитывающий около 800 родов и до 15 тысяч видов, относимых примерно к 80 семействам. Все мхи - листостебельные растения, но в отличие от листостебельных юнгерманниевых мхи по большей части обладают хорошо выраженной нитчатой протонемой, все листья у мхов нормально развиты (нет амфигастриев) и всегда цельные, а коробочка спорогония обычно имеет колонку и никогда не содержит элатер.
Класс мхов разделяют на три подкласса, хорошо различающихся между собой по строению спорогония и некоторым особенностям гаметофита: сфагновые (Sphagnidae), андреэевые (Andreaeidae) и бриевые (Bryidae); первые два класса невелики по объему и содержат соответственно около 300 и около 100 видов.
Все представители подкласса сфагновых мхов - влаголюбивые растения, и побег сфагнов обладает многочисленными особенностями, позволяющими растению удерживать и сохранять воду. Стебель сфагнов прямостоячий, редкооблиственный, с многочисленными густооблиственными боковыми ветвями (табл. 44). Ветви на верхушке стебля скучены в плотную головку, ниже - собраны в пучки; при этом часть ветвей каждого пучка оттопырена в сторону, благодаря чему побеги соедршяются в более или менее плотные дерновинки или подушечки. Другая часть ветвей каждого пучка свисает вдоль стебля, образуя своеобразный фитиль, по которому передвигается вода. Ризоидов у взрослого растения нет. Побег нарастает в верхней части и постепенно отмирает снизу. Некоторые ветви на верхушке побега растут быстрее остальных и при отмирании стебля снизу дают начало новому побегу, вследствие чего подушечки сфагна постепенно увеличиваются в объеме.

Таблица 44. 1 - сфагн дубравный, 2 -nсфагн Вульфа, 3 - сфагн компактный, 4 - сфагн Гиргензона, 5 - сфагн магелланский, 6 - сфагн оттопыренный (а - растение, б - отстоящая веточка)
Снаружи стебель главного побега и ветвей (рис. 20) покрыт бесцветной кожицей - гиалодермисом, состоящим из крупных мертвых клеток, в стенках которых у многих видов имеются отверстия - поры. В гиалодермисе ветвей часто встречаются водосборные ретортовидные клетки с отверстиями на концах. Листья сфагнов однослойные, без жилки, разнообразной формы; листья главного побега обычно отличаются по форме, микроструктуре и размерам от веточных листьев того же вида (рис. 30). Пластинка листа (рис. 20) состоит из клеток двух типов - узких живых фотосинтезирующих клеток, оболочка которых нередко пропитана красящими веществами, и широких мертвых водоносных клеток. Прозрачные, или гиалиновые, мертвые клетки, как и клетки гиалодермиса стебля, имеют отверстия - поры. Микроскопические особенности строения листьев играют важную роль при определении видов сфагновых мхов. Через поры гиалиновых клеток листьев и стебля, через отверстия ретортовидных клеток, а также по капиллярным промежуткам между свисающими ветвями и стеблем вода легко передается от одной части растения к другой, и это объясняет замечательную способность сфагнов быстро и в большом количестве впитывать воду. При испарении воды гиалиновые клетки заполняются воздухом, поэтому сухие растения сфагна приобретают беловатый оттенок.
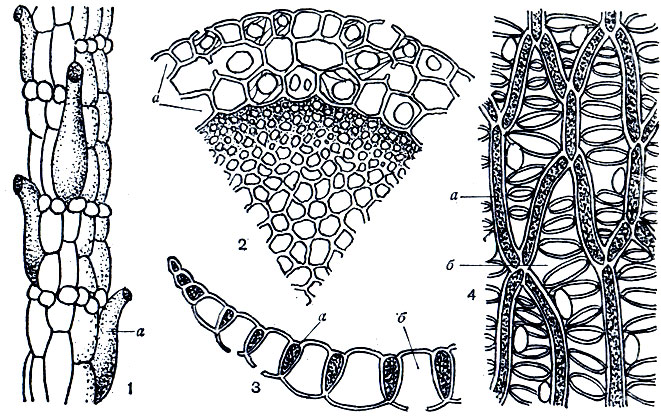
Рис. 20. Строение стебля и листа сфагна: 1 - поверхность стебля с ретортовидными клетками (а), 2 - поперечный срез стебля (а - гиалодермис), 3 - поперечный срез листа, 4 - лист с поверхности (а - фотосинтезирующие клетки, б - гиалиновые клетки)
Антеридии и архегонии у сфагнов располагаются на особых ветвях (рис. 21,4) одного и того же растения или на разных растениях. Антеридии сидят в пазухах покровных листьев. Ветви с антеридиями обычно булавовидно вздуты и ярко окрашены в различные цвета, но после созревания антеридиев ветви удлиняются и не отличаются по окраске от остальных отстоящих ветвей растения. Архегонии располагаются на концах укороченных почковидных женских веточек также под защитой покровных листьев.

Рис. 21. Строение сфагновых мхов: 1 - спора, 2 - молодая протонема, 3 - пластинчатая зрелая протонема с молодым листостебельным побегом (гаметофором), 4 - часть стебля с антеридиальными (а) и с архегониальными (б) ветвями, 5 - антеридиальная ветвь, частично лишенная листьев (а - антеридий), в - вскрывающийся антеридий, 7 - сперматозоид, 8 - верхушка архегониальной ветви на продольном срезе, 9 - продольный срез спорогония (а - ложноножка, б - стопа, в - колонка, г - спорангий, д - колпачок), 10 - зрелый спорогоний на ложноножке (а - ложноножка, б - остатки колпачка, в - коробочка, г - крышечка)
Молодой спорогоний (рис. 21, 9), находящийся внутри колпачка, прикрыт снаружи покровными листьями и состоит из стопы и коробочки; ножка у спорогония сфагновых мхов недоразвита. Со дна коробочки поднимается куполообразная колонка, прикрытая сводообразным спорангием. При созревании коробочка прорывает колпачок, который остается в виде оборки при ее основании, и в это же время часть стебля, расположенная между стопой и покровными листьями, сильно удлиняется, образуя ложноножку, которая выносит шаровидную коробочку далеко за пределы покровных листьев (рис. 21, 10). Коробочка обычно вскрывается путем резкого отбрасывания крышечки, при этом споры выбрасываются на расстояние до 10 см. Процесс вскрывания коробочки и выбрасывания спор легко проследить, подсушивая выбранные из влажной дернинки растения со зрелыми спорогониями.
Протонема имеет сначала вид короткой нити, а затем лопастной пластинки (рис. 21, 2, 3), на которой закладывается почка, дающая впоследствии листостебельный побег, или гаметофор.
Представители подкласса андреэевых мхов отличаются от остальных мхов целым рядом признаков. Спора андреэевых, прорастая, дает начало стелющейся по поверхности субстрата (обычно камней и скал) разветвленной пластинчатой протонеме с листообразными придатками и древовидными выростами (рис. 22). На протонеме появляются почки, из которых вырастает гаметофор, прикрепляющийся к субстрату толстыми цилиндрическими или пластинчатыми ризоидами. Спорогоний андреэевых сидит на верхушке стебля; он обладает колонкой, не доходящей до верхушки коробочки, и не имеет развитой ножки. Созревшую коробочку за пределы покровных листьев выносит удлиняющаяся ложноножка. Коробочка вскрывается 4-8 продольными щелями (рис. 27, 1).
Подкласс бриевых, или зеленых, мхов - наиболее многочисленная группа мохообразных. Протонема этих мхов обычно имеет вид сильноразветвленной, стелющейся по поверхности субстрата нити, от которой в почву отходят незеленые ризоиды (рис. 22, 2-3). У некоторых бриевых протонема листовидная, например у тетрафиса (Tetraphis) (рис. 22, 5). У схистостеги (Schistostega) - мха, растущего в расщелинах скал, в пещерах, дуплах деревьев, одни ветви протонемы, прикрепляющие ее к субстрату, нитчатые, другие, располагающиеся над субстратом, имеют вид однослойных пластинок, направленных перпендикулярно к лучам света (рис. 22, 4). Пластинки эти состоят из линзовидно-воронковидных клеток, в воронковидной части которых находятся зеленые пластиды (хлоропласты), а над хлоропластами, подобно хрусталику в глазном яблоке, располагается бесцветная масса цитоплазмы. Рассеянные лучи света, падающие на поверхность пластинки, преломляются в линзовидной части клетки и фокусируются на хлоропластах; при этом часть лучей отражается, что вызывает свечение пластинки зеленым светом (отсюда народное название схистостеги - "светящийся мох").
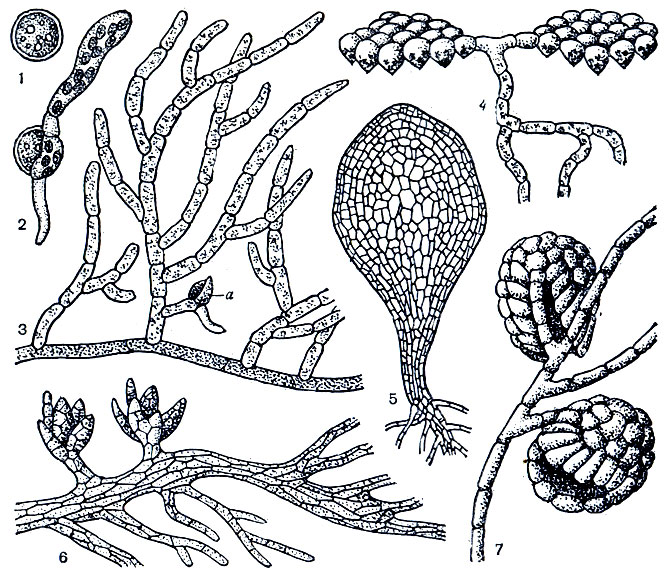
Рис. 22. Спора и протонема андрезевых и бриевых мхов: 1 - спора у фанарии, 2 - прорастающая спора у фанарии, 3 - нитчатая протонема с почечкой (а) у фанарии, 4 - протонема схистостеги, 5 - пластинчатая протонема тетрафиса, 6 - протонема андрези, 7 - протонема и мужской гаметофор у буксбаумии
Протонема некоторых бриевых мхов (например, фаска - Phascum, поттии - Pottia) в противоположность листостебельному побегу этих растений многолетняя, зимующая. У буксбаумии (Buxbaumia) протонема также многолетняя, а мужской гаметофор у буксбаумии сведен до одного листа, под защитой которого находится антеридий (рис. 22, 7).
Из почек, образующихся на протонеме, вырастают листостебельные побеги, длина которых варьирует от долей миллиметра до полуметра. Побеги бывают простыми или разветвленными, прямостоячими, приподнимающимися или восходящими, висячими, стелющимися или лежачими (ползучими). Способ ветвления мхов часто связан с расположением половых органов, а точнее, спорогониев. У мхов, у которых спорогоний располагается на верхушке главного побега или столь же мощных ветвей (верхоплодные мхи), побеги простые либо вильчато или мутовчато ветвящиеся. У мхов, у которых спорогоний располагается на верхушке сильно укороченных почковидных боковых ветвей (бокоплодные мхи), побеги ветвятся обычно по-иному (главным образом перисто) и обладают хорошо выраженным главным стеблем. У некоторых бриевых мхов (например, климация) имеются мощные подземные корневище-подобные побеги (табл. 51), которые служат для вегетативного размножения и, выйдя на поверхность, превращаются в обычные побеги.
Стебель у бриевых мхов тонкий, 0,1-1 мм в диам., почти бесцветный либо окрашенный - зеленоватый, красноватый, бурый или черноватый, в нижней части или почти на всем своем протяжении покрытый ризоидами - однорядными многоклеточными, обычно бурыми или красноватыми, простыми или разветвленными волосками (рис. 23, 7). Ризоиды, соприкасающиеся с субстратом, внедряются в него, прикрепляют к субстрату растение и снабжают его водой с растворенными в ней минеральными солями; на остальной части стебля ризоиды нередко образуют густой войлок, который может объединять отдельные побеги в более или менее плотную дерновинку.

Рис. 23. Строение бриевых мхов: 1 - стебель на поперечном срезе (на поверхности стебля видны ризоиды, в центре - проводящий пучок), 2 - парафилл, 3 - корзиночка с выводковыми телами у тетрафиса, 4 - ложноножка с выводковыми телами у аулакомний, 5 - чашевидная (а) и проросшая (б) мужская почка погоната, в - обоеполая почка брия на продольном разрезе (а - анеридий, б - архегоний, в - парафиза), 7 - мужские растения сплахна, 8 - спорогоний сплахна с зонтиковидной апофизой
Листья бриевых мхов не имеют черешка и прикрепляются к стеблю более или менее широким основанием всегда цельной пластинки поперек, наискось или почти вдоль стебля, располагаясь в два или несколько вертикальных рядов. Побеги могут быть густо и рыхлооблиственными. Иногда у верхушки стебля листья более тесно скучены, чем на остальном его протяжении, и образуют розетку или хохолок. Во влажном состоянии листья плотно прижаты к стеблю, черепитчато налегая один на другой, или отстоят от стебля под тем или иным углом. В зависимости от величины угла различают листья прямостоячие (угол до 15°), прямо-отстоящие (угол до 30°), отклоненные (угол около 45°), оттопыренные (угол 45-90°) и отогнутые (угол более 90°); иногда основание листа может быть прямоотстоящим, а верхушка - отогнутой. Обычно листья равномерно обращены во все стороны, реже - одностороннеобращенные. У сухих растений по сравнению с влажными характер отхождения листьев от стебля может меняться, а пластинка листа иногда завивается вокруг стебля либо изгибается или закручивается.
Пластинка обычного фотосинтезирующего листа бывает плоской, ложковидно- или чашевидно-вогнутой, килеватой, желобчатой, трубчатой, продольно-складчатой (далее в тексте просто складчатой) или поперечно-волнистой (далее в тексте просто волнистой). По очертаниям различают листья треугольные, сердцевидные, округлые, яйцевидные, эллиптические, продолговатые, языковидные, лопатчатые, ланцетные, линейные, серповидные, крючковидные, треугольно-сердцевидные, продолговато-ланцетные и т. д. Пластинка листа (за исключением жилки, а иногда и каймы) всегда однослойная и состоит из клеток, которые могут различаться между собой по форме, величине и строению. Основание листа бывает суженным или расширенным, закругленным или выемчатым; иногда основание низбегает по стеблю или, разрастаясь, образует более или менее крупное, охватывающее стебель влагалище. Клетки, расположенные в углах основания пластинки, могут отличаться по форме, величине и цвету от остальных клеток пластинки, иногда образуя служащие для сохранения воды участки - листовые крылья; если крылья сильно вздуты, они называются листовыми ушками (в описаниях те и другие обозначены как ушки). Края листовой пластинки бывают цельными или зубчатыми; они также могут быть плоскими, либо загнутыми (завороченными) на верхнюю сторону, либо отогнутыми (отвороченными) на нижнюю сторону. Клетки, расположенные по краю и отличающиеся от остальных клеток пластинки формой, величиной и окраской, образуют плоскую или вздутую кайму. Верхушка листовой пластинки может быть выемчатой, закругленной, тупой, длинно-или короткозаостренной, шиловидной, прямой или крючковато-изогнутой, колпачковидной, с остроконечием.
У большинства видов бриевых мхов листья обладают более или менее мощной жилкой, которая представляет собой тяж, сложенный несколькими слоями клеток. Жилка бывает узкой или широкой, на верхней стороне жилки иногда располагаются продольные зеленые пластинчатые выросты - ассимиляционные пластиночки, или ассимиляторы. Жилка чаще простая (но может также быть вильчатой или двойной), короткая либо длинная, заканчивающаяся в нижней или верхней половине листа. Она может выступать из верхушки листа в виде острия либо волоска.
Микроскопические особенности строения пластинки обычно играют решающую роль в определении видов бриевых мхов.
Фотосинтезирующие листья на протяжении одного и того же стебля или листья главного побега и боковых ветвей, а также спинные, боковые и брюшные листья у плоскооблиственных побегов могут различаться между собой по форме и величине. Листья на подземных или стелющихся по поверхности субстрата корневище-подобных частях побега по форме, величине и строению обычно отличаются от нормальных фотосинтезирующих листьев и часто имеют вид чешуи. Быстрорастущие побеги с расставленными чешуевидными листьями называются столоновидными побегами. Нередко от обычных вегетативных листьев отличаются по форме и величине, а иногда и по окраске листья, окружающие и защищающие половые органы, расположенные на верхушках побегов и ветвей (рис. 23, 5). Листья, окружающие антеридии, называются перигониальными, а окружающие архегонии - перихециальными.
Стебли некоторых бриевых мхов несут кроме листьев расположенные без всякого порядка зеленые листовидные или нитевидные придатки - парафиллы (рис. 23, 2), образующие иногда густой зеленый войлок на поверхности стебля и служащие для ассимиляции и для поглощения воды. Недоразвитые листья в основании ветвей, напоминающие по облику парафиллы, называются псевдопарафиллами.
Бриевые мхи обладают прекрасно выраженной способностью к вегетативному размножению. Они могут размножаться посредством разделения на части единой протонемы или путем образования вторичных предростков из ризоидов. Отломанные кусочки побега или листа, попав в благоприятные условия, также могут дать начало новым побегам или же вторичной протонеме, на которой возникают многочисленные почки. Образование новых экземпляров происходит и при обособлении ветвей вследствие отмирания нижних участков побега. Вегетативное размножение осуществляется также с помощью столоновидных или корневи-щеподобных побегов, вследствие чего постепенно увеличивается площадь дерновинки.
Есть у бриевых мхов и специализированные органы вегетативного размножения: легко отламывающиеся от материнского побега флагеллы (нитевидные хрупкие побеги с чешуйчатыми листьями); выводковые веточки, расположенные в пазухах листьев и по созревании легко отделяющиеся от материнского растения; выводковые клубеньки и ломкие листья; выводковые тела нитевидной, булавовидной или шарообразной формы, развивающиеся на обычных стеблях и листьях или в особых чашевидных вместилищах (рис. 23, 3) или же на конце удлиненного безлистного стебля-ложноножки (рис. 23, 4).
Некоторые виды мхов на большей части территории своего распространения размножаются исключительно вегетативным путем и не образуют спорогониев.
Антеридии и архегонии бриевых мхов вместе с нитевидными структурами - парафизами располагаются на верхушках главных побегов или боковых ветвей и находятся под защитой покровных листьев, образуя половые почки, или "цветки" (рис. 23, 5, 6). Почки имеют дисковидную, головчатую или эллипсоидальную форму; они могут быть обоеполыми, т. е. содержать антеррщии и архегонии, и однополыми - мужскими или женскими, т. е. содержать, соответственно, только антеридии или только архегонии. У двудомных мхов мужские экземпляры иногда резко отличаются по размерам и облику от женских, будучи меньше последних в несколько раз (рис. 23, 7,8; табл. 47).
Мужские почки у некоторых мхов после созревания антеридиев могут прорастать и давать начало олиственному побегу (рис. 23, 5). В таких случаях благодаря наличию мутовок перигониальных листьев, отличных по форме и цвету от обычных вегетативных листьев, легко можно установить величину годичного прироста и возраст растения.
Оплодотворение у наземных, наскальных и эпифитных видов происходит во время дождя или обильной росы, у водных и болотных мхов - по мере созревания половых органов. У некоторых двудомных мхов сперматозоиды, а точнее, слизь со сперматозоидами, по-видимому, может переноситься к женским почкам мелкими животными (насекомыми, червями и т. п.).
В результате оплодотворения у большинства бриевых мхов в половой почке развивается лишь один спорогоний, но у некоторых видов брия (Вrуum) и мния (Mnium) в почке могут появиться несколько спорогониев. Развитие спорогония продолжается от одного до нескольких месяцев, и созревание его, так же как и процесс оплодотворения, у большинства видов приурочено к определенному времени года.
Зрелый спорогоний большинства бриевых мхов состоит из стопы, погруженной в ткань олиственного побега, ножки и коробочки, которая сверху прикрыта колпачком. Ножка спорогония может быть очень короткой, и в этом случае коробочка бывает погружена в перихециальные листья (рис. 24, 1). Но чаще ножка бывает удлиненной, прямой либо более или менее согнутой (рис. 24, 2). При созревании и подсыхании спорогония ножка нередко спирально скручивается (рис. 25, 2). Поверхность ножки может быть гладкой или шероховатой из-за мелких бородавочек.
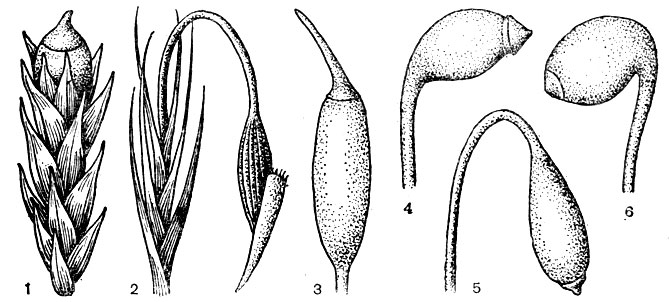
Рис. 24. Коробочка бриевых мхов: 1 - погруженная, прямая, с короткоклювовидной крышечкой, 2 - повислая. прямая, эллипсоидальная, бороздчатая, 3 - прямая, продолговато-эллипсоидальная. с длинно-клювовидной крышечкой. 4 - горизонтальная,слегка согнутая, продолговато-яйцевидная, с короткой шейкой и конусовидной крышечкой, 5 - повислая, обратнояйцевидная, с длинной шейкой и сводообразной крышечкой с сосочком, 6 - горизонтальная, согнутая, продолговато-яйцевидная. с короткой прямой шейкой и тупоконусовидной крышечкой
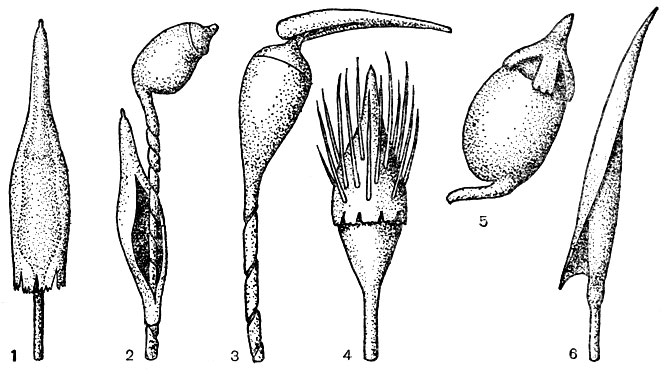
Рис. 25. Колпачок и коробочка бриевых мхов: 1 - колпачок узкоколокольчатый, бахромчатый по краю, голый, коробочка прямостоячая, пряма; 2 - колпачок остающийся на ножке в виде манжеты, голый, коробочка на скрученной ножке, наклоненная. яйцевидная. со слабосогнутой шейкой, с зобиком; 3 - колпачок клобуковидный, голый, коробочка прямостоячая, прямая, с шейкой; 4 - колпачок конусовидно-колокольчатый, волосистый коробочка прямостоячая,прямая; 5 - колпачок шапочковидный, голый, по краю лопастной, коробочка на короткой согнутой ножке, широкоэллипсоидальная; 6 - колпачок клобуковидный, голый, коробочка прямостоячая,прямая, продолговато-эллипсоидальная, с короткой шейкой
Коробочка (рис. 24, 25) бывает прямой либо более или менее согнутой, радиально-симметричной или двусторонне-симметричной, прямостоячей, наклоненной, горизонтальной, повислой. Форма коробочки варьирует от шаровидной до удлиненно-цилиндрической, а поверхность коробочки бывает ровной, ребристой или бороздчатой.
В коробочке бриевых мхов можно выделить шейку, апофизу (гипофизу), урночку и крышечку (рис. 26). Шейка - это нижняя часть коробочки, более или менее постепенно переходящая в ножку. Шейка (рис. 24, 25) бывает очень короткой, практически незаметной либо более или менее длинной, прямой или согнутой, иногда со вздутием с одной стороны (зобиком). Апофиза - это вздутая нижняя часть коробочки, резко отграниченная от урночки, иногда яркоокрашенная и превышающая урночку по диаметру (рис. 23, 8). Урночка - это наиболее сложно устроенная средняя часть коробочки, в которой развиваются споры (рис. 26). У большинства бриевых мхов в центре урночки проходит колонка, верхняя часть которой у пол штриховых мхов образует эпифрагму - пленку, прикрывающую устье коробочки. Крышечка, прикрывающая урночку сверху, может быть от более или менее плоской до удлиненно-конусовидной, тупой, с бородавочкой, с прямым или изогнутым клювиком (узкой, как бы насаженной верхушкой) (рис. 24).
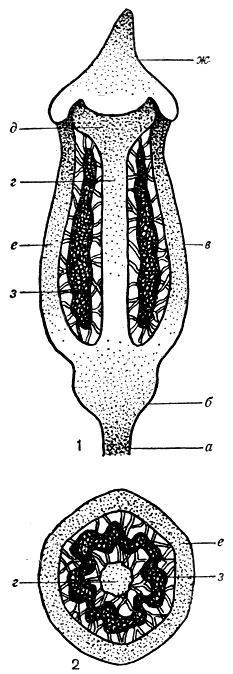
Рис. 26. Строение коробочки политриха на продольном (1) и поперечном (2) срезах: а - ножка, б - шейка, в - урночка, г - колонка, д - эпифрагма, е - стенка, ж - крышечка, з - спорангий
Колпачок (рис. 25) бывает односторонним и кососидящим (двусторонне-симметричным) и прямостоячим (радиально-симметричным). Односторонний, или клобуковидный, колпачок расщеплен с одной стороны. Прямостоячий колпачок может быть шапочковидным (прикрывающим только верхнюю часть коробочки) и колокольчатым (покрывающим всю или почти всю коробочку). Край колпачка бывает ровным, лопастным, бахромчатым или реснитчатым, а поверхность - голой или волосистой. При созревании спорогония колпачок спадает, реже он остается на ножке в виде манжетки (рис. 25, 2).
У большинства бриевых мхов коробочка при созревании спор вскрывается путем отделения крышечки, реже коробочка разрывается на части или споры высвобождаются после сгнивания стенки урночки. По краю урночки после отделения крышечки обычно располагаются небольшие зубцы или реснички, составляющие в своей совокупности окраину, или перистом (рис. 27), строению которого (хорошо различимому лишь с помощью микроскопа) придается большое значение в систематике бриевых мхов. Иногда перистом недоразвит. Если зубцы располагаются в один ряд, перистом называется простым, если в два - двойным, и тогда различают наружный и внутренний перистомы. Число зубцов простого перистома или число зубцов наружного перистома равно 4, 8, 16, 32 или 64. Зубцы простого перистома иногда срастаются попарно или сидят по краю общей довольно высокой основной перепонки. Зубцы перистома совершают гигроскопические движения и этим способствуют высеиванию спор, которые затем распространяются с помощью ветра, воды или насекомых.
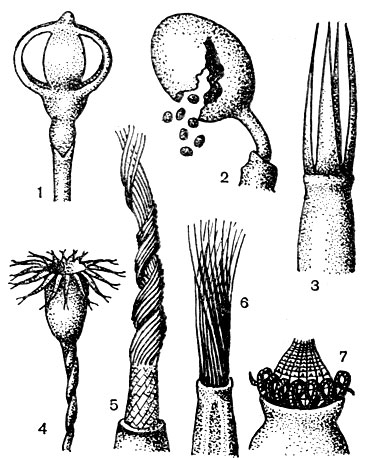
Рис. 27. Коробочки и перистомы андрезевых (1) и бриевых (2-7) мхов: 1 - коробочки на ложноножке, вскрывающаяся 4 створками, 2 - при вскрывании коробочки стенка разрывается на неправильные куски, 3 - простой перистом с 4 зубцами, 4 - простой перистом с 16 двурасщепленными зубцами, 5 - простой перистом со спирально завитыми зубцами на трубчатой основной перепонке, 6 - простой реснитчатый перистом, 7 - двойной перистом
Полная информация на странице http://www.dosugnov.ru/.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'