Физиология и экология почвенных водорослей
Интенсивное развитие почвенных водорослей, как фототрофных организмов, возможно только в пределах проникновения света, в так называемом фототрофном слое. Наибольшее количество водорослей во всех случаях (за исключением нарушенных вспашкой земель) обнаруживается в верхних слоях почвы. В зависимости от влажности почвы и от наличия растительного опада максимум водорослей наблюдается или на самой поверхности почвы или несколько глубже, но в пределах нескольких сантиметров. В нижележащих горизонтах уменьшается как количество видов, развивающихся в культурах, так и количество клеток, выявляемых счетом.
Максимальная глубина, на которой были найдены жизнеспособные водоросли - 2,7 м в окультуренной почве и 1,5 м - в целинной.
Активная жизнь водорослей в почве за пределами проникновения света доказывается прямыми наблюдениями: при микроскопировании почвы просматриваются клетки морфологически целые и нормально окрашенные. Люминесцентное микроскопирование обнаруживает интенсивную флуоресценцию хлорофилла водорослей в пробах, взятых из глубоких слоев почвы.
Почвенные водоросли - но единственный пример жизни водорослей в темноте. Известно нахождение живых водорослей в пещерах на далеком расстоянии от входа, под арктическими льдами и на глубинах океана, в фитопланктоне на такой глубине, где интенсивность света менее 0,06-0,07% от наружной.
Метаболизм почвенных водорослей. Почвенные водоросли не имеют уникальных физиологических свойств, но отражают своеобразие почвенной среды (Lund, 1962). Разнообразие способов питания является особенностью почвенных водорослей.
Водорослям свойственна фототрофия (фртолитотрофия и фотоорганотрофия) и хемотрофия (хемблитотрофия и хемоорганотрофия).
Большинство водорослей представляют собой фотолитоавтотрофы. Многие водоросли способны переходить от одного Фила обмена к другому.
Облигатные автотрофы (фотоавтотрофы), живущие за счет только фотосинтеза и, следовательно, только в пределах проникновения света в заселенный ими субстрат, обнаружены в разных отделах водорослей, хотя предполагается, что их немного среди почвенных водорослей. К ним относятся некоторые виды диатомей, из зеленых - ряд видов хлорококковых и хламидомонад, некоторые цианобактерии.
По поводу облигатной автотрофии цианобактерии существуют различные мнения. С одной стороны существуют многочисленные сведения о том, что синезеленые водоросли - облигатные автотрофы (smith, 1973; Brock, 1973). С другой - есть бесспорные доказательства гетеротрофии у абсолютно чистых культур - представителей Cyanophyta (Carr, 1973; Smith, 1973).
До какой глубины возможно фототрофное питание в почве? Проникновение света, очевидно, ограничивается несколькими миллиметрами. При наличии трещин эта глубина увеличивается до нескольких сантиметров. Проникновение света зависит от свойств почвы, от наземной растительности и толщины опада, от влажности почвы.
Нужно помнить, что многие водоросли являются облигатно теневыми растениями и могут использовать для фотосинтеза свет очень слабой интенсивности. Фотосинтез водорослей возможет при интенсивности света 0,04-0,1% от полного света (см. сводку Голлербах, Штина, 1969).
Облигатные автотрофы способны к разным способам усвоения углерода. Это организмы, у которых восстановление CO2 идет по типу фотосинтеза, фоторедукции или хемосинтеза. Клетки некоторых водорослей могут осуществлять сразу три процесса восстановления CO2 - фотосинтез, фоторедукцию и хемосинтез, продукты которых очень близки. Так Scenedesmus quadricauda и Chlorella vulgaris на свету осуществляют фотосинтез или фоторедукцию, в темноте - хемосинтез.
Фоторедукция - фотосинтез бескислородного типа. Многие цианобактерии осуществляют бескислородный фотосинтез. При этом синтез АТФ в клетке сопряжен только с циклическим электронным транспортом, связанным с фотосистемой I. В качестве экзогенных доноров электронов цианобактерии могут использовать некоторые восстановленные соединения серы (H2S, Na2S2O3, молекулярный водород, ряд органических соединений (сахара, кислоты). Предполагают, что процесс бескислородного фотосинтеза может протекать у цианобактерий двумя путями.
- Фотосистема I индуцирует как циклический, так и нециклический транспорт электронов. Вследствие функционирования обоих электронных потоков фотохимическим путем образуются АТФ и НАДФ·Н2, используемые для ассимиляции CO2 в цикле Кальвина. В этом случае процесс бескислородного фотосинтеза цианобактерий сходен с таковым зеленых бактерий.
- Функция фотосистемы I сводится к получению клеточной энергии в процессе циклического фосфорилирования. Восстановитель образуется в темновых реакциях. Этот вариант близок к схеме фотосинтеза, осуществляемого пурпурными бактериями.
В настоящее время способность к бескислородному фотосинтезу обнаружена у многих цианобактерий из разных групп, но активность фиксации CO2 за счет этого процесса низка, составляя несколько процентов от скорости ассимиляции CO2 в условиях функционирования обеих фотосистем. Только некоторые цианобактерий могут расти за счет бескислородного фотосинтеза, например Oscillatoria limnetica, выделенная из озера с высоким содержанием сероводорода, термофильный штамм Spirulina, компонент микрофлоры термального сероводородного источника (Гусев, Минеева, 1985).
Способность цианобактерий переключаться при изменении условий с одного типа фотосинтеза на другой служит иллюстрацией гибкости их светового метаболизма, имеющей важное экологическое значение. Однако считается, что широкого распространения в природе процессы фоторедукции и хемосинтеза у водорослей не имеют, так как условия для их протекания слишком специфичны.
Давно известна способность многих водорослей к усвоению органических веществ в темноте и на свету в отсутствии CO2. Многие водоросли, особенно зеленые, обладают лабильным обменом веществ и способны перестраивать его от фототрофного до темнового хемогетеротрофного, в зависимости от условий среды. Степень проявления разных типов питания в метаболизме водорослей регулируется различными экологическими факторами, важнейшие из которых - освещенность, состав и количество органических и минеральных веществ в среде, окислительно-восстановительные условия и рН среды.
Способность к гетеротрофному росту отмечена у зеленых, диатомовых, желтозеленых.
В отношении способности цианобактерий к гетеротрофному росту существуют противоречивые мнения. Брок (Brock, 1973) подчеркивает отсутствие роста в гетеротрофных условиях как одну из экологических особенностей синезеленых. Смит (Smith, 1973) и Кар (Carr, 1973) приводят доказательства гетеротрофии у бактериологически чистых культур синезеленых водорослей. Совершенно неспособными к росту в темноте оказались водоросли, лишенные слизистой капсулы. Рост в темноте идет всегда медленнее, чем на свету.
При сравнении водорослей из разных экологических условий наибольший процент сапротрофных видов дали почвенные водоросли (Тоом, 1969).
В экспериментах, доказывающих способность водорослей расти гетеротрофно в темноте, обычно использовались в качестве источника углерода сахара и органические кислоты. Простых органических веществ в почве мало, зато множество почвообитающих микроорганизмов, более конкурентоспособных вследствие своей облигатной гетеротрофности. Водоросли - факультативные гетеротрофы - не способны к разложению сложных веществ, которое производится лишь облигатными гетеротрофами. Поэтому расти в темноте водоросли могут лишь в ассоциации с облигатными гетеротрофами (Parker, 1961).
В отличие от способности жить в темноте полностью за счет органического вещества (хемоорганотрофно), свойственной лишь некоторым водорослям, усвоение тех или иных органических веществ на свету одновременно с фотосинтезом (фотоорганотрофия или миксотрофия), присуще большинству водорослей.
Особое значение для роста имеют органические вещества при условии недостаточной интенсивности света. При слабом освещении органические вещества могут действовать косвенно, через стимуляцию роста бактерий.
У многих цианобактерий экзогенные органические вещества действуют по-разному в темноте и на свету. Anabaena variabilis энергично потребляла глюкозу и ацетат на свету, хотя в темноте не росла ни на одном из органических веществ.
Хемогетеротрофный рост цианобактерий всегда немного медленнее фотогетеротрофного и особенно фотоавтотрофного.
В настоящее время достоверно доказанными и имеющими реальное значение в природе считаются два способа питания почвенных водорослей: 1) фототрофное и миксотрофное питание в пределах проникновения света и 2) гетеротрофное питание в темноте. Способность переходить на гетеротрофный образ жизни в толще почвы наиболее выражена у зеленых и желтозеленых водорослей.
Известны формы водорослей, которые относятся к числу ауксотрофных организмов и нуждаются в экзогенных источниках некоторых физиологически активных веществ, в частности витаминах B12, B1, в биотине.
В отношении азотного питания водоросли подобны зеленым растениям. Они хорошо усваивают минеральные формы азота - нитраты и аммонийные соли. Усвоение NO3 и NH4. клетками водорослей подчиняется тем же закономерностям, которые найдены Д. Н. Прянишниковым для высших растений.
Специфическим способом ассимиляции азота является фиксация молекулярного азота, свойственная некоторым цианобактериям, являющимся уникальными организмами, сочетающими в одной клетке фотосинтез и способность к усвоению молекулярного азота.
Как факультативная форма азотного питания для водорослей показано усвоение азота из органических соединений. В противоположность ограниченной способности использовать органические вещества как источник углерода, многие цианобактерии могут получать азот в органической форме, вырабатывая соответствующие ферменты.
Мочевина, амиды, аминокислоты хорошо используются для азотного питания зелеными водорослями.
По потребности в других элементах и способам их усвоения водоросли принципиально не отличаются от высших растений, хотя у отдельных видов и отделов обнаруживаются специфические особенности.
Фосфор поглощается в виде фосфатов, но доступны и органические соединения, например, аденозинтрифосфат и глицерофосфат.
Сера доступна в виде SO4, SO3, S2O3 а также в виде цистеина, l-цистина и метионина.
Необходимы для водорослей макроэлементы - K, Mg, Ca, Fe, для большинства цианобактерии - Na.
Водорослям необходимы для роста микроэлементы - Mn, Mo, V, Co, Cu, Zn, Br. Диатомовым водорослям необходим кремний. Многим водорослям нужен хлор.
Приспособительные особенности почвенных водорослей. Водоросли, живущие в почве и на ее поверхности, имеют разнообразные морфологические приспособления, обеспечивающие им существование при низкой влажности почвы и сохранение жизнеспособности при высыхании.
Почвенные водоросли - преимущественно пойкилоксерофиты, т. е. растения, не регулирующие водный режим и существующие за счет увлажнения после выпавших осадков.
Ряд морфологических и физиологических особенностей почвенных водорослей обеспечивают поглощение и удержание воды, а также сохранение жизнеспособности при засухе.
- Обильное образование слизи - слизистых чехлов и обверток, широко распространенных у зеленых и синезеленых водорослей. Они состоят из гидрофильных коллоидных полисахаридов и способны поглощать и удерживать воду.
- Мелкие размеры почвенных водорослей. Мелкие клетки легче перемещаются в почве, более устойчивы против засухи, имеют большую водоудерживающую способность, легче распространяются с пылью. Например, в дерново-подзолистой почве (Домрачева, Штина, 1985) клетки с диаметром менее 10 мкм составили 60% общего количества клеток.
- У многих почвенных водорослей отсутствуют зооспоры, жгутиковые формы обладают слабой подвижностью. Период увлажнения после засухи приводит к быстрому образованию гамет и зооспор, например у зеленых, которые обладают фототаксисом и распространяются на поверхности почвы.
- "Эфемерность" вегетации - способность почвенных водорослей быстро переходить из состояния покоя к активной вегетации и наоборот, без образования особых стойких спор и специфических стадий.
- Протопласт обладает физиолого-биохимическими особенностями, которые обеспечивают высокую засухоустойчивость даже видам, не имеющим чехлов. Эти особенности - вязкость протоплазмы и устойчивость против плазмолиза; высокая концентрация клеточного сока и относительно высокое содержание связанной воды в клетке; отсутствие крупных вакуолей и большая сосущая сила, позволяющая использовать даже гигроскопическую влагу воздуха; накопление масла (у диатомей).
- Снижение интенсивности дыхания в клетке по мере ее обезвоживания. При этом дыхание сохраняет свою энергетическую полноценность, что видно по количеству органического кислоторастворимого фосфора, который не разрушается при высыхании у пойкилоксерофитов.
При обезвоживании сохраняется вся организация клетки, происходит "гелефикация протоплазмы", и растение переходит из жизнедеятельного в нежизнедеятельное, но жизнеспособное состояние.
Есть много примеров большой выносливости и длительного сохранения жизнеспособности в сухом состоянии, Nostoc commune был оживлен после 107-летнего хранения в гербарии.
Наиболее устойчивыми к засухе считаются сине зеленые водоросли, особенно спорообразующие, и ряд одноклеточных зеленых, в частности хлорококковые.
Наименее засухоустойчивы диатомеи и желтозеленые водоросли.
Почвенные водоросли распространены до аридных почв, где поверхность почвы нагревается до 50-70°, что заставляет предположить значительную стойкость протопласта этих водорослей к высоким и низким температурам. Теплоустойчивость водорослей зависит от продолжительности действия температуры, от влажности и от природы водорослей, причем особенно устойчивы одноклеточные зеленые.
Устойчивость к экстремальным температурам связана с экологическими условиями, из которых выделены исследуемые водоросли.
Водоросли, выделяемые из лесных почв, заметнее ингибировались высокими температурами, чем пустынные почвенные водоросли.
Как и в отношении засухоустойчивости, отмечаются две группы водорослей, отличающиеся высокой теплоустойчивостью - синезеленые и коккоидные зеленые.
Тепловую границу сохранения жизнеспособности, однако, надо отличать рт температурных границ роста; например, для Chlorella protothecoides тепловая смерть наступала во влажных условиях при 44-46°, в сухих - при 64-67°, а кардинальные температурные точки роста были минимум 5-6°, оптимум - 27°, максимум - 30-31° (Штина, Голлербах, 1976).
Почвенные водоросли выдерживают и максимально низкие температуры. Известны примеры выживаемости при -80°.
Возможно, что гибель при замораживании связана с потерей воды и поэтому одни и те же виды бывают одновременно засухоустойчивыми и холодоустойчивыми.
Среди почвенных водорослей нет отдела, в котором бы не встречались холодостойкие виды, наибольшее количество холодостойких форм известно среди зеленых и синезеленых водорослей; они преобладают в альгофлоре Антарктиды.
Среди почвенных водорослей есть светолюбивые и тенелюбивые формы; часто светолюбив связано с засухоустойчивостью. Теневыносливыми считаются желтозеленые водоросли, поскольку они обычно развиваются в подповерхностных слоях почвы. Однако, желтозеленые могут интенсивно развиваться и на открытой поверхности почвы, вызывая ее "цветение". Диатомовые водоросли могут активно передвигаться в почве в зависимости от степени освещения. Устойчивостью против ультрафиолетового облучения обладают синезеленне водоросли. Этим пользуются для получения бактериологически чистых культур, поскольку УФ-лучи губят бактерии. УФ-облучение в течение 30 с - 5 мин стимулировало рост и азотфиксирующую активность цианобактерий.
Почвенные водоросли проявляют активность против радиоактивных излучений. При исследовании разрушенной взрывом почвы в районе ядерных испытаний в штате Невада обнаружено, что первыми в ней из растений появились водоросли. Microcoleus vaginatus был обнаружен через 3 мес после взрыва на расстоянии 0,6 мили от места взрыва, через 10 мес - на расстоянии 0,4 мили. Другие растения появились лишь через 2 года в 0,6 мили от эпицентра. γ-радиацию от Co60 используют для очистки синезеленых от бактерий.
При росте не засоленных почвах водоросли проявляют еще одну черту высокой толерантности - солеустойчивость. Клетки Zygogonium ericetorum остаются неплазмолизированными в 10%-ном растворе солей морской воды с осмотической концентрацией 75 атм.
Водоросли устойчивы к отдельным токсическим солям или ионам - они поселяются на поверхности зольных промышленных отвалов, содержащих Al2O3, в отвалах с мышьяком и цианистым калием (Штина, 1984).
Таким образом, почвенные водоросли способны существовать даже при крайне неблагоприятных условиях среды. Лабильность обмена веществ, высокая устойчивость против засухи, резких колебаний влажности и температуры, против недостатка и избытка света, засоленности почвы и т. п. объясняют широкое распространение почвенных и наземных водорослей и быстроту их разрастания даже при кратковременном проявлении необходимых факторов.
Жизненные формы почвенных водорослей. Э. А. Штина и М. М. Голлербах (1976) применили к почвенным водорослям понятие жизненной формы, используемое как единица экологической классификации растений, под которым подразумевается группа растений со сходными приспособительными структурами.
Выделяются 9 жизненных форм (экобиоморф) почвенных водорослей (рис. 9). Для их обозначения взяты в качестве индексов первые буквы названий таксонов, рассматриваемых в качестве эталона данной жизненной формы.

Рис. 9. Жизненные формы почвенных водорослей: 1 - Chlorococcum - форма (Ch-форма); 2 - Cylindrospermum - форма (С-форма); 3 - Xanthophyta - форма (X-форма); 4 - Bacillariophyta - форма (В-форма; 5 - Phormidium - форма (P-форма); 6 - Microcoleus - форма (M-форма); 7 - Heterothrix - форма (H-форма); 8 - Nostoc - форма (N-форма); 9 - Vaucheria - форма (V-форма)
По оставшемуся неопубликованным предложению М. М. Голлербаха, даются двойные - развернутые и краткие - обозначения жизненных форм.
1) Chlorococcum - форма (Ch-форма) - одноклеточные и колониальные зеленые и частично желтозеленые водоросли, обитающие в толще почвы, но при благоприятной влажности дающие разрастания и на поверхности почвы. Это виды, отличающиеся выносливостью к различным экстремальным условиям, обычно обозначаемые как "убиквисты". Они первыми начинают заселение почвы или материнской породы, заносятся в глубокие слои почвы, непереваренными проходят через кишечники беспозвоночных. Название происходит от рода Chlorococcum. Сюда не относятся виды порядка Chlamydomonadales и некоторые колониальные формы, образующие обильную слизь. В эту группу включаются повсеместно встречающиеся виды одноклеточных зеленых (Chlorococcum, Bracteacoccum, Myrmecia, Chlorella), желтозеленых (Pleurochloris magna, Botrydiopsis arhiza) И даже синезеленых (Synechococcus).
2) Cylindrospermum - форма (C-форма) включает одноклеточные, колониальные или нитчатые формы, которые могут образовывать обильную слизь. Обитают как в толще почвы, так и формируют тонкие слизистые пленки или хлопья на поверхности почвы. В отличие от Ch-формы, эти формы более требовательны к воде и переносят высыхание в виде спор, зигот, реже - в вегетативном состоянии. В большинстве это теневыносливые виды и на поверхности почвы развиваются под укрытием высших растений. Эта форма включает виды из разных отделов: Cylindrospermum, Anabaena, Nostoc, Gloeocapsa, Gloeothece, Microcystis - из синезеленых; Chlamydomonas - из зеленых.
3) Xanthococcophyceae - форма (X-форма) - большинство одноклеточных желтозеленых и многие зеленые, предпочитающие теневые условия среди почвенных частиц, теневыносливые, но не устойчивые против засухи и экстремальных температур. Им свойственна способность к миксотрофному питанию. Отмечено большое их разнообразие во влажных торфяных почвах (Голлербах, Штина, 1969).
4) Bacillariophyta - форма (B-форма) - диатомовые водоросли - подвижные клетки, живущие в самых поверхностных слоях влажной почвы или в слизи других водорослей. Холодостойкие, светолюбивые, часто солевыносливые, но не устойчивые против высыхания.
5) Phormidium - форма (P-форма) - нитевидные синезеленые (Phormidium, Oscillatoria, Lyngbya, Plectonema), не образующие значительной слизи. Они рассеяны в толще почвы, оплетая почвенные частицы, или образуют на поверхности тонкие кожистые пленки, ксерофиты.
6) Microcoleus - форма (M-форма) - синезеленые в виде более или менее слизистых нитей, образующие макроскопически заметные корочки или дерновинки на поверхности почвы: Microcoieus, Schizothrix, Hydrocoleus. Отличаются засухоустойчивостью, теплоустойчивостью и холодоустойчивостью.
7) Heterothrix - форма (H-форма) - нитевидные зеленые и желто-зеленые. Большинство видов Chlorhormidium, виды Ulothrix, Stichococcus, Bumilleria, Tribonema. Не устойчивы против засухи, сильного света. Живут рассеянными среди почвенных частиц или, при достаточной влажности и затенении, образуют поверхностные налеты. Могут покрывать основания стеблей. Входят в состав водорослевых ассоциаций, образующих "цветение" почвы. В качестве эталона H-формы взята желтозеленая водоросль Heterothrix.
8) Nostoc - форма (N-форма) - наземные микроскопические талломы N. commune, N. sphaeroides, N. microscopicum, N. flagelliforme; световыносливые и засухоустойчивые виды - пойкилоксерофиты. Слизистые талломы ностоков служат местообитанием многих бактерий, грибов, одноклеточных водорослей.
9) Vaucheria - форма (V-форма) - нитевидные водоросли, образующие войлокообразные налеты на поверхности влажных почв. Название от рода Vaucheria. По соотношению числа видов различных жизненных форм определяют экологические спектры альгофлоры в той или иной почве.
Ценология почвенных водорослей. Водоросли составляют структурно и экологически оформленную часть биогеоценоза. При характеристике водорослевых группировок (водорослевые группировки иногда называют сообществами, альгоценозами, альгосинузиями) отмечают признаки, используемые в геоботанике для описания фитоценозов.
- Видовой состав. Определяется культуральными методами.
- Доминантные виды.
- Количественная характеристика сообщества. Определяется общее количество клеток водорослей, соотношение численности отдельных групп, биомасса.
- Встречаемость отдельных видов или групп водорослей.
- Специфические виды или группы видов.
- Вертикальное сложение сообщества водорослей (распространение водорослей в профиле почвы).
- Сезонная динамика.
Наиболее сложно определить доминантные виды и встречаемость отдельных видов сообщества.
В макроскопически заметных поверхностных разрастаниях водорослей доминанты выявляются сравнительно легко путем прямого микроскопирования разрастаний. Водорослевые группировки в этих случаях называются по доминирующим видам. Например "ностоко-сцитонемовый ценоз степей и полупустынь", "формидиевые пленки", "ностоковый ценоз".
В случае, если водоросли диффузно рассеяны среди почвенных частиц, труднее выделить доминирующие виды. Для этого используются культуральные методы и сочетание определения видового состава водорослей на "стеклах обрастания" с учетом количества водорослей в исходной почве с помощью метода прямого счета (Некрасова, Бусыгина, 1977). На "стеклах обрастания" просматривается определенное количество полей зрения, устанавливаемое предварительно на основании равномерности распределения водорослей, и в каждом поле зрения отмечается обилие и встречаемость отдельных видов. Каждое поле зрения принимается за повторность, равноценную учетной площадке в геоботанических исследованиях, затем вычисляется относительная средняя встречаемость родов. На основании этого показателя рассчитывают количество отдельных видов или родов в общей массе водорослей и получают представление о доминирующих формах.
Состав водорослей разных зональных почв дан в табл. 1.
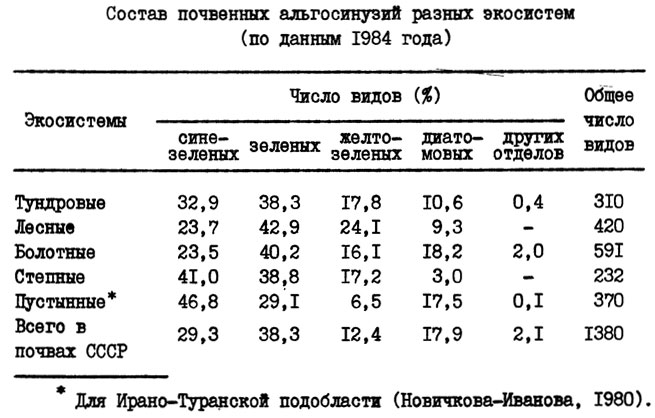
Таблица 1. Состав почвенных альгосинузий разных экосистем (по данным 1984 года)
Соотношение числа видов разных отделов, в частности синезеленых и зеленых, использовано для уточнения некоторых географических закономерностей. По предложению Л. Н. Новичковой-Ивановой (1980), соотношение синезеленых и зеленых во флорах рассматривается как показатель аридности: чем больше перевес синезеленых, тем выше аридность (в материалах автора этот показатель колеблется от 3,6:1 до 1,9:1).
Группировки водорослей разных почв. Специфичность водорослевых группировок разных процессов почвообразования представляется в следующем виде.
Подзолообразовательный процесс - сравнительно простые альгосинузии с преобладанием зеленых и желтозеленых водорослей, ограниченное число видов и небольшая биомасса. Дерновый процесс - разнообразие видов всех систематических групп, интенсивное развитие азотфиксирующих синезеленых и мелких диатомей, распространение водорослей по всему дерновому горизонту и значительная биомасса. Болотный процесс - большое разнообразие зеленых, слабое развитие синезеленых, значительная примесь гидрофильных видов, особенно диатомовых. Степной процесс - преобладание синезеленых и одноклеточных зеленых - убиквистов; в отдельные периоды - развитие на поверхности почвы "ностоко-сцитонемового ценоза", небольшая роль зеленых. Солонцовый процесс - при сохранении основных черт альгосинузии зональных почв - более значительное развитие зеленых и особенно диатомовых водорослей. Осолодение - обилие зеленых (в том числе крупных нитчаток) и диатомовых, слабое развитие синезеленых. Пустынный процесс (применительно к глинистым пустыням) - сокращение разнообразия и численности зеленых и особенно желтозеленых, доминирование синезеленых из порядка Oscillatoriales.
Окультуривание почвы резко изменяет почвенную альгофлору и стирает ее зональные особенности. Поэтому сообщества водорослей, характерные для данной зоны, обнаруживаются только в целинных почвах под естественными фитоценозами (табл. 1). В таблице приводится число видов, многие из которых включают значительное число внутривидовых таксонов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'