19. Отдел 10. Зеленые водоросли - Chlorophyta (Chlorophycophyta, Chlorophyceae, Chlorobionta, Viridiplantae)
19.1. Общая характеристика (Н. П. Масюк)
Отдел объединяет свыше 20000 видов микро- и макроскопических эукариотических водорослей, по окраске напоминающих высшие растения. Мельчайшие (например, Micromonas pusilla (Butch.) Manton et Parke) граничат по размерам с бактериальными клетками, не превышая нескольких микрометров в диаметре, наиболее крупные - харовые - достигают - 2 м в высоту. Представлены одноклеточные, колониальные (в т. ч. ценобиальные), многоклеточные и неклеточные индивиды, активно подвижные и неподвижные, прикрепленные и свободноживущие, а также все типы структуры вегетативного тела от монадного до паренхиматозного, исключая амебоидный (рис. 19.1).
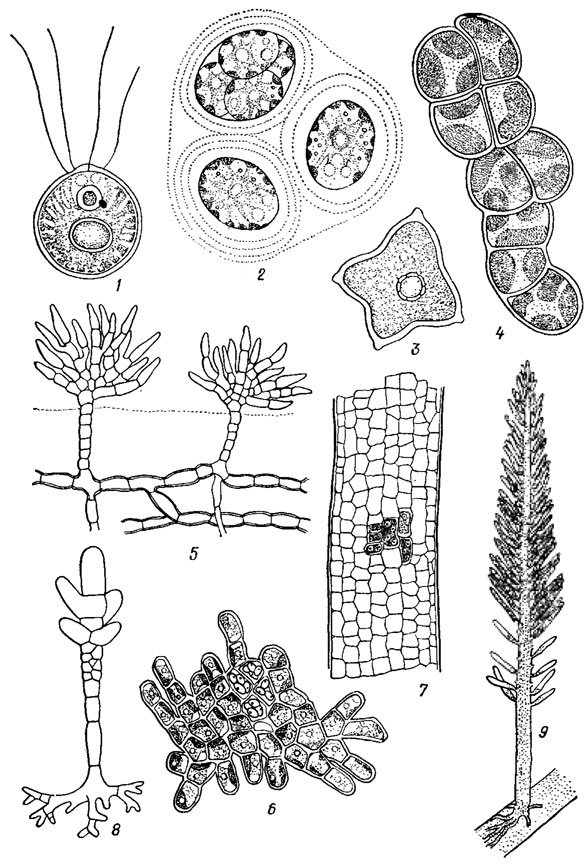
Рис. 19.1. Типы структуры вегетативного тела зеленых водорослей: 1 - монадный (Carteria radiosa Korsch.); 2 - гемимонадный (Asterococcus superbus (Cienk.) Scherff.); 3 - коккоидный (Tetraedron minimum (A. Br.) Hansg.); 4 - нитчатый (Pleurochloris vulgaris Menegh.); 5 - разнонитчатый (Fritschiella tuberosa Iyengar); 6 - псевдопаренхиматозный (Protoderma viride Kütz.); 7 - паренхиматозный (Enteromorpha pilifera Kütz.); 8 - сифонокладальный (Siphonocladus pusillus (Kütz.) Hauck.); 9 - сифональный (Bryopsis Lamour.)
Строение клетки и клеточные покровы разнообразны. У некоторых клетки окружены лишь плазмалеммой (Quadricnloris Fott), у других снабжены дополнительными образованиями в виде субмикроскопических чешуек (Prasinophyceae Chad.) или структур, напоминающих перипласт [279]. Такие клетки метаболичны. У большинства зеленых водорослей образуются клеточные оболочки, сохраняющие более или менее постоянную форму клеток. В световом микроскопе в оболочке многих зеленых водорослей видны два слоя - внутренний, более плотный, обычно целлюлозный и наружный, пектиновый. Клеточная оболочка Haematococcus С. A. Ag. по данным электронной микроскопии, состоит из четырех слоев, Chlamydomonas reinhardtii Dang. - из семи, Pleurotaenium nodosum (Bailey) Lund - из многочисленных слоев. Она представлена аморфным матриксом и погруженными в него микрофибриллами целлюлозной природы, количество и ориентация которых меняются от слоя к слою.
Клеточная оболочка может составлять до 39,2% сухой массы клетки (Hydrodictyon Roth); 7,2% этого количества приходится на жиры, остальное - на белки и углеводы. Целлюлоза - основной компонент клеточной оболочки большинства зеленых водорослей. Однако у Chlamydomonas Ehr. она отсутствует; в состав клеточных оболочек у этого рода входит гликопротеин. Протеин в оболочках других зеленых водорослей составляет 10-69% их массы. У Prasinophyceae оболочки состоят из пектина, галактозы и арабинозы. Основную часть клеточных оболочек Pleurotaenium Näg. слагают глюкоза, галактоза, ксилоза, арабиноза, глюкуроновая и галактуроновая кислоты. У Bryopsidalps оболочки состоят из маннанов и ксиланов, причем у спорофитов преобладает маннан, у гаметофитов - ксилан и целлюлоза. В клеточных оболочках Chara L., Pediastrum Meyen, Chlorella Beijer., помимо внутреннего целлюлозного, обнаружен спорополлениновый слой, состоящий из окисленных полимеров каротина и (или) каротиноидных эфиров, которые входят в состав клеточных оболочек спор и пыльцы высших растений. Попытки обобщения данных о химическом составе клеточных оболочек и использования их в систематических целях затрудняются недостаточной изученностью зеленых водорослей и отсутствием необходимого количества сравнительного материала [385, 393].
Оболочки могут быть инкрустированы солями железа (Phacotaceae) или кальция (Charophyceae, Bryopsidales). У видов рода Oedogonium Link на поверхности оболочки выделяется дополнительный твердый покров (кутикула), состоящий из кутина.
Клеточная оболочка обычно цельная, реже состоит из двух - нескольких фрагментов (Microspora Thur., Desmatractum West, Desmidiales). На поверхности оболочки нередко образуются разнообразные выросты, шипы, щетинки, гранулы, бородавки (см. рис. 19.17), выполняющие защитную функцию и облегчающие парение в воде.
Обмен клеток с окружающей средой происходит через поры, образующиеся вследствие неполного смыкания оболочек при цитокинезе (первичные) или под воздействием ферментов (вторичные поры). Диаметр пор у Desmidiales 12-20 нм. У некоторых колониальных (Volvox L., Pediastrum) и многоклеточных (Ulotrichales, Oedogoniales) зеленых водорослей через поры связь между соседними клетками осуществляется посредством плазмодесм, у других - через поры выделяется слизь, способствующая образованию колоний (Hormotila Borzi, Palmodictyon Kütz.) или активному перемещению организма в пространстве (Closterium Nitzsch.). У некоторых представителей порядка Volvocales слизь окружает протопласт внутри клеточной оболочки (Haematococcus С. A. Ag.). Нередко слизь возникает вследствие ослизнения клеточных оболочек (Botryococcus Kütz.). Образование слизистых капсул вокруг клеток свойственно конъюгатам. Иногда (Chlamydomonas, Ulothrix Kütz.) при неблагоприятных условиях наблюдается временное ослизнение клеточных оболочек и переход в пальмеллевидное состояние.
У некоторых Prasinophyceae обнаружены стрекательные структуры, напоминающие эджектосомы криптомонад.
Клетки содержат от одного (Chlamydomonas) до нескольких сотен (Cladophora Kütz.) ядер; размеры их колеблются от одного (Chlorella, Chlorococcum Menegh., Oocystis Näg.) до 45 мкм (Bryopsis halimeniae Berth.), форма обычно шаровидная или линзовидная. В нуклеоплазме от 1 до 100 (Micrasterias thomasiana Arch.) ядрышек; число, форма и размеры их изменяются на протяжении клеточного цикла. Многообразие интерфазных ядер проявляется в соотношении объема ядрышка и нуклеоплазмы, степени спирализации хромосом. Наряду с типичными картинами митоза выявлены разнообразные отклонения, связанные с разной степенью выраженности веретена деления, наличием или отсутствием центриолей, поведением ядер ной оболочки и ядрышка, микротрубочек веретена. Иногда наблюдается внутриядерный митоз. В ядрах Spirogyra Link ядрышки во время митоза диспергируют, маскируя их содержимое [279, 3911. У зеленых водорослей наблюдается значительно большее разнообразие картин митоза по сравнению с высшими растениями [278].
Хлоропласты разнообразны по форме, размерам, положению в клетке. Их количество варьирует от одного (многие виды Chlamydomonas) до нескольких десятков (Sphaeroplea Ag.) или сотен (Cladophora) в клетке. По форме различают чашевидные, пластинчатые, корытовидные, лентовидные, дисковидные, сетчатые, губчатые, звездчатые хлоропласты, по положению - пристенные и осевые (центральные) (рис. 19.2). Хлоропласты окружены двухмембранной оболочкой, не связанной с каналом эндоплазматического ретикулюма и ядерной оболочкой; тилакоиды в группах по 2-6 или в плотных стопках, напоминающих граны высших растений [391].
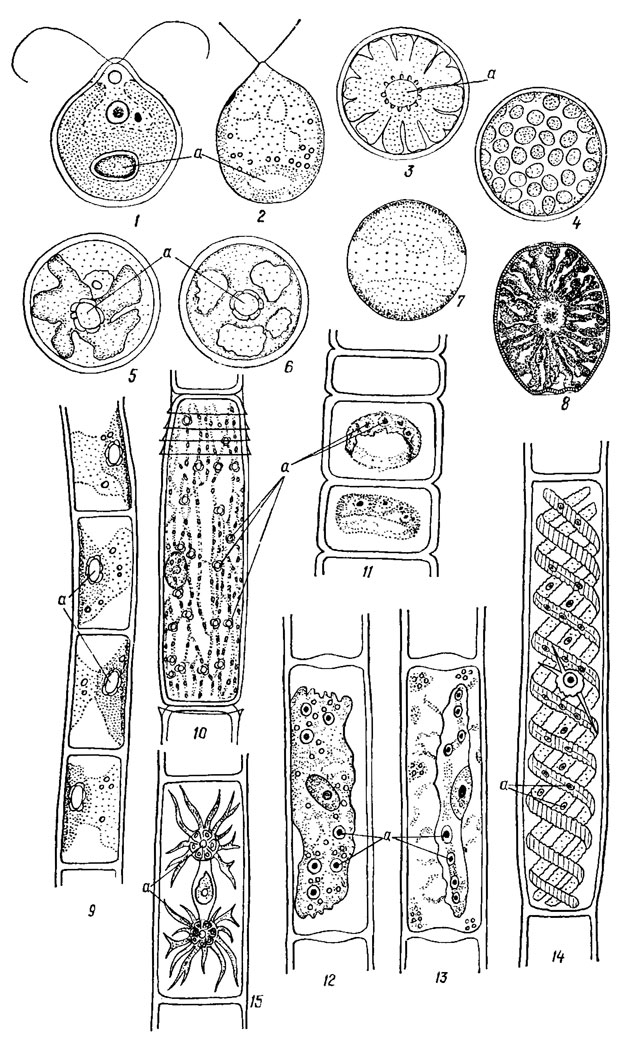
Рис. 19.2. Хлоропласты зеленых водорослей: 1 - чашевидный с базальным пиреноидом (Chlamydomonas proboscigera Korsch.); 2 - чашевидный перфорированный с базальным пиреноидом (Dunallella carpatica Massjuk); 3 - звездчатый, осевой с центральным пиреноидом (Radiosphaera dissecta (Korsch.) Starr); 4 - пристенные дисковидные без пиреноидов (Bracteacoccus sp.); 5 - асимметрично звездчатый, осевой с центральным пиреноидом (Axylosphaera vegetata Cox et Deas.); 6 - губчатый с центральным пиреноидом (Neospongiococcum punctatum (Arcet et Bold) Deas.); 7 - пристенный, разделенный надвое трещиной, без пиреноидов (Lobococcus macronucfeatus (Peas.) Bourr.); 8 - радиально расположенные, лентовидные (Oocystis gigas Arch. var. incrassata W. et G. S. West sensu Skuja); 9 - пристенный полуцилиндрический с пиреноидами (Chlorhormidium flaccidum (Kütz.) Fott); 10 - пристенный, сетчатый с многочисленными пиреноидами (Oedogonium crassum (Hass.) Wittr.); 11 - кольцевидный со многими пиреноидами (Ulothrix zonata (Web. et Mohr.) Kütz.); 12, 13 - осевые, пластинчатые с несколькими пиреноидами (Mougeotla sp.); 14 - три пристенных, лентовидных, спиралевидно изогнутых хлоропласта с пиреноидами (Spirogyra sp.); 15 - два осевых, звездчатых хлоропласта с пиреноидами (Zygnema sp.); а - пиреноид
Хлоропласты обычно окрашены в различные оттенки зеленого цвета. Окраска обусловлена наличием хлорофиллов а и b, α-, β-, γ-, ξ-каротинов и их окисленных производных - ксантофиллов: лютеина, неоксантина, виолаксантина, зеаксантина, антераксантина. У некоторых зеленых водорослей обнаружен криптоксантин, представителям Siphonophyceae свойственны сифоненн и сифоноксантин, у Prasinophyceae выявлены микронон и ликопин, последний отмечен и у Charophyceae. По составу фотосинтетических пигментов зеленые водоросли близки к высшим растениям. В неблагоприятных условиях (недостаток биогенных элементов, повышение концентрации хлористого натрия и др.) соотношение основных групп пигментов в вегетативных клетках некоторых зеленых водорослей изменяется в пользу каротиноидов, водоросли приобретают оранжево-красную окраску (Trentepohlia Mart., Dunaliella salina Teod., Chlamydomonas nivalis (Bauer) Wille), отдельные представители способны накапливать вторичные каротиноиды: эхиненон, каитаксантин, астаксантин, астацин (Chlorococcum, Spongiochloris Starr, Dunaliella pseudosalina Massjuk et Radch.). Подобные изменения пигментации нередко наблюдаются в покоящихся клетках - зиготах, акинетах. Способность к синтезу вторичных каротиноидов используется в таксономических целях на низших таксономических уровнях [195, 502 и др.]. У некоторых зеленых водорослей наблюдается утрата фотосинтетических пигментов (апохлороз) при сохранении в клетках бесцветных лейкопластов (Polytomella Aragao, Tetrablepharis Senn, Hyalogonium Pasch.), а иногда н утрата пластид - апопластия (Polytoma Ehr., Hyalobrachion Swindell, Hyaliella Pasch.). Функцию хлоропластов в бесцветных клетках могут выполнять внутриклеточные симбиотические синезеленые водоросли - цианеллы (Hyalocardium printzii (Schill.) Ettl, Gloeochaete wittrockiana Lagerch., Glaucocystis nostochinearum Itz.). У Dasycladaceae наряду с хлоропластами присутствуют амилопласты.
Хлоропласты обычно содержат от одного до нескольких десятков (Draparnaldia Bory, Cladophora Kütz.) погруженных пиреноидов. Их число и локализация в пластиде обычно видоспецнфичны. Ламеллярная система хлоропласта нередко проникает в пиреноид (Pyramimonas Schmarda, Cosmarium Corda, Mougeotia Ag.), у иных (Scenedesmus quadricauda (Turp.) Breb., Oedogonium cardiacum Wittr., Chlamydomonas chlamydogama Bold) пиреноиды лишены ламеллярной системы. Попытки связать особенности тонкого строения пиреноида с систематическим положением объекта пока не увенчались успехом. Только у одного рода Ulothrix наблюдается 8 типов пиреноидов [540]. Полагают, что пиреноид зеленых водорослей является ферментативным центром, продуцирующим энзимы, полимеризующие молекулы глюкозы* в крахмал - основной продукт ассимиляции зеленых водорослей**. Крахмал образует вокруг пиреноида сплошную или состоящую из отдельных зерен обкладку. Наличие светопреломляющей крахмальной обкладки облегчает обнаружение пиреноида. Пиреноиды некоторых зеленых водорослей (Monoraphidium mirabile (W. et G. S. West) Pankow, Kirchneriella Schmidle, Ankistrodesmus Corda) лишены крахмальной обкладки, и выявить их гораздо труднее.
* (Основные ассимиляты зеленых водорослей - глюкоза, фруктоза и сахароза. Prasinophyceae синтезируют маннитол и глюкозоманнитол, для аэрофитных водорослей (Trentepohlla Mart. и др.) характерны сахарные спирты.)
** (Крахмал зеленых водорослей (α-1, 4-глюкан), как и у высших растений, состоит из амилозы и амилопектина (в различных пропорциях). Лишь у Dasycladaceae вместо крахмала откладывается инулин и близкие полифруктозиды. Кроме крахмала зеленые водоросли могут накапливать масло, иногда в большом количестве, маскирующем наличие крахмала (Botryococcus braunii Kütz.). Гипергалобная водоросль Dunaliella salina Teod. в больших количествах синтезирует глицерин.)
Крахмал может откладываться вне связи с пиреноидом; локализация его в клетках зеленых водорослей, как и высших растений, строго приурочена к пластидам. Лишь у Dasycladaceae запасные полисахариды могут откладываться за пределами пластид. Пиреноиды воспроизводятся делением надвое или фрагментацией. Обычно деление их по времени совпадает с завершением митоза, делением пластид и клетки, хотя не исключены отклонения. При спорогенезе наблюдается новообразование пиреноидов, которому предшествует исчезновение старого пиреноида.
У монадных форм и стадий зеленых водорослей, а также у некоторых гемимонадных, филогенетически связанных с монадными предками, имеется стигма, являющаяся частью хлоропласта. Стигма в виде небольшого красного пятна расположена в передней части клетки, реже медиально. Она состоит из одного - нескольких слоев пигментированных глобул, расположенных у поверхности хлоропласта, прилегающей к поверхностным покровам клетки, и разделенных тилакоидами (см. рис. 1.34). У Chlamydomonas Ehr. стигма состоит из 15-45 электронно-плотных гранул 75-100 нм каждая, которые тесно прилегают друг к другу, образуя один слой. Место фоторецептора в клетках зеленых водорослей с точностью не установлено. В настоящее время считают, что функцию фоторецепции выполняет участок наружной мембраны хлоропластной оболочки или плазмалеммы, прилегающий к стигме и отличающийся от соседних участков мембраны, стигме же отводят вспомогательную роль в фоторецепции. В пользу этого свидетельствует способность к фототопотаксису некоторых монадных форм и стадий зеленых водорослей, лишенных стигмы (зооспоры Chlorhormidium flaccidum (Kütz.) Fott. мужские гаметы Bryopsis hypnoides Lamour., мутанты Chlamydomonas Ehr.).
Монадные формы и стадии обычно снабжены 2-4 изоморфными (акронематическими), изоконтными, изодинамическими жгутиками (Dunaliella Teod., Chlamydomonas, Carteria Dies.) (рис. 19.3). Количество Жгутиков может варьировать от одного (Pedinomonas Korsch.) до многих (монадные стадии Oedogonium, зооспоры Derbesia Sol.). Наряду с гладкими акронематическими наблюдаются плевронематические жгутики, покрытые одним (Haematococcus) или большим количеством рядов мастигонем (Tetraselmis Stein, Pedinomonas Korsch.). Жгутики Prasinophyceae и Charophyceae покрыты субмикроскопическими чешуйками. У Nephroselmis Stein и Trichloris Scherff. et Pasch. жгутики гетероконтные и гетеродинамические. Жгутики прикрепляются апикально, реже субапикально к морфологически переднему концу клетки, который не всегда соответствует переднему по ходу движения концу клетки. Иногда жгутики выходят из апикальной жгутиковой ямки (Tetraselmis) (рис. 19.3, 4) или субапикального желобковидного углубления. Аксонема жгутиков имеет типичное для эукариот расположение микротрубочек (структура "9 + 2"). В месте погружения жгутиков в клетку периферические дуплеты микротрубочек замещаются триплетами, образующими базальные тела. В переходной зоне жгутика имеется комплекс структур, соединяющих дуплеты микротрубочек, которые в поперечном сечении образуют звездчатую фигуру. Базальные тела располагаются под углом 90 или 180°. Они соединены поперечно исчерченной или неисчерченной перемычкой (рис. 19.4). С базальными телами связаны три типа структур: корни, ризопласты и многослойные структуры, расположенные в цитоплазме (см. рис. 19.4). Корни имеют микротрубчатую, ризопласт - микрофибриллярную, поперечно полосатую структуру. Ризопласт может быть связан с ядром. Многослойная структура, обнаруженная, например, у Charophyceae, Coleochaete Breb. - сложно организованная группа микротрубочек, расположенных вблизи от базальных тел; она напоминает таковую в подвижных репродуктивных клетках некоторых высших растений. От многослойной структуры отходит унилатеральный микротрубчатый корень (см. рис. 19.4).
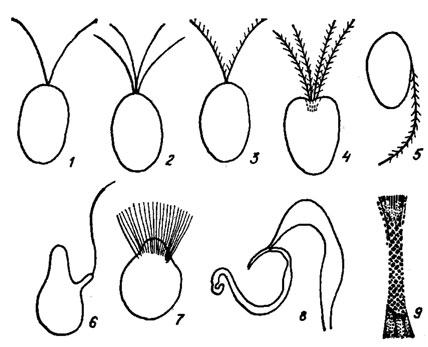
Рис. 19.3. Жгутики зеленых водорослей: 1 - Chlamydomonas Ehr.; 2 - Carteria Dies.; 3 - Haematococcus С. A. Ag.; 4 - Tetraselmis Stein; 5 - Pedinomonas Korsch.; 6 - Micromonas Manton et Parke; 7 - Oedogonium Link (зооспоры, андроспоры, антерозоиды); 8 - Chara (Vaill.) L. (антерозоиды); 9 - субмикроскопические чешуйки на жгутике антерозоида Chara (Vaill.) L
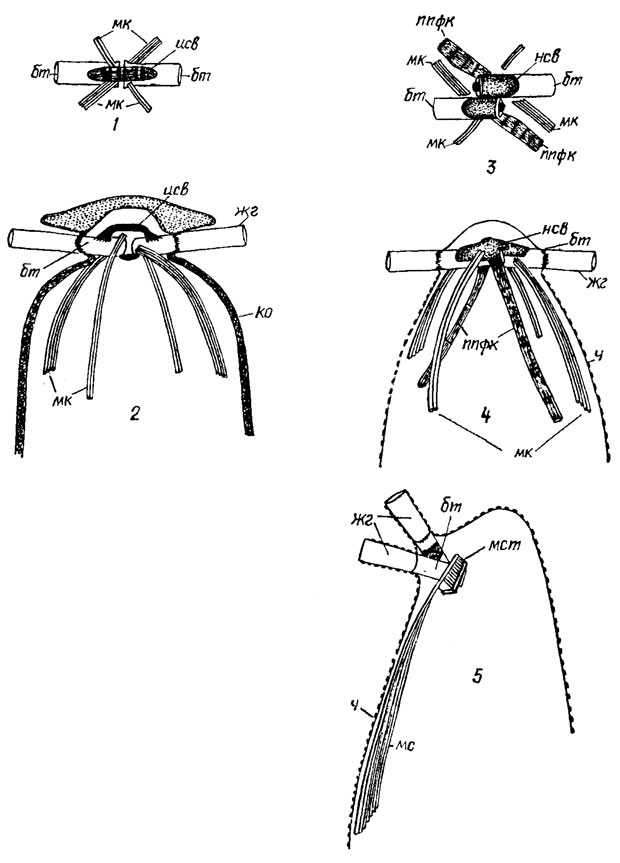
Рис. 19.4. Организация жгутикового аппарата монадных форм и стадий зеленых водорослей из классов Chlorophyceae s. str. (1, 2), Ulvophyceae (3, 4), Charophyceae (5), вид сверху (1, 3) и сбоку (2, 4, 5): бт - базальное тело; мк - микротрубчатые корни; исв - исчерченное соединительное волокно; жг - жгутик; ко - клеточная оболочка; ппфк - поперечно-полосатый фиброзный корень; нсв - неисчерченное соединительное волокно; лес - микротрубчатый сплин; мст - многослойная структура; ч - чешуйки
У основания жгутиков монадных форм и стадий находятся диктиосомы (аппарат Гольджи). В неподвижных клетках диктиосомы обычно располагаются вокруг ядра; у Bryopsidales - преимущественно в меристоплазме. В клетках Pediastrum Meyen, Tetracystis Brown et Bold, Chlorella они заключены в выросты наружной мембраны ядерной оболочки - амплексы. Размеры диктиосом колеблются от 0,7 (Spirogyra) до 20 мкм (Cladophora). Число цистерн в диктиосоме у Cosmarium и Closterium 8-12, у Oocystis 12-16. Диктиосомы участвуют в синтезе клеточных покровов, в секреторной деятельности клетки, возможно, в образовании лизосом.
Число митохондрий в клетках колеблется от одной (Chlorella, Chlamydomonas noctigama Korsch.) до нескольких и может изменяться в онтогенезе. Размеры митохондрий варьируют в пределах одного растения, иногда достигая больших значений (Bryopsis Lamour.). Митохондрии зеленых водорослей имеют пластинчатые кристы. Являясь источником энергии, митохондрии располагаются в активных центрах клетки.
Разветвленная система канальцев эндоплазматического ретикулюма располагается в клетках неравномерно, концентрируясь возле стигмы (Volvox) или вокруг хлоропласта и диктиосом (Closterium littorale Gay), на периферии клетки, вдоль плазмалеммы (Fritschiella Iyengar, Chlorella), либо вокруг ядра, образуя дополнительную перинуклеарную оболочку (Hydrodictyon, Kirchneriella, Tetrasdron Kütz.). Установлена связь эндоплазматического ретикулюма с комплексом Гольджи, митохондриями и хлоропластом, иногда наблюдается контакт с наружной мембраной в ядерной оболочке. У водорослей монадной и гемимонадной структуры наблюдаются сократительные вакуоли, число и распределение которых в клетке видоспецифичны, у остальных - вакуоли с клеточным соком (тонопласт), нередко занимающие в клетке центральное положение (Spirogyra). В цитоплазме обнаружены лизосомы, пероксисомы, сферосомы, ирисовые тела, ломасомы, а также система микротрубочек, существенно не отличающаяся от таковой у представителей других отделов.
В зависимости от степени согласованности процессов митоза и цитокинеза наблюдается последовательное (в вегетативных клетках Chlamydomonas Ehr., Ulothrix Kütz., Spirogyra Link), одновременное (обычно в спорангиях, гаметангиях) и сегментационное (в вегетативных клетках Siphonocladales) клеточное деление (рис. 19.5). Цитокинез осуществляется перетяжкой (кольцевой бороздой) или с помощью клеточной пластинки, формирование которой происходит по типу фико- или фрагмопласта (рис. 19.6). Деление с помощью фрагмопластоподобных структур, напоминающих таковые высших растений (см. рис. 19.6, 4), наблюдается у Chlorhormidium Fott, Coleochaete, Charophyceae, рассматриваемых в качестве представителей прогрессивной линии эволюции зеленых водорослей [589, 660, 661 и др.].
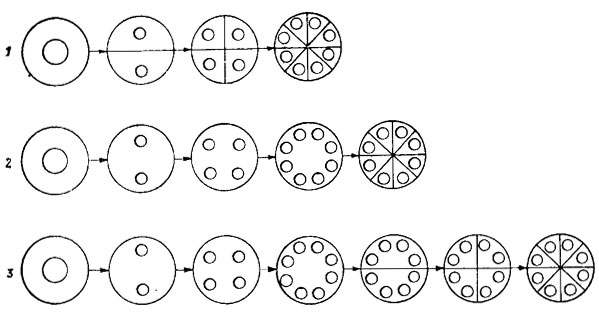
Рис. 19.5. Способы клеточного деления зеленых водорослей: 1 - последовательное, или сукцессивное; 2 - одновременное, или симультанное; 3 - сегментационное, или сегрегативное
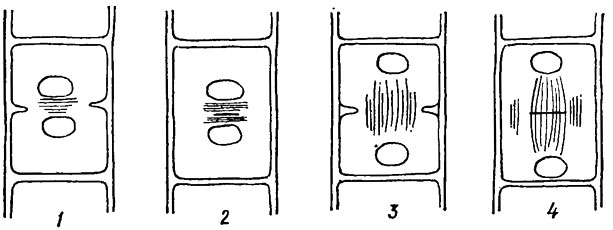
Рис. 19.6. Способы деления клеток зеленых водорослей: 1, 2 - веретено деления отсутствует, ядра сближены (1 - деление кольцевой бороздой с участием фикопласта; 2 - деление с помощью клеточной пластинки, образующейся по типу фикопласта); 3, 4 - веретено деления сохраняется, ядра отдалены друг от друга (3 - деление кольцевой бороздой; 4 - деление с помощью клеточной пластинки, образующейся по типу фрагмопласта)
Размножаются зеленые водоросли половым и бесполым путем, с помощью вегетативных и специализированных клеток. Бесполое (вегетативное) размножение у одноклеточных, лишенных клеточной оболочки, происходит делением клетки надвое (например, Pedinomonas, Dunaliella), у колониальных (неценобиальных) и многоклеточных - фрагментацией таллома (Raciborskiella Wisl., Dictyosphaerium Näg., Botryococcus, Chlorhormidium; рис. 19.7). Ценобиальные (Volvox, Hydrodictyon, Scenedesmus Meyen) образуют дочерные ценобии внутри клеток материнского ценобия (рис. 19.8). Многие размножаются с помощью акинет (Pithophora Wittr., Chlorhormidium; см. рис. 19.7). У Charophyceae из узловых клеток таллома могут возникать новые растения, отчленяющиеся от материнского, наблюдаются также специальные органы вегетативного размножения - одно- и многоклеточные корневые и стеблевые клубеньки.
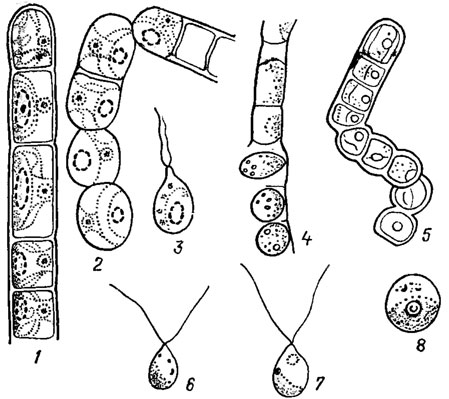
Рис. 19.7. Различные способы размножения видов рода Chlorhormidium Fott: 1 - участок вегетативной нити; 2 - фрагментация нити; 3 - зооспора; 4 - образование апланоспор; 5 - образование акинет; 6 - микрогамета; 7 - макрогамета; 8 - зигота
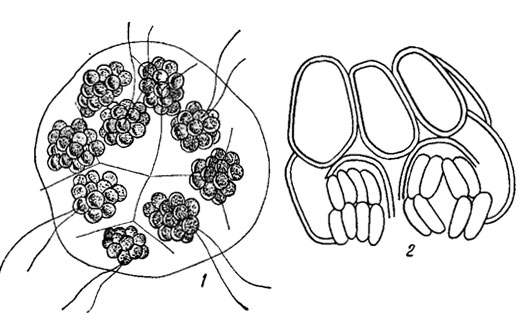
Рис. 19.8. Бесполое размножение ценобиальных зеленых водорослей: 1 - Voivulina steinii Playf.; 2 - Scenedesmus obfusus Meyen f. aiternans (Reinsch) Comp
Бесполое размножение с помощью специализированных клеток - спор - широко представлено у многих зеленых водорослей, нередко наряду с вегетативным и половым размножением (см. рис. 19.7). Иногда образование спор является единственным способом размножения (Chlorella), у некоторых споры отсутствуют (Charа). Споры зеленых водорослей бывают подвижными (зооспоры) и неподвижными (апланоспоры) (рис. 19.9; см. рис. 19.7). Наблюдается также образование автоспор, гипноспор, гемизооспор и гемиавтоспор. Споры бывают шаровидными, яйцевидными, эллипсоидными, с клеточной оболочкой или без нее. Зооспоры обычно имеют 2-4 изоморфных (акронематических), изоконтных, изодинамических жгутика. У Oedogoniales и некоторых Bryopsidales зооспоры (андроспоры) с многочисленными жгутиками, расположенными венцом у переднего конца клетки (см. рис. 19.9, 10). Наблюдаются и другие отклонения от обычного строения жгутикового аппарата; обнаружены разные типы тонкой структуры жгутиковой корневой системы. Считают, что различия в строении спор (а также гамет) в известной мере отражают различия в строении предков соответствующих таксонов. Спорангии одноклеточные, обычно не отличающиеся от вегетативных клеток, но иногда они образуются как выросты последних, выполняя только функцию продуцирования спор. Специализированные спорангии наблюдаются у представителей семейства Trentepohliacea (см. рис. 19.9). Количество спор в спорангиях колеблется от одной (Oedogonium) до нескольких сотен (Cladophora).

Рис. 19.9. Спорангии и споры зеленых водорослей: 1-3 - Trentepohiia annuiata Brand (1 - участок нити с субспорангиальной клеткой и зрелым спорангием на вершине; 2, 3 - зооспоры); 4, 5 - Chiorococcum sp. (4 - спорангий, 5 - зооспора); 6 - Chloreiia sp. (освобождение автоспор из спорангия); 7 - Tetraedron minimum (A. Br.) Hansg. (освобождение автоспор); 8, 9 - Uiothrix zonata (Web. et Moh.) Kütz. (8 - спорангий, 9 - зооспора); 10 - Oedogonium concatenatum (Hass ) Wittr. (выход зооспоры из спорангия); 11 - Microspore willeana Lagerh. (освобождение апланоспор); 12 - Ulothrix sp. (образование гипноспор)
Половой процесс представлен разнообразными формами: гологамией (Dunaliella, Asteromonas Artary), изогамией (Chlamydomonas moewusii Gerloff), гетерогамией (Eudorina unicocca G. Sm.) оогамией (Volvox), конъюгацией (Conjugatophyceae). Наблюдаются формы полового процесса, промежуточные между изо- и гетерогамией (Draparnaldiella Meyer et Skabitsch.), гетеро- и оогамией (Aphanochaete А. Вr.). Наличие различных форм полового процесса (изо-, гетеро- и оогамии) в пределах одного рода Chlamydomonas свидетельствует о том, что эволюция его происходила независимо от морфологической эволюции. В строении гамет, особенно их жгутикового аппарата, наблюдается разнообразие, отмеченное для спор и монадных форм. Половые органы одноклеточные, морфологически не отличающиеся (Cladophora) или отличающиеся (Coleochaete) от вегетативных клеток. Лишь у видов рода Draparnaldiella отмечены многокамерные гаметангии [217]. Образования, рассматриваемые ранее как многоклеточные половые органы Charophyceae, трактуются ныне как сильно редуцированные гаметофиты, развивающиеся из прорастающих in situ (в микро- и макроспорангиях - инициальных клетках антеридиев и оогониев) спор [253, 623]. Среди зеленых водорослей есть гомо- и гетероталличные, одно- и двудомные виды. Изредка наблюдается половой диморфизм (наннандриевые виды Oedogoniales). Некоторым свойственны партеногенез (Chara spp.) и вторичная утрата полового процесса (многие Chlorococcales). Половой процесс не всегда сопровождается размножением (одноклеточные Desmidiales). У некоторых половое размножение - единственная форма репродукции (Draparnaldiella). Обычно в цикле развития зеленых водорослей сочетаются различные формы размножения, нередко наблюдается поливариантность индивидуального развития.
Зиготы, образующиеся при голо-, изо- и гетерогамии, обычно проходят стадии планозиготы и гипнозиготы (Chlamydomonas, Ulothrix). При конъюгации и оогамии образуются неподвижные зиготы, которые обычно проходят стадию гипнозиготы (ооспоры, зигоспоры, например, у Oedogonium, Spirogyra, Chara). Иногда, минуя покоящееся состояние, зиготы непосредственно прорастают в новые растения (многие Bryopsidales).
Циклы развития зеленых водорослей очень разнообразны (см. раздел 3.2.2). Наряду с цикломорфозом, характерным для агамных и апогамных видов, у эугамных наблюдается зиготический (Spirogyra, Oedogonium), спорический (Ulva), гаметический (Codium Stack.) и соматический (Prasiola Ag., Coleochaete) мейоз. Смена форм развития изо- (Ulva, Enteromorpha Link) или гетероморфная (Bryopsis, Monostroma Thur.), с преобладанием гаметофита (Bryopsis Lamour.), гаметоспорофита (Monostroma, Urospora Aresch.) или спорофита (Halicystis Aresch.). В цикле развития харовых различают крупный спорофит и развивающиеся на нем сильно редуцированные карликовые мужской и женский гаметофиты [253]. Иногда зиготы осуществляют фотоассимиляцию и значительно увеличиваются в размерах в процессе созревания (Chlamydomonas); у них наблюдается смена зигоспорофита и гаметофита [45, 253]. В цикле развития Coleochaete происходит смена гаметоспорофита и развивающихся на нем диплоидных спорофитов, в которых происходит соматический мейоз [253].
Циклы развития могут быть однотипными у всех представителей крупного таксона (Charophyceae) или различаться у видов одного рода (Ulothrix), в различных популяциях одного вида и у различных особей одной популяции в зависимости от условий произрастания (Bryopsis, Protosiphon Klebs). Считают, что исходным в эволюции эугамных зеленых водорослей был неизвестный у ныне существующих цикл развития с нефиксированным положением мейоза. В дальнейшем процессе эволюции в различных группах одновременно могли закрепиться зиготическая, спорическая и соматическая редукция. Наиболее широкое распространение получил спорический мейоз с дифференциацией форм развития на изо- и гетероморфные. Приобретение талломами, принадлежащими к различным формам развития, разных размеров и строения имело большое адаптивное значение. Гаметическая редукция, связанная с более совершенной формой полового процесса - гаметогамией (мерогамией), имеет вторичное происхождение. Она не получила широкого распространения. Вершиной в эволюции циклов развития зеленых водорослей явилось возникновение организмов двойственной природы - дибионтов (Coleochaete, Charophyceae).
По составу фотосинтетических пигментов зеленые водоросли близки к Euglenophyta, Prochlorophyta, Chlorarachniophyta и высшим растениям. Только у представителей этих групп содержатся хлорофиллы а и b. На основании сходства пигментного состава, окраски таллома и общего габитуса Euglenophyta и некоторых зеленых водорослей (Volvocales) их иногда объединяют в отделе Chlorophyta [374 , 682]. Однако принципиальные различия между зелеными водорослями и Euglenophyta в тонкой структуре клетки, особенно в строении клеточных покровов, митотических ядер, стигм, в химизме клеточных покровов, основных продуктов ассимиляции и локализации последних свидетельствуют об отсутствии прямых родственных связей между этими таксонами [120]. Открытие Prochlorophyta послужило основанием для гипотезы об эндосимбиотическом происхождении хлоропластов зеленых растений от древних прокариотических зеленых водорослей [369, 612]. Единственный представитель Chlorarachniophyta Hibberd et Norris - Chlorarachnion reptans Geitl. - характеризуется необычным сочетанием уникальных ультраструктурных признаков и наличием хлорофиллов а и b; его происхождение предположительно рассматривают как результат симбиоза бесцветного амебоида и зеленого эукариотического организма, однако родственные связи эндосимбионта остаются загадочными [478]. Сходство зеленых водорослей и высших растений, проявляющееся не только в составе фотосинтетических пигментов, но и в химизме клеточной оболочки, локализации основных продуктов ассимиляции, в строении вегетативных и репродуктивных клеток, тонкой структуре жгутикового аппарата и хлоропластов, наиболее фундаментально. Оно свидетельствует в пользу тесных родственных связей зеленых водорослей и высших растений вследствие происхождения их от общих предков [120, 414, 415, 417, 423, 425, 500, 643, 645].
Рассматривая зеленые водоросли и высшие растения как единый филогенетический ствол зеленых растений, Д. К. Зеров объединил их в отделе Chlorophyta sensu lato, в котором зеленые водоросли занимают исходное положение в качестве подотдела Chlorophycophytina. Если же по традиции рассматривать зеленые водоросли в качестве отдела, то, по мнению Д. К. Зерова, используя номенклатуру, предложенную Г. Ф. Папенфуссом [570], этот отдел следует называть Chlorophycophyta.
История развития взглядов на пути эволюции и классификацию зеленых водорослей насчитывает почти столетие [120]. В середине XX ст. наиболее популярной была система, включающая три класса: Chlorophyceae (с порядками Volvocales, Tetrasporales, Chlorococcales, Ulotrichales, Bryopsidales, Siphonocladales), Conjugatophyceae (с порядками Mesotaeniales, Zygnemales, Gonatozygales, Desmidiales) и Charophyceae (с одним порядком Charales) [414, 417 и др.]. Она отражала взгляды на эволюцию зеленых водорослей от жгутиковых предков в направлении развития неподвижного многоклеточного таллома растительного типа. Считалось, что основные этапы эволюции зеленых водорослей представляют порядки собственно зеленых водорослей (Chlorophyceae), а Conjugatophyceae и Charophyceae рассматривали как боковые крайне специализированные ветви, рано отделившиеся и уклонившиеся от магистрального пути эволюции зеленых растений.
Исследования последних десятилетий, проведенные с применением электронно-микроскопических и биохимических методов [420, 501, 588, 589, 500, 607, 642, 660-662 и др.], послужили основой для попыток реклассификации зеленых водорослей. К. Д. Стюарт и К. Р. Мэттокс [547, 660, 661] считают, что предками зеленых водорослей были покрытые чешуйками жгутиковые, близкие современным Prasinophyceae. Этот класс, согласно М. Шадефо [371], характеризуется наличием жгутиковой ямки у монадных форм и стадий. Электронно-микроскопические и биохимические исследования обнаружили у них мелкие чешуйки, покрывающие тело монады, а иногда и несущие волоски жгутики, своеобразное строение переходной зоны жгутиков, базальных тел и жгутиковых корней, необычный состав фотоассимиляционных пигментов [202, 559, 560]. Класс Prasinophyceae принимается многими авторами [359, 362, 373, 405, 407, 486, 487, 632 и др.] и нередко рассматривается как исходный в эволюции зеленых водорослей. Отмечается также возможное родство Prasinophyceae с хромофитной линией эволюции водорослей [371, 372, 560]. Ф. Раунд [611] возводит их в ранг самостоятельного отдела Prasinophyta.
Т. Христенсен [374], кроме Prasinophyceae, выделил еще класс Loxophyceae, куда он отнес монадные организмы, не обладающие радиальной симметрией (например, Pedinomonas). Однако выделение этого класса не встретило широкой поддержки.
К. Стюарт и К. Мэттокс [660, 661] на основании тонкого строения монадных форм и стадий, в первую очередь жгутикового аппарата (см. рис. 19.4), особенностей митоза и цитокинеза (см. рис. 19.6) разделили зеленые водоросли на три класса: Chlorophyceae, Ulvophyceae и Charophyceae. Первый объединяет Volvocales, Tetrasporales, Chlorococcales, Chtorosarcinales, Ulotrichales (частично), Chaetophorales и Sphaeropleales, у которых деление клеток происходит с помощью кольцевой борозды или клеточной пластинки, образующейся по типу фикопласта, жгутиковая корневая система крестообразная, субмикроскопические чешуйки и ризопласт отсутствуют. Ulvophyceae включает Ulotrichales (частично), Trentepohliales, Ulvales, Cladophorales, Acrosiphonales, Caulerpales, Siphonocladales, Dasycladales, у которых деление клеток происходит с помощью кольцевой борозды, жгутиковая корневая система крестообразная, есть ризопласт и образуются субмикроскопические чешуйки. Charophyceae, в понимании К. Стюарта и К. Мэттокса, включает Ulotrichales (частично), Klebsormidiales, Coleochaetales, Charales. Деление клеток у них происходит с помощью кольцевой борозды с сохранением веретена деления или с помощью клеточной пластинки, образующейся по типу фрагмопласта, жгутиковая корневая система унилатеральная с многослойной структурой (см. рис. 19.4), ризопласт отсутствует, монадные клетки обычно покрыты субмикроскопическими чешуйками. Charophyceae sensu Stewart et Mattox, соответствующий Phragmophyceae sensu Chadefaud [372], по указанным признакам наиболее близок высшим растениям. Эту группу рассматривают как прогрессивную линию эволюции зеленых водорослей, а иногда относят к высшим растениям [641].
В дальнейшем К. Мэттокс и К. Стюарт [547] несколько пересмотрели критерии классификации зеленых водорослей, придавая меньше значения наличию или отсутствию субмикроскопических чешуек у монадных форм и стадий и больше значения - ориентации базальных тел и жгутиковых корней (рис. 19.10), поведению митотического веретена деления в течение цитокинеза, привлекая дополнительные данные об особенностях размножения, циклах развития, экологии и др. В новой системе [547], кроме выделенных ранее трех классов (Chlorophyceae, Ulvophyceae, Charophyceae), дополнительно предложено еще два: Micromonadophyceae и Pleurastrophyсеае. Первый объединяет исключительно монадные голые или покрытые чешуйками микроорганизмы с интразональным митотическим веретеном деления, сохраняющимся во время цитокинеза. Ко второму отнесены монадные, коккоидные, сарциноидные и нитчатые организмы с крестообразной микротрубчатой жгутиковой корневой системой и ризопластом, метацентрическим митотическим веретеном деления, коллапсирующим в телофазе, и цитокинезом с помощью фикопласта особого типа. В новой системе определено место Oedogoniales среди Chlorophyceae и Zygnemales в пределах Charophyceae [547].
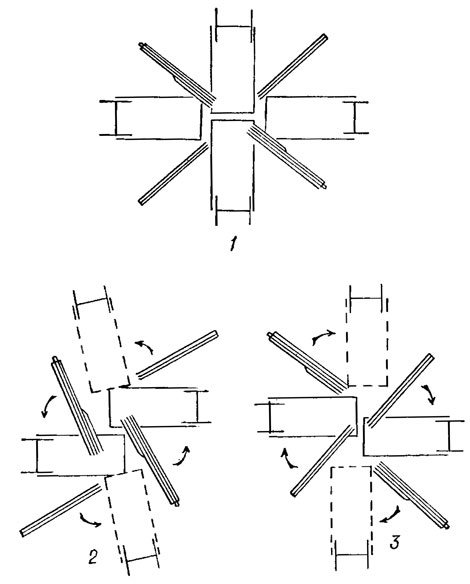
Рис. 19.10. Ориентация базальных тел и жгутиковых корней у некоторых зеленых водорослей: 1 - гипотетический жгутиковый аппарат со строго крестообразным расположением базальных тел и микротрубчатых корней; 2 - сдвиг компонентов жгутикового аппарата против часовой стрелки, характерный для Ulvophyceae; 3 - сдвиг компонентов жгутикового аппарата по часовой стрелке, характерный для Chlorophyceae sensu str
Предлагались и другие критерии для реклассификации зеленых водорослей на уровне классов: стефаноконтный тип монадных стадий и особый тип вегетативного клеточного деления с образованием колпачков (Oedogoniophyceae [610]), многоядерность, в результате разобщенности процессов митоза и цитокинеза (Bryopsidophyceae [610]), особенности циклов развития, в частности наличие особой стадии Codiolum (Codiolophyceae [512]), наличие клеточных оболочек у монадных форм и стадий, хламидомонадный тип строения репродуктивных клеток (Chlamydophyceae [406]), особенности экологии и циклов развития в комплексе с некоторыми другими признаками (Siphonophyceae [50 и др.)]. Ряд авторов принимает деление отдела зеленых водорослей на пять-шесть классов, соответствующих порядкам в общепринятом понимании* [113, 239], другие отказываются от выделения классов, разделяя отдел на многочисленные порядки [356]. Рассматривая зеленые водоросли как гетерогенную группу, возникшую в результате нескольких актов эндосимбиоза, в которых принимало участие несколько различных жгутиковых предков и один (или несколько?) фотосинтезирующий симбионт - предок хлоропластов современных зеленых растений, Т. Кавалер-Смит [368, 369] и Ф. Раунд [612] предлагают объединить их с высшими растениями в царство Viridiplantae, а в пределах последнего выделить ряд самостоятельных отделов: Prasinophyta, Chlorophyta, Ulvophyta, Charophyta, Bryophyta, Tracheophyta.
* (Основной критерий выделения порядков в альгологии, по мнению большинства авторов, - тип структуры вегетативного тела водорослей.)
Поскольку с точки зрения новых критериев большинство зеленых водорослей остается все еще слабо изученным, в настоящем издании принимается традиционная система отдела зеленых водорослей [417] с добавлением к имеющимся в ней трем классам (Chlorophyceae, Conjugatophyceae, Charophyceae*), четвертого - Siphonophyceae, выделение которого можно считать достаточно обоснованным [50].
* (Следуя М. М. Голлербаху [79, 81], автор раздела, посвященного харовым водорослям, рассматривает их в качестве самостоятельного отдела Charophyta (см. раздел 20).)
Зеленые водоросли широко распространены в воде, почве, в наземных местообитаниях, на разных географических широтах, во всех морях и на всех континентах земного шара. Они встречаются в водах с различной степенью солености (от пресных до пересоленных), трофности (от дистрофных до эвтрофных), с различным содержанием органических веществ (от ксено- до полисапробных), водородных ионов (от кислых до щелочных), при различных температурах (термо-, мезо- и криофильные виды). Среди них есть планктонные, перифитонные и бентосные организмы. Многие обитают в почве, на ее поверхности, на скалах, коре деревьев, стенах домов, в других вневодных местообитаниях, включая толщу воздуха (аэропланктон).
Разнообразны формы взаимоотношений зеленых водорослей с другими организмами - бактериями, грибами, высшими растениями, животными и другими водорослями. Они поселяются на других организмах в качестве эпи- и эндобионтое (эпифитов и эпизоитов, эндофитов и эндозоитов). Вступал в симбиотические отношения с грибами, зеленые водоросли входят в состав многих лишайников. В качестве эндосимбионтов зеленые водоросли существуют в клетках простейших, губок, гидр (зоохлореллы). Некоторые развиваются на шерсти млекопитающих. Случаи паразитизма зеленых водорослей на других организмах редки (Cephaleuros Kunze на различных органах тропических и субтропических растений). Зеленые водоросли могут служить хозяевами для поселяющихся на или внутри них эпи- и эндобионтов, паразитических и эндосимбиотических организмов, в том числе внутриклеточных (например, цианеллы в клетках Glaucocystis nostochinearum Itz.).
Зеленые водоросли входят в состав различных водных и вневодных фитоценозов, биоценозов, экосистем. Развиваясь иногда в массовом количестве, зеленые водоросли выходят на доминирующее положение в биоценозах. Массовое развитие микроскопических зеленых водорослей нередко вызывает зеленое или красное цветение воды, рапы, почвы, снега, скал, коры деревьев, стен домов, что иногда сопровождается образованием сапропелей (например, при массовом развитии Botryococcus braunii Kütz. в озерах).
Ближайшие родственники высших растений, берущие начало от общих с ними предков, зеленые водоросли - уникальный модельный объект эволюционной биологии, а также разнообразных генетических, физиологических, биохимических, цитологических и других исследований. Ряд видов имеет хозяйственное значение как индикаторные организмы в системе экологического мониторинга, агенты самоочищения загрязненных и доочистки сточных вод, первичное звено трофической цепи гидробионтов в рыбохозяйственных водоемах. Благодаря высокому КПД утилизации солнечной энергии, некоторые зеленые водоросли - перспективный объект биотехнологии для получения биогаза, продуктов питания и кормов, ценных витаминных, ферментных, фармацевтических препаратов. Рассматривается возможность использования зеленых водорослей в качестве фотосинтетического звена в системе жизнеобеспечения космонавтов (см. раздел 6), объекта промышленного фотосинтеза.
Более детальные сведения о зеленых водорослях можно почерпнуть в монографиях и других изданиях [42, 44, 48, 51, 113, 122, 239, 359, 362, 487, 552, 559, 560, 588, 625, 670] (см. также ссылки в описаниях классов и порядков зеленых водорослей).
https://lesnojgorodoksm.date предлагает анонимность и конфиденциальность для вашего удобства.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'