15. Отдел 6. Диатомовые водоросли - Bacillariophyta (Diatomeae) (С. П. Вассер, Л. Н. Бухтиярова)
Диатомовые водоросли представлены одноклеточными и колониальными микроскопическими индивидами светло-желтого или бурого цвета, имеющими исключительно коккоидный тип структуры тела. Жесткая оболочка диатомовых состоит из прозрачного, как правило, симметричного кремнеземного панциря. Размер клеток от 4 до 2000 мкм. В отделе, по данным разных авторов, насчитывается 12-25 тыс. видов.
Структура панциря, его форма, соотношение осей и плоскостей симметрии лежат в основе систематики диатомовых водорослей. Панцирь состоит из аморфного кремнезема, напоминающего по составу опал (Si2 × xН2O, плотность 2,07), с примесью металлов (алюминия, железа, магния) и органического компонента, возможно, белка. Толщина стенок панциря зависит от концентрации кремния в среде и изменяется в значительных пределах: у тонкостенных форм - от сотых до десятых долей микрометра, у толстостенных - 1-3 мкм. Панцирь состоит из двух частей - эпитеки и гипотеки. Большая часть - эпитека надвигается своими краями на гипотеку как крышка на коробку. Эпитека состоит из плоской или выпуклой створки - эпивальвы и поискового ободка - эпицингулюма (рис. 15.1; 15.2). Гипотека имеет аналогичные части: створку - гиповальву и поисковый ободок - гипоцингулюм. Эпицингулюм и гипоцингулюм составляют вместе поясок панциря. Поясок панциря отсутствует у некоторых ископаемых видов, спор и иногда у инициальных клеток. Загнутая часть створки - загиб створки, бывает низким или высоким, иногда он имеет характерную структуру. Поисковый ободок в виде узкой ленты плотно охватывает край загиба створки, но не срастается с ним. Элементы поискового ободка, которые располагаются проксимально к створке, называют вставочными ободками (копулами), иногда они отличаются формой и структурой от дистальных элементов. Вставочные ободки бывают воротничковидные, кольцевидные, полукольцевидные или состоят из отдельных сегментов разнообразной формы (рис. 15.3). Вставочный ободок, непосредственно примыкающий к створке, часто называют вальвокопулой. Форма вставочных ободков - один из характерных признаков некоторых родов (Mastogloia Thw.). Разрастание вставочного ободка в полость клетки приводит к образованию неполной кремниевой перегородки - септы (рис. 15.4), параллельной створкам. Вставочные ободки обеспечивают увеличение объема клетки и ее рост за счет увеличения их числа или их раздвигания. Дистальные к створке элементы поискового ободка называются соединительными ободками, или плеврами. Поисковый ободок, состоящий из одного элемента, - также соединительный. В том случае, если загиб створки в виде неширокой закраины углубляется внутрь панциря, образуется диафрагма, которая в отличие от септ, является частью створки.
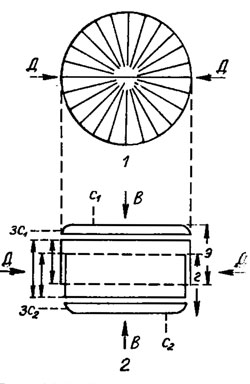
Рис. 15.1. Схема строения панциря диатомовых водорослей центрического типа (род Coscinodiscus): 1 - вид со стороны эпитеки; 2 - вид со стороны пояска; ДД - диаметр панциря; ВВ - центральная ось панциря; с1 - створка эпитеки; э - эпитека; г - гипотека; с2 - створка гипотеки; зс2 - загиб створки гипотеки; зс1 - загиб створки эпитеки
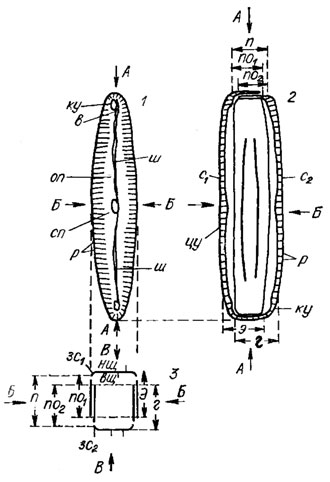
Рис. 15.2. Схема строения панциря диатомовых водорослей пеннатного типа (род Pinnularia): 1 - вид со стороны эпитеки; 2 - вид со стороны пояска; 3 - поперечное сечение панциря; АА - продольная ось панциря; ББ - поперечная ось панциря; ВВ - вертикальная первальварная ось панциря; ку - конечный (полярный) узелок; в - воронка; оп - осевое поле; сп - среднее поле; р - ребра; ш - шов; п - поясок; по1 - поисковый ободок эпитеки; по2 - поисковый ободок гипотеки; с1 - створка эпитеки; с2 - створка гипотеки; цу - центральный узелок; зсх - загиб створки эпитеки; нщ - наружная щель шва; вщ - внутренняя щель шва; э - эпитека; г - гипотека; зс2 - загиб створки гипотеки
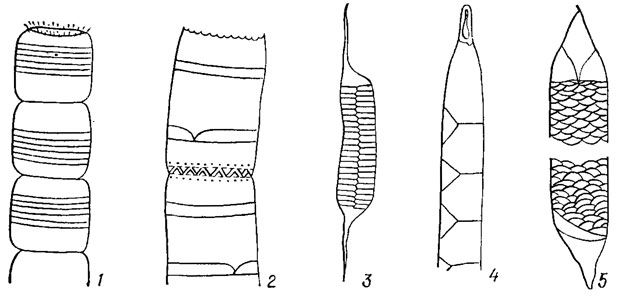
Рис. 15.3. Форма вставочных ободков панцирей диатомовых водорослей: 1 - кольцевидная (Lauderia); 2 - воротничковидная (Stephanodiscus); 3 - трапециевидно-чешуевидная; 4 - полукольцевидная; 5 - чешуевидная (3-5 Rhizosolenia)
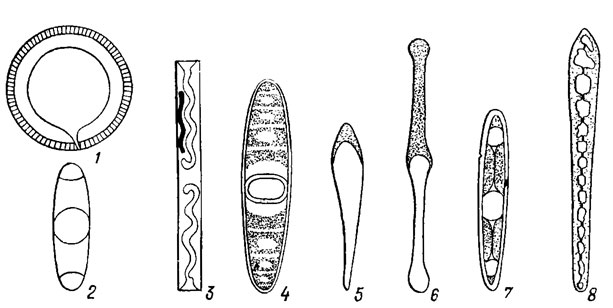
Рис. 15.4. Форма и расположение септ у диатомовых водорослей: 1-3 - вставочный ободок с септой (1 - центрическая диатомея Thalassiosira antigua (Grun.) А. Cl. var. septata Pr.-Lavr., 2, 3 - пеннатные диатомеи Grammatophora marina (Lyngb.) Kütz. и G. serpentina (Raifs) Ehr.); 4 - центральная септа; 5 - полярная септа (Licmophora ehrenbergii (Kütz.) Grun.); 6 - септа, занимающая половину панциря (Tabellaria fenestrate (Lyngb.) Kütz.); 7 - септа с тремя отверстиями (Rhabdonema adriaticum Kütz.); 8 - септа с многочисленными отверстиями (Climacosphenia moniligera)
Форма панциря разнообразна: в виде шара, диска, цилиндра и т. п. Она определяется формой створок (рис. 15.5) и их высотой. Благодаря симметричности строения панциря через него можно провести продольную, поперечную и центральную (первальварную) оси симметрии, длина которых соответственно определяет длину, ширину и высоту панциря, а также продольную (апикальную), поперечную (трансапикальную), створковую (вальварную) плоскости симметрии (рис. 15.6). Если через панцирь можно провести все три плоскости симметрии, то он полностью симметричный, две - бисимметричный, одну - моносимметричный. Встречаются асимметричные панцири, через которые нельзя провести ни одной плоскости симметрии.
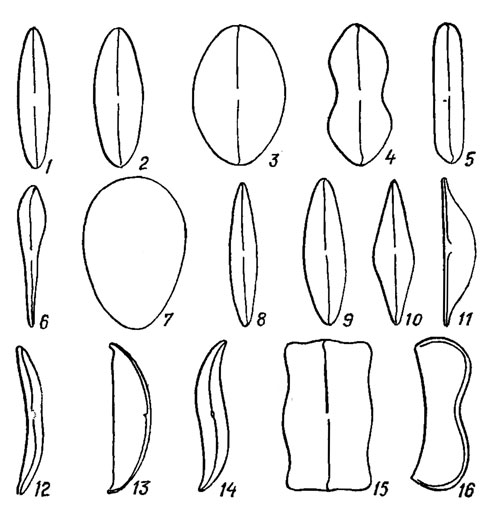
Рис. 15.5. Форма створок панциря диатомовых водорослей: 1, 2 - эллиптическая; 3 - широкоэллиптическая; 4 - гитаровидная; 5 - линейная; 6 - булавовидная; 7 - яйцевидная; 8, 9 - ланцетная; 10 - ромбическая; 11, 13 - полуланцетная; 12 - полудугообразная; 14 - сигмоидная: 15 - прямоугольная; 16 - почковидная
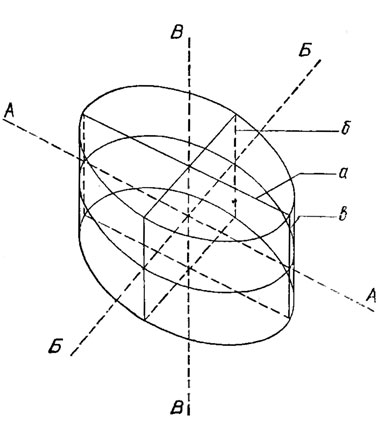
Рис. 15.6. Оси и плоскости симметрии панциря диатомовых водорослей. Оси симметрии: АА - продольная, ББ - поперечная, ВВ - центральная; плоскости симметрии; а - продольная, б - поперечная, в - створковая
Различают два основных типа створок: актиноморфные (круглые, треугольные, многоугольные), через которые можно провести три и более плоскостей симметрии (Centrophyceae) и зигоморфные, продолговатые с бисимметричной (перистой) структурой, через которое можно провести не более двух плоскостей симметрии (Pennatophyceae). Среди зигоморфных преобладают створки с одинаковыми концами - изопольные (см. рис. 15.5, 1-5, 8-16), реже - с разными - гетеропольные (см. рис. 15.5, 6, 7). Концы створок также имеют разнообразную форму (рис. 15.7).
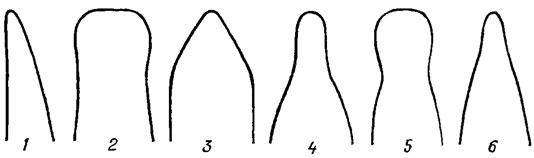
Рис. 15.7. Концы створок панциря диатомовых водорослей: 1 - острый; 2 - тупой; 3 - клиновидный; 4 - клювовидный; 5 - головчатый; 6 - оттянутый
Наружный и внутренний рисунки панциря, наблюдаемые в световой и электронный микроскоп, называют структурой панциря. Она специфична для разных таксонов и образована различными структурными элементами, из которых всеобщими и наиболее важными являются перфорации - система отверстий различного строения расположенных на створках, через которую происходит связь протопласта с внешней средой.
Наиболее распространенные из перфораций ареолы, бывают двух типов: пороидные (пороиды) (рис. 15.8, 1, 2), не имеющие заметных сужений к поверхности створки, и локулярные (локулусы), заметно суженные к одной из поверхностей створки (рис. 15.8, 3, 4). С внутренней или внешней стороны, иногда с обеих, ареолу закрывает тонкий перфорированный кремнеземный слон - велум. Положение велума имеет важное значение в таксономии крупных рангов класса Centrophyceae. Противоположное незамкнутое отверстие ареолы называется фораменом [230].
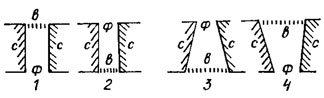
Рис. 15.8. Схема строения ареол панциря диатомовых водорослей: 1, 2 - пороидные; 3, 4 - локулярные (1,4 - внешнее положение велума; 2, 3 - внутреннее положение велума); ф - форамен; в - велум; с - стенки панциря
Удлиненная камера или одна из серии удлиненных камер, проходящих от осевой части створки к краю и открывающихся внутрь панциря большим отверстием с ареолированным наружным слоем, называется альвеолой. При наблюдении в световой микроскоп перфорации имеют вид точек, либо при плотном расположении в ряд - штрихов. У центрических диатомовых перфорации образуют радиальные штрихи, проходящие от центра к краю створки, пучковые, сгруппированные в параллельные радиальному штриху ряды, и тангенциальные, образующие прямые или изогнутые не радиальные ряды (рис. 15.9). У пеннатных диатомовых перфорации образуют относительно края створки параллельные, радиально-симметричные и конвергентные штрихи (рис. 15.10). Иногда штрихи взаимно пересекаются.
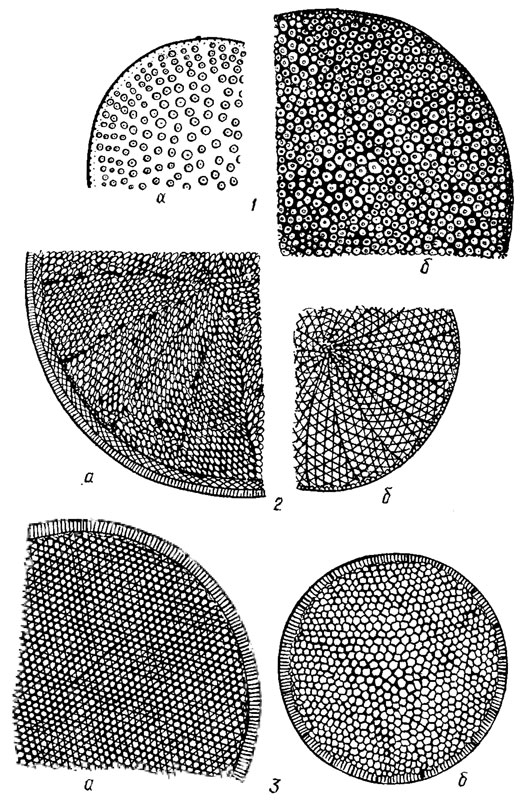
Рис. 15.9. Перфорационные штрихи центрических диатомовых водорослей: 1 - радиальные (а - отдельные ареолы, б - ареолы с вторичными спиральными рядами); 2 - пучковые (а - прямые пучки, б - изогнутые пучки); 3 - тангенциальные (а - прямые ряды - линейный тип, б - вогнутые по направлению к краю ряды - эксцентрический тип)
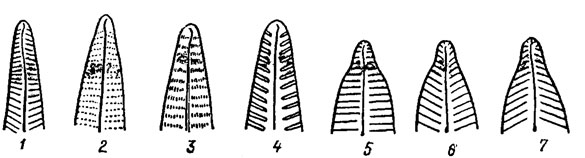
Рис. 15.10. Перфорационные штрихи пеннатных диатомовых водорослей: 1 - гладкие; 2 - пунктирные; 3 - линеолированные; 4 - ребровидные; 5 - параллельные; 6 - радиально-симметричные; 7 - конвергентные
На створках панциря большинства пеннатных диатомей имеется шов в виде пары сквозных щелей, каждая из которых называется ветвью шва. Швы имеют различную длину, разнообразное строение и могут располагаться на обеих створках или на одной из них. Хорошо развитый щелевидный шов характерен для представителей семейства Naviculaceae. Он состоит из пары коленчато согнутых в толще створки щелей, заметно суженных в колене и расширяющихся наружу (наружная щель) и внутрь (внутренняя щель) панциря (рис. 15.11; 15.12). На середине створки ветви шва соединяются в центральном узелке (внутреннее утолщение стенки створки) горизонтальным и вертикальным воронковидным каналами, причем наружный выход последнего имеет вид точки - центральная пора. В конечных (полярных) узелках - внутренних утолщениях стенки створки или обычно губообразно выпяченных образованиях - хоботках (см. рис. 15.11; 15.12) - наружная щель заканчивается извитой, обычно короткой полярной щелью.
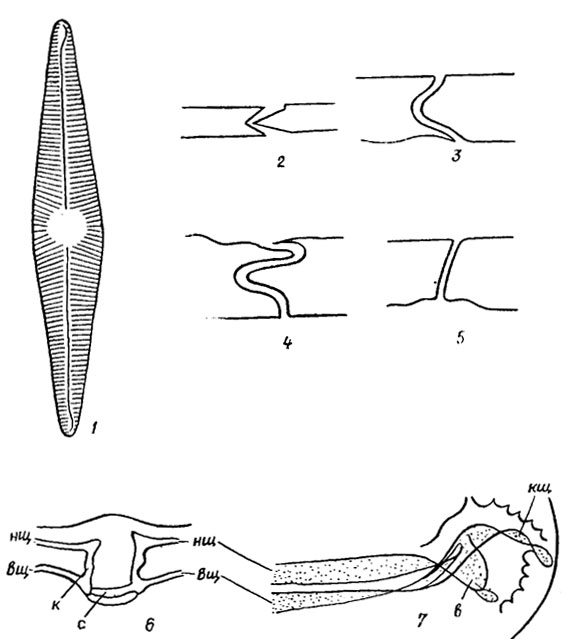
Рис. 15.11. Строение щелевидного шва створок панциря диатомовых водорослей: 1 - створка со щелевидным швом; 2-5 - щели шва в поперечном разрезе; 6 - продольный срез через центральный узел; 7 - шов в районе конечного узелка (вид сверху); нщ - наружная щель; вщ - внутренняя щель; к - канал, соединяющий наружную и внутреннюю щели; с - канал, соединяющий две ветви шва в центральном узелке; кщ - полярная щель; в - конец внутренней щели шва
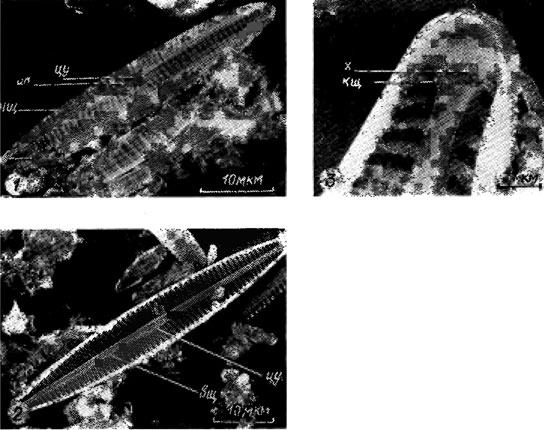
Рис. 15.12. Строение щелевидного шва створок панциря Navicula gracilis Ehr.: 1 - внешняя поверхность створки (цу - центральный узелок, цп - центральная пора, нщ - наружная щель, пщ - полярная (конечная) щель); 2 - внутренняя поверхность створки (цу - центральный узелок, вщ - внутренняя щель); 3 - внутренняя поверхность конца створки (х - хоботок, кщ - конечная щель)
Каналовидный шов представителей семейств Nitzschiaceae, Epithemiaceae, Surirellaceae и других в виде трубковидной полости проходит в толще створки, гребневидном утолщении ее - киле, или крыловидном выросте - крыле, окружающем створки по краю (рис. 15.13-15.15). Наружу каналовидный шов открывается узкой щелью, а от внутренней полости панциря отделяется рядом кремнеземных пластинок - фибул, открытые пространства между которыми называются интерфибулами.
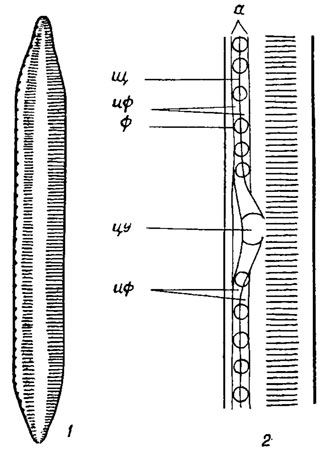
Рис. 15.13. Строение каналовидного шва у диатомовых водорослей: 1 - створка с каналовидным швом; 2 - средняя часть каналовидного шва (а - канал-шов, щ - щель каналовидного шва, ф - фибула, цу - центральный узелок, иф - интерфибулы)
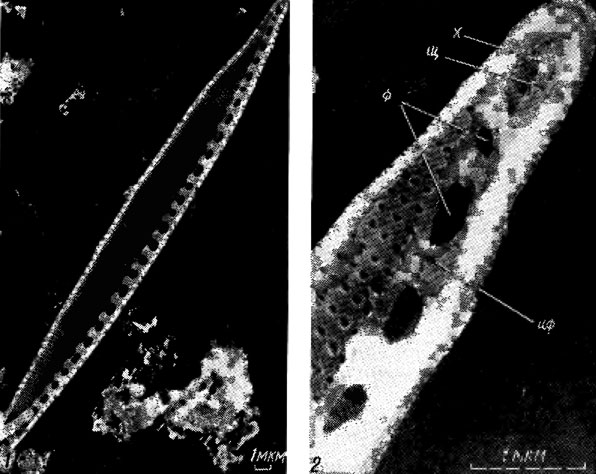
Рис. 15.14. Строение каналовидного шва у Nitzschia sp.: 1 - внутренняя поверхность створки; 2 - внутренняя поверхность конца створки (х - хоботок, щ - щель шва, ф - фибулы, иф - интерфибула)
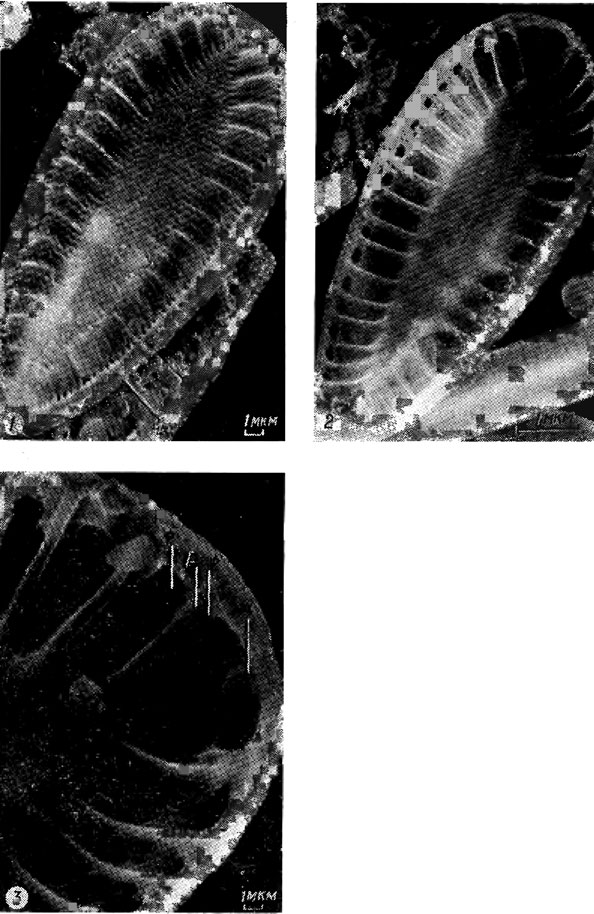
Рис. 15.15. Строение каналовидного шва у Surirella sp.:
Швы обеспечивают сообщение протопласта с внешней средой и способность к движению. В филогенетическом отношении наличие шва - прогрессивный признак, характерный для эволюционно более молодых видов.
Исследование диатомовых водорослей с помощью сканирующей электронной микроскопии позволило открыть разнообразные выросты, соединяющие протопласт с внешней средой, число и расположение которых на створке может быть различным. Большинству центрических и бесшовных диатомей свойственны двугубые выросты, конец внутренней части трубки которых сплющен, или трубка укорочена до щели, имеющей иногда валиковидно утолщенные края ("губы"); внешнее отверстие обычно округлое (рис. 15.16; 15.17). Выросты с опорами - трубки, окруженные 2-5 камерами или сопутствующими порами, изолированными изогнутыми опорами (рис. 15.18), - характерный признак только семейства Thalassiosiraceae. У некоторых видов выявлены полые наружные трубки, замкнутые с одного конца - замкнутые выросты [263, 272].
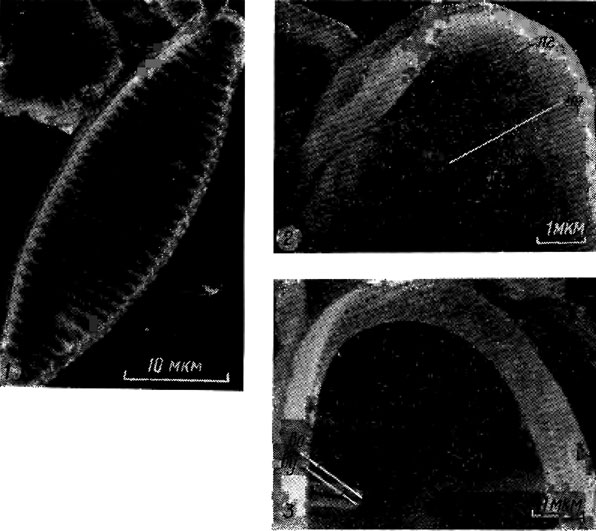
Рис. 15.16. Строение двугубого, выроста и его положение на створке Diatoma vulgare Bory: 1 - внешняя поверхность створки; 2 - внешняя поверхность конца створки (лг - ложный глазок, но - наружное отверстие); 3 - внутренняя поверхность конца створки (во - внутреннее отверстие укороченной трубки, ву - валиковидное утолщение края трубки)
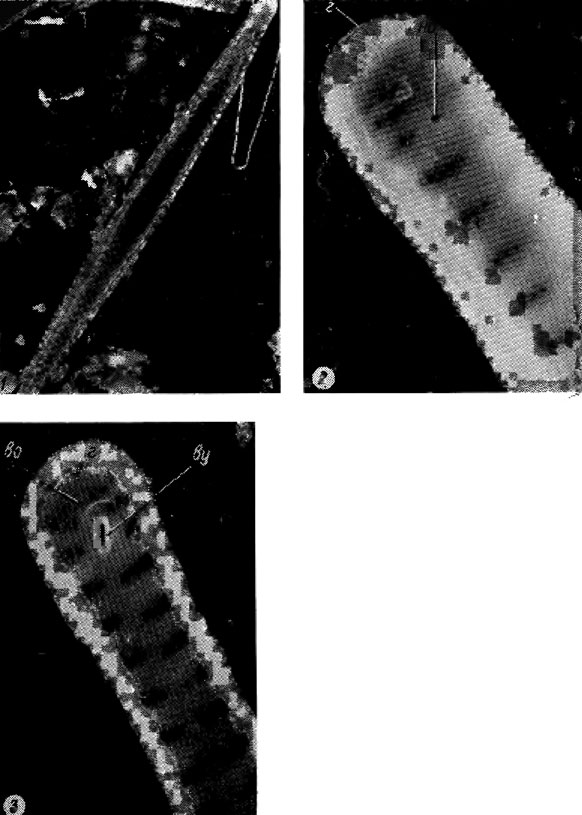
Рис. 15.17. Строение двугубого выроста у Synedra sp.: 1 - внешняя поверхность створки; 2 - внешняя поверхность конца створки (г - глазок, но - наружное отверстие); 3 - внутренняя поверхность конца створки (г - глазок, во - внутреннее отверстие укороченной трубки, ву - валиковидные утолщения края щели)
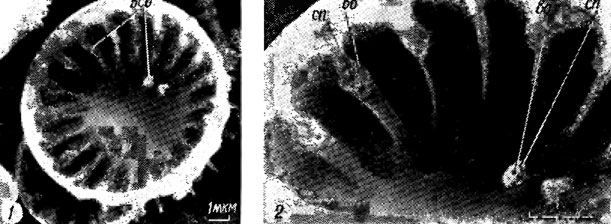
Рис. 15.18. Строение выроста с опорами и его положение на створке Cyclotella sp.: 1 - внутренняя поверхность створки (всо - выросты с опорами, дв - двугубый вырост); 2 - увеличенная часть той же створки (сп - сопутствующая пора, во - внутреннее отверстие трубки)
Гладкие, свободные от перфорации участки створки образуют гиалиновые поля: осевое - в направлении продольной оси, среднее - расширение осевого поля на его середине, центральное - в центре створки, радиальное - между радиальными штрихами, боковое - прерывающее штрихи параллельно осевому полю.
Утолщения, выступающие над наружной или внутренней поверхностью створки называются ребрами, они обеспечивают прочность панциря. У многих диатомовых водорослей на внешней поверхности панциря образуются выступы, щетинки, шипы, шипики, которые увеличивают его поверхность и служат для соединения клеток в колонию.
Большое биологическое значение имеет выделяемая диатомовыми водорослями слизь, которая не только способствует образованию колоний и прикреплению водорослей к субстрату, но и защищает их от высыхания, воздействия других неблагоприятных условий среды, способствует, вероятно, процессам деления и ауксоспорообразования. Химический состав слизи у различных видов неодинаков; в трубках колоний Berkeleya rutilans слизь состоит из соединений маннозы и ксилозы со следами рамнозы, в капсулах Navicula pelliculosa (Breb.) Hilse - из полиуронида (полимера глюкуроновой кислоты). Слизь диатомей имеет различную консистенцию - от плотной, хрящеватой, до жидкой.
Диатомовые водоросли, имеющие шов, способны к активному движению по субстрату, иногда в толще воды (Bacillaria paradoxa Gmel., NHzschia seriata Cl., N. closterium (Ehr.) W. Sm.). Относительно механизма движения выдвинут ряд гипотез. Предполагают, что движение обусловлено током цитоплазмы в щели (канале) шва, либо током воды в полости шва. Предполагают также, что в результате сокращения фибрилл из полости шва наружу выталкивается некоторый материал, как бы приклеивающий клетку к субстрату и оставляющий на нем след. Вещества, растворяющие слизь (эфедрин, изопренелин, бензодрин), обладающие наркотическими свойствами (папаверин, антропин, кофеин) нарушают и останавливают движение диатомей.
Основное условие существования диатомовых водорослей в толще воды - способность препятствовать погружению - парение. Это обеспечивается небольшим объемом протопласта и содержанием многочисленных капелек масла, наличием тонкого панциря, часто снабженного разнообразными выростами, щетинками, другими структурными элементами, увеличивающими поверхность. У некоторых крупных диатомей выявлена способность к активному удалению из клеточного сока ионов тяжелых металлов и уменьшению суммарной концентрации ионов всех компонентов в клеточном соке по сравнению с их концентрацией в морской воде.
Колонии диатомовых образуются из одной клетки в результате ряда делений и имеют свойственную виду форму (рис. 15.19). Распад колонии не приводит к гибели составляющих ее клеток - каждая из них может дать начало новой колонии. Как правило, колонии образуют виды, лишенные способности к движению: не имеющие шва, с зачаточным швом, со швом на одной из створок. В случае образования колонии подвижными диатомеями они и внутри колонии сохраняют способность к движению.
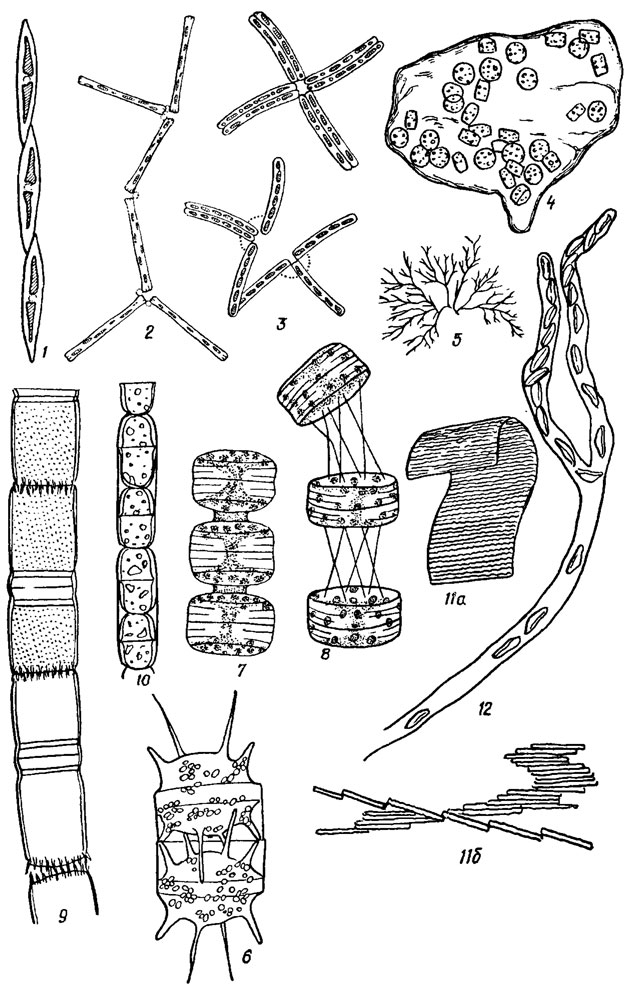
Рис. 15.19. Формы колоний подвижных диатомовых водорослей: 1, 9, 10, 16 - нитевидная (1 - Nitzschia seriata Cl., 9 - Melosira italica (Ehr.) Kütz., 10 - M. varians Ag., 16 - Leptocyl indrus danicus Cl.); 2, 3 - зигзаговидная цепочка (2 - Diatoma elongatum (Lyngb.) Ag., 3 - Thalassionema nitzschioides Grun.); 6, 7, 15 - прямая цепочка (6 - Biddulphia mobiliensis Bailey, 7 - Porosira glactalis (Grun.) Jorg., 15 - Rhizosolenia fragilissima); 8, 13 - гибкая цепочка (8 - Thalassiosira polichorda (Griin.) Pr.-Lavr., 13 - Th. exentrica (Ehr.) Cl.); 4 - студенистая бесформенная (Thalassiosira subtil is (Ostf.) Gran.); 5, 12 - студенистая ветвистая (5 - Navicula divergens, 12 - Cynibella caespitosa (Kütz.) Grun.); 11 - лентовидная (Bacillaria paradoxa Gmel.; 11a - сжатая колония, 11б - раздвинутая колония)
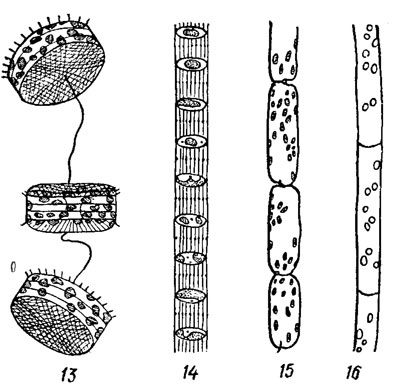
Рис. 15.19. Формы колоний подвижных диатомовых водорослей: 1, 9, 10, 16 - нитевидная (1 - Nitzschia seriata Cl., 9 - Melosira italica (Ehr.) Kütz., 10 - M. varians Ag., 16 - Leptocyl indrus danicus Cl.); 2, 3 - зигзаговидная цепочка (2 - Diatoma elongatum (Lyngb.) Ag., 3 - Thalassionema nitzschioides Grun.); 6, 7, 15 - прямая цепочка (6 - Biddulphia mobiliensis Bailey, 7 - Porosira glactalis (Grun.) Jorg., 15 - Rhizosolenia fragilissima); 8, 13 - гибкая цепочка (8 - Thalassiosira polichorda (Griin.) Pr.-Lavr., 13 - Th. exentrica (Ehr.) Cl.); 4 - студенистая бесформенная (Thalassiosira subtil is (Ostf.) Gran.); 5, 12 - студенистая ветвистая (5 - Navicula divergens, 12 - Cynibella caespitosa (Kütz.) Grun.); 11 - лентовидная (Bacillaria paradoxa Gmel.; 11a - сжатая колония, 11б - раздвинутая колония)
В слизистых колониях клетки целиком погружены в выделяемую ими гомогенную слизь и расположены беспорядочно, при этом колонии имеют вид бесформенных комочков или пленок, простых или разнообразно разветвленных трубок, обычно прикрепленных к субстрату (рис. 15.19, 4, 5, 12).
Форма колоний, не имеющих общей слизи, зависит от формы створок и способов соединения в колонию. При соединении створок всей поверхностью (соединения поясками бывают чрезвычайно редко, так как в плоскости пояска происходит деление клетки) образуются нитевидные (панцирь цилиндрический, створка округлая - Melosira Ag., Stephanodiscus Ehr.), лентовидные (панцирь плоский, створка удлиненная - Fragilaria Lyngb., Achnanthes Bory), вееровидные (панцирь клиновидный, створка булавовидная - Meiidion Ag., Licmophora Ag.) колонии. При соединении створок удлиненной формы участками, на которых расположены слизевые поры, образуются зигзаговидные (Tabellaria Ehr., Diatoma D. С.) или звездчатые (Synedra Ehr., Asterionella Hass.) колонии (см. рис. 15.19, 2, 3). Клетки в таких колониях скреплены слизью, выделяемой порами глазка или ложного глазка. Особенно распространены цепочковидные колонии, в которых смежные клетки соединены слизистыми подушечками или тяжами, короткими или длинными трубковидными выростами, рогами, щетинками и другими способами (см. рис. 15.19, 6-8, 13). Рыхлое расположение клеток в цепочковидной колонии на некотором расстоянии обеспечивает лучшие условия парения и фотосинтеза.
В результате обильного выделения слизи на базальном конце клетки образуется общая слизистая подошва, на которой делящиеся клетки сидят пучками или, в случае ветвления подошвы, каждая из них имеет индивидуальную слизистую ножку.
Уникальную колонию из 200-500 клеток, соединенных в поясковой зоне перегородками органической природы, образует Coenobiodiscus muriformis. Эти колонии воспроизводятся без прохождения одноклеточной стадии.
Клетки диатомовых водорослей имеют типичное эукариотическое строение. Цитоплазма в них образует пристенный слой либо скапливается у полюсов или в центре клетки, соединяясь цитоплазматическими мостиками. У центрических диатомей ядро лежит в центральной массе цитоплазмы или в пристенном слое ближе к гипотеке, а у пеннатных - в цитоплазматическом мостике в непосредственном контакте с хлоропластом ближе к эпитеке. В клетке имеется одна или несколько вакуолей с клеточным соком, пронизанных многочисленными тяжами цитоплазмы.
Плазмалемма плотно прилегает к панцирю. Галофильные виды очень хорошо пропускают через плазмалемму ионы солей. Проницаемость плазмалеммы для неэлектролитов выше, чем у других растительных клеток, но не у всех диатомей одинакова. Так, у некоторых из них высока проницаемость для сахаров, а у Caloneis obtusa (W. Sm.) Cl. и Cymbella aspera (Ehr.) Cl. плазмалемма почти непроницаема для них. Целлюлозной оболочки, в отличие от большинства водорослей, у диатомовых нет.
У подробно изученных таксонов ядро грушевидное, но описаны и другие формы ядер: овальные (Cylindrotheca fusiformis Reiman et Lewin), чечевицеобразные (Melosira varians Ag.), H-образные (Surirella ovalis Breb.), гантелевидные (Lauderia annulata Cl.).
Ядро окружено двойной пористой мембраной. Многочисленные поры пронизывают боковые поверхности ядра и отсутствуют в его основании. У Strepthothoca thamesis ядро имеет 6-7 тыс. пор [93]. Размер пор 80-100 им. Отверстие поры окружено тонкой диафрагмой с центральной гранулой около 19 нм в диаметре. В ядре расположены 1-8 ядрышек - наиболее плотных образований из всех клеточных органоидов. Из-за отсутствия мембраны границы ядрышек не имеют резких очертаний.
Хлоропласты диатомовых водорослей разнообразны по форме, размерам и положению в клетке. Обычно они располагаются в пристенном слое цитоплазмы, прилегая к створкам, реже к пояску; у колониальных диатомей с лентовидными колониями они всегда сдвинуты в сторону пояска. У большинства центрических диатомовых хлоропласты мелкие, многочисленные, в виде зернышек или дисков. Такие же хлоропласты характерны и для бесшовных из класса Pennatophyceae. У более высокоорганизованных пеннатных хлоропласты крупные, немногочисленные, часто с лопастными краями или перфорированные. Форма и размеры хлоропластов у диатомовых водорослей изменчивы.
Хлоропласт окружен системой мембран. Его оболочка состоит из двух мембран. Кроме того, к нему прилегает одна или несколько цистерн эндоплазматического ретикулюма и выросты ядерной оболочки. В этом случае оболочка ядра на некоторых участках становится одновременно и оболочкой хлоропласта, внутренняя мембрана оболочки хлоропласта одновременно является внешней мембраной ядра и наоборот. Этот признак выявлен не у всех диатомовых, например, связь хлоропласта ядра не удалось обнаружить у Melosira varians Ag. На некоторых участках внешней мембраны хлоропласта находятся рибосомы.
В строме хлоропласта расположены тилакоиды - двухмембранные уплощенные цистерны. Тилакоиды диатомовых, в отличие от высших растений и некоторых водорослей, не организованы в граны, а образуют пачки из трех, иногда большего числа цистерн. Количество тилакоидов в пачке непостоянно и зависит как от систематической принадлежности, так и от условий жизни организма. Характерно образование анастомозов - ответвлений тилакоидов - между пачками. Один или несколько тилакоидов расположены по периферии хлоропласта, непосредственно под его оболочкой. Это опоясывающие тилакоиды, которые помимо диатомовых обнаружены у красных, бурых и золотистых водорослей. Под опоясывающими гилакоидами, обычно на полюсах хлоропласта, выявлены светлые зоны электронно-плотного фибриллярного материала. Считают, что это нуклеоиды или генофоры хлоропласта, содержащие ДНК.
У диатомовых с многочисленными хлоропластами обнаружены связанные с ними структуры - микрофиламенты, которые расположены в слое цитоплазмы между хлоропластом и плазмалеммой. Считается, что они обусловливают движение хлоропластов от периферии к ядру при усилении освещения и наоборот. То, что это микрофиламенты, а не микротрубочки подтверждается ингибированием движения хлоропластов цитохалазином В, но не колхицином.
Хлоропласт содержит один или несколько пиреноидов, имеющих форму линзы, веретена, вогнутой пластинки и расположенных на участках хлоропласта, контактирующих с вакуолями. Тилакоиды хлоропласта обходят пиреноид, прерываются перед ним или пронизывают его в различных направлениях. У некоторых диатомовых обнаружены перфорированные участки в пачках тилакоидов между пиреноидом и оболочкой хлоропласта, обращенной к вакуоли. У ряда видов выявлена мембран, окружающая пиреноид. Матрикс пиреноида состоит из гранулярного материала, более электронно-плотного, чем матрикс хлоропласта. Иногда пиреноид имеет упорядоченную кристаллическую структуру, ламеллярное строение.
Окраска хлоропласта, обусловленная составом пигментов, изменяется от светло-желтой, золотистой до зеленовато-бурой и посмертно приобретает зеленый цвет вследствие растворения бурых пигментов в мертвых клетках. Более интенсивно окрашены бентосные виды, массовое развитие которых создает бурый налет на других гидробионтах, дне водоема и различных погруженных в воду предметах. Подверженные сильному освещению планктонные виды не окрашивают воду даже при массовом развитии, а при наблюдении в микроскоп имеют светло-желтый, золотистый цвет.
В хлоропластах диатомовых содержатся пигменты: хлорофиллы а и с, β- и ε-каротины, ксантофиллы (фукоксантин, диатоксантин, неофукоксантины А и В, диадиноксантин). Есть указание о наличии следов феофитина. Пигментная система диатомовых водорослей обеспечивает фотосинтез на глубине до 50 м благодаря дополнительным пигментам из группы фукоксантинов, которые передают поглощенную ими энергию хлорофиллу а. У гетеротрофных бесцветных диатомовых водорослей (Nitzschia) пигменты отсутствуют.
Продукты ассимиляции диатомовых - липиды, волютин, хризоламинарин - располагаются в питоплазме в вакуолях. Капли липидов обычно окружены мембраной. Крупные голубоватые капли волютина (тельца Бючли) занимают определенное положение: у полюсов клетки у родов семейства Nitzschiaceae, по обе стороны цитоплазматического мостика у родов семейства Naviculaceae. Хризоламинарин близок к ламинарину бурых водорослей. По продуктам ассимиляции диатомовые резко отличаются от других водорослей, образующих в ходе фотосинтеза углеводы.
Всюду в цитоплазме располагаются митохондрии, имеющие вид палочкообразных, нитевидных и зернистых элементов. Митохондрии диатомовых имеют трубчатые кристы.
В небольшой впадине на одной из сторон ядра расположена центросома (постоянный полярный комплекс, центр микротрубочек), которая у разных представителей диатомовых отличается в деталях строения, но всегда содержит цитоплазматические микротрубочки. Центросома формирует веретено деления при кариокинетическом делении клетки.
Вблизи ядра, со стороны, удаленной от хлоропласта, располагается аппарат Гольджи, состоящий из нескольких диктиосом. Диктиосома состоит из 4-12 цистерн, образующихся при слиянии пузырьков, которые отпочковывает внешняя мембрана ядерной оболочки. В различных диктиосомах одной и той же клетки число цистерн одинаково. Со стороны, противоположной ядру, диктиосомы отпочковывают пузырьки нескольких типов: 1) пузырьки разнообразной формы с гладкой поверхностью; 2) крупные пузырьки с гранулярным содержимым; 3) мелкие опоясанные пузырьки с гранулярным содержанием и палочковидными частицами на поверхности.
Диктиосомный аппарат диатомовых очень развит. Отмечена высокая активность диктиосом в стадии гаметогенеза клетки, когда продуцируемые ими пузырьки лизируют пикнотические постмейотические ядра. По мнению ряда авторов, диктиосомы принимают участие в отложении кремниевого панциря и выделении слизи - функциях, ярко выраженных у диатомовых.
Между плазмалеммой и диктиосомой многих диатомовых, чаще в стареющих клетках, обнаружен везикулярный комплекс - совокупность многочисленных пузырьков различной формы и размеров. Предполагают, что это продукт активности диктиосомы, пул мембран клетки.
В периферической части клетки находится силикалемма - сложная система цистерн и трубочек, в которых обнаружены элементы кремниевого панциря. Внешняя мембрана цистерн силикалеммы тесно примыкает к плазма лемме. По мнению одних исследователей, пузырьки силикалеммы формируются диктиосомами, по мнению других, - эндоплазматическим ретикулюмом и даже плазмалеммой. Элементы силикалеммы несут, вероятно, двойную функцию: формирования панциря за счет их содержимого и образования новой плазмалеммы за счет мембран пузырьков.
Основной способ размножения диатомовых водорослей - вегетативное деление клетки надвое. Деление обычно происходит ночью и на рассвете и осуществляется по-разному у разных видов, а также у одних и тех же видов в зависимости от условий среды. В культуральной среде планктонные диатомовые могут делиться 3-8 раз в сутки, бентосные - один раз в 4 суток. Наиболее интенсивного развития диатомовые водоросли достигают весной и осенью.
Перед делением в протопласте скапливаются капли масла, он увеличивается в размерах, раздвигает эпитеку и гипотеку так, что они соприкасаются лишь краями поисковых ободков. У многих видов митозу предшествует деление хлоропласта. При некоторых различиях в деталях у исследованных центрических и пеннатных диатомовых общая схема митоза на ультраструктурном уровне одинакова (рис. 15.20).
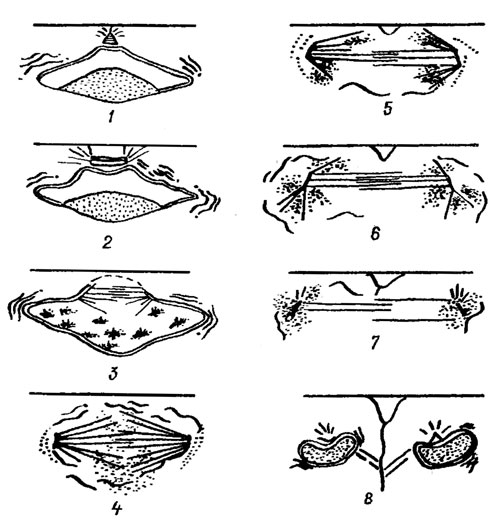
Рис. 15.20. Схема митоза Melosira varians Ag.: 1 - интерфаза; 2 - профаза (в выемке на вершине ядра расположено веретено деления с центриолями; радиальные микротрубочки проникают в ядро через поры); 3 - поздняя профаза (микротрубочки проникают в глубокие впячивания ядерной оболочки, при этом разрушаемой, и вместе с центр нолями погружаются в матрикс ядра; хроматин конденсируется, ядрышко разрушается); 4 - метафаза (веретено достигает полного развития; микротрубочки погружены в хроматин, асимметрично расположенный вокруг веретена); 5 - анафаза (передвижение хроматина к полюсам веретена; появление широкого впячивания плазмалеммы; начало цитокинеза); 6 - телофаза (концентрация хроматина вокруг полярных центриолей); 7 - поздняя телофаза (освобождение цетриолей от микротрубочек; формирование оболочки ядра; завершение цитокинеза); в - клетки после цитокинеза
Многократные вегетативные деления приводят к постепенному уменьшению размеров клеток, получающих гипотеку материнской клетки. У некоторых видов они уменьшаются в 3 раза по сравнению с первоначальными. Восстановление первоначальных размеров клеток происходит во время прорастания покоящихся спор, покоящихся клеток и в результате полового процесса, сопровождающегося образованием ауксоспор. В культуре переход к образованию ауксоспор сопровождается морфологическими изменениями отдельных клеток и колоний в целом: у Sceletonema costatum (Grev.) Cl. за 2-3 недели образовывались изогнутые, спиралевидные колонии; у Melosira moniliformis (О. Mull.) Ag. - длинные веревкообразные колонии; у Actinocyclus ehrenbergii Ralfs - удлиненные, сильно пигментированные клетки (материнские клетки ауксоспор); у Thalassiosira excentrica (Ehr.) Cl. - за 10-14 дней - нежизнеспособные ауксоспоры ("псевдоауксоспоры") [178]. Считается, что до начала спорообразования клетки проходят внутреннюю перестройку, направленную на ликвидацию отклонений в метаболизме, вызванных нарушением соотношения объемов ядра и цитоплазмы в результате митотических делений. При этом происходит дифференциация клеток по их роли в репродуктивном процессе, так как не все клетки, достигшие определенного размера, переходят к спорообразованию.
Перед началом полового процесса две клетки сближаются, покрываются общим слизистым чехлом и раздвигают створки. После оплодотворения образуется зигота, которая без стадии покоя начинает расти, увеличивается в размерах и превращается в ауксоспору ("растущую спору"). При созревании ауксоспора превращается в инициальную клетку, значительно превосходящую по размерам исходную материнскую и иногда отличную от нее по форме. По положению относительно материнской клетки и связи с ней различают свободные, боковые, конечные, интеркалярные и полуинтеркалярные ауксоспоры (рис. 15.21). Следует подчеркнуть, что диатомовые водоросли - единственная группа растительных организмов, в жизненном цикле которых есть стадия ауксопорообразования.
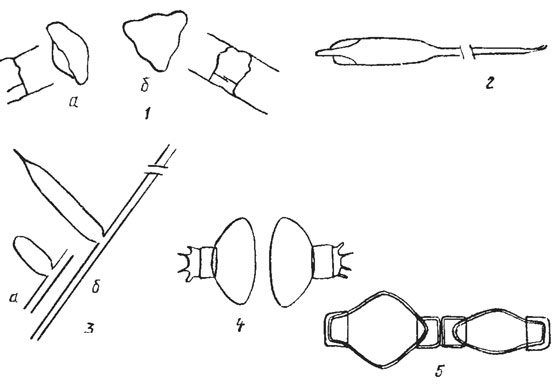
Рис. 15.21. Положение и связь ауксоспор диатомовых с материнской клеткой: 1 - свободная ауксоспора Lithodesmium undulatum Ehr. (a - вид со стороны пояска с первой инициальной клеткой, б - вид со стороны створки); 2 - конечная ауксоспора Rhizosolenia alata Bright с первой инициальной клеткой, образовавшейся внутри ауксоспоры и разорвавшей ее стенку; 3 - боковая ауксоспора Rhizosolenia shrubsolii Cl. (а - общий вид, б - дистальная часть сброшенной стенки ауксоспоры, замещенной первой инициальной текой); 4 - полуинтеркалярная ауксоспора Odontella regia; 5 - интеркалярная ауксоспора Melosira varians Ag
У диатомовых выявлено несколько типов полового процесса. При изогамном половом процессе (рис. 15.22, 1-5) в двух материнских клетках образуется по две неподвижные гаметы, которые копулируют (сливаются; попарно (виды родов Amphora Ehr., Epithemia Breb., Rhopalodia О. Mull., Surirella Turp.).
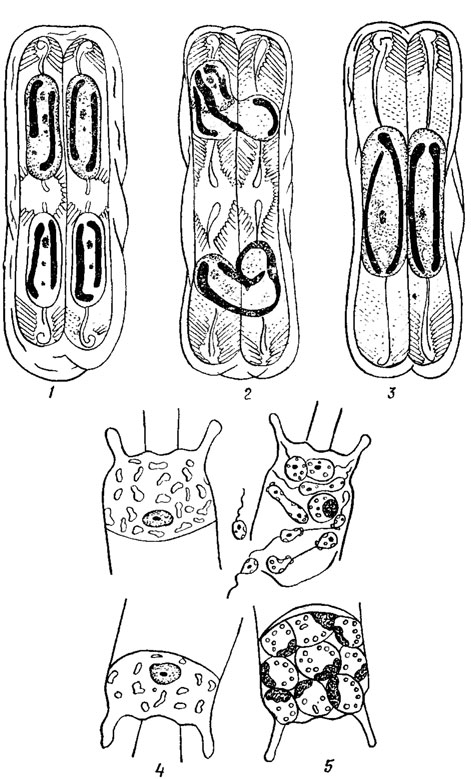
Рис. 15.22. Типы полового процесса у диатомовых водорослей: 1-3 анизогамный (Pirnnularia sp.); 4, 5 - оогамный (Biddtilphia mobiliensis Bailey)
Анизогамный (гетерогамный) половой процесс протекает двояко. В первом случае в ходе последовательных мейотического и митотического делений в каждой материнской клетке образуется по одной подвижной и одной неподвижной гамете. Подвижные гаметы передвигаются к неподвижным и сливаются с ними. Этот тип характерен для большинства представителей семейства Naviculaceae и некоторых видов рода Nitzschia. Во втором случае в одной клетке обе гаметы неподвижные, в другой - обе подвижные, переходящие в клетку с неподвижными гаметами. Такой тип анизогамии характерен для Navicula halophila (Grun.) Cl. и Synedra ulna (Nitzsch.) Ehr.
При оогамном половом процессе (рис. 15.22, 4, 5) женская репродуктивная клетка (оогоний) производит одну яйцеклетку (виды рода Stephanopyxis Ehr., Melosira varians Ag.) или две (Biddulphia mobiliensis Bail.), а мужская репродуктивная клетка (сперматогоний) образует два (Melosira varians Ag.) или четыре (Biddulphia rhombus (Ehr.) W. Sm., Cyclotella sp.) сперматозоида, оплодотворяющих яйцеклетку. У пентрических диатомей, в отличие от других водорослей, сначала образуется большое число мелких сперматогониев, а мейоз происходит в самый последний момент, непосредственно перед обособлением гамет. Обычно же при гаметической или спорической редукции у других водорослей сначала совершается мейоз, затем при митотических делениях увеличивается число ядер и лишь после этого формируется большое число гамет или гаплоидных зооспор.
У диатомовых в ходе редуцированного полового процесса репродуктивная клетка развивается в зиготу и ауксоспору через апомиксис или автогамно (Chaetoceros borealis Bailey, Cyclotella meneghiniana Kütz.).
Клетки, связанные с половым процессом, имеют некоторые структурные отличия от обычных клеток. Так, в сперматозоидах Lithodesmium undulatum Ehr. отсутствуют диктиосомы, а хлоропласты более мелкие и упрощенные; в аксонеме жгутиков сперматозоидов Lithodesmium undulatum Ehr. и Pleurosira laevis (Ehr.) Compere нет двух внутренних микротрубочек, жгутики покрыты мастигонемами - волосками диаметром 11 нм.
У многих морских планктонных диатомей в клетках возникают мелкие тельца - микроспоры (рис. 15.23). Они бывают со жгутиками и без жгутиков, с хлоропластами и бесцветные. Образование микроспор особенно характерно для видов рода Chaetoceros Ehr., у которых наблюдалось и их прорастание. Пути образования и природа микроспор не установлены.
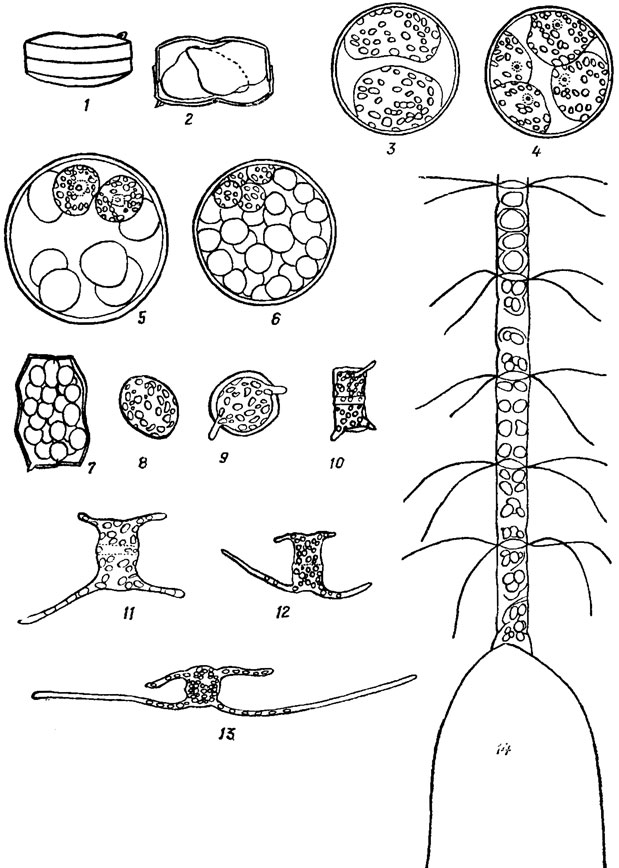
Рис. 15.23. Образование и прорастание микроспор у диатомовых водорослей: 1-7 - последовательные стадии образования микроспор у Coscinodiscus jonesianus (Grev.) Ostf. 8-13 - последовательные стадии прорастания микроспор у Chaetoceros danicus Cl.; 14 - различные стадии образования микроспор у Chaetoceros curvisetus Cl
При неблагоприятных условиях диатомовые водоросли переходят в состояние покоя. В ходе образования покоящихся клеток протопласт передвигается к одному из концов клетки и, вследствие потери клеточного сока, сильно сжимается. Жизнедеятельность этих клеток возобновляется при наступлении благоприятных условий. Некоторые пресноводные планктонные озерные виды в зимний период погружаются на дно водоемов, где пребывают в состоянии покоя или пониженной жизнедеятельности до начала нового вегетационного периода.
У большинства диатомей образованию покоящихся спор (рис. 15.24; 15.25) предшествует деление вегетативной клетки на две, каждая из которых в дальнейшем становится материнской клеткой споры. Протопласт материнской клетки сжимается, округляется, на поверхности его возникает первичная створка споры, затем вторичная, которая выдвигается своими краями в края первичной. Содержимое споры гомогенно. Структура створок споры постоянна для каждого вида и отличается от структуры вегетативной клетки. Это один из наиболее важных видовых признаков. Как правило, материнская клетка производит одну экзогенную, полуэндогенную или эндогенную спору (у Rhizosoleniasetigera Braight. - две). Зрелая экзогенная спора находится вне материнской клетки; одна створка зрелой полуэндогенной покоящейся споры включена в материнскую клетку, другая остается свободной; зрелая эндогенная покоящаяся спора находится внутри материнской клетки (рис. 15.26). Спора прорастает в вегетативную клетку, размер которой значительно превышает размер самой споры.
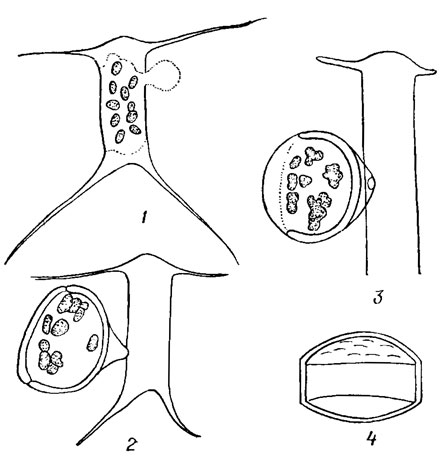
Рис. 15.24. Последовательные стадии (1-4) образования покоящейся споры у Chaetoceros heterovalvatus Schütt
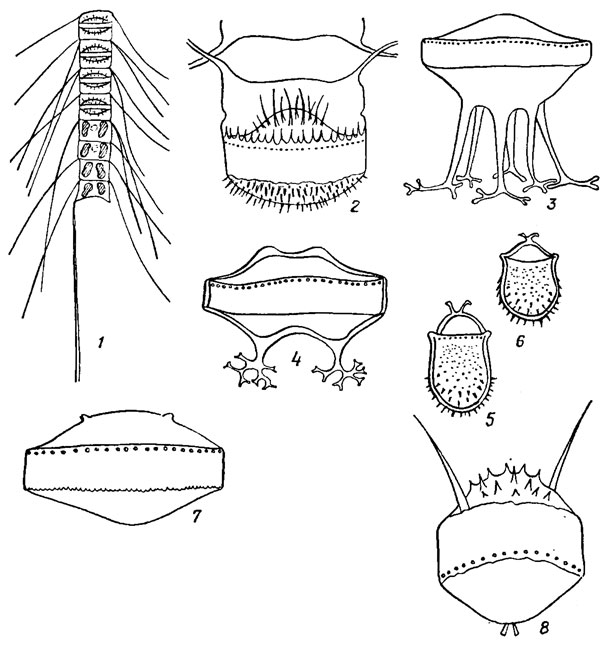
Рис. 15.25. Споры видов рода Chaetoceros Ehr.: 1 - цепочка с молодыми спорами Chaetoceros abnormis Pr.-Lavr.; 2 - Ch. seiracanthus Gran; 3 - Ch. subsecundus (Grun.) Hust.; 4 - Ch. lorenzianus Grun.; 5, 6 - Ch. paulsenii Ostf.; 7 - Ch. compress us Laud.; 8 - Cn. rigidus Ostf
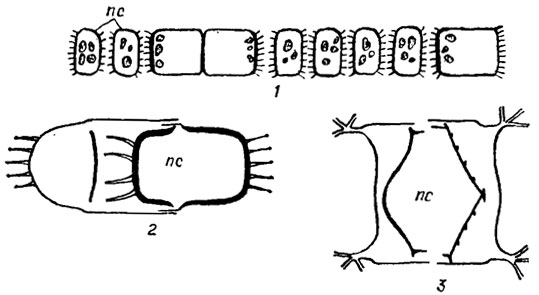
Рис. 15.26. Виды покоящихся спор (пс) диатомовых водорослей: 1 - экзогенные (Detonula confervacea (Cl.) Gran); 2 - полуэндогенная (Stephanopyxis turris (Grev. et Arn.) Ralfs); 3 - эндогенная (Chaetoceros compressus Laud.)
Цикл развития диатомовых водорослей проходит в диплоидной фазе с гаметической редукцией.
Диатомовые не имеют прямых родственных связей с другими отделами водорослей, но ряд признаков (общность пигментного состава и продуктов ассимиляции, наличие кремния в клеточных покровах и покоящихся спор) указывает на их отдаленное родство с желтозелеными, золотистыми и бурыми водорослями. Все указанные отделы, вероятно, произошли от общих фотосинтезирующих жгутиковых (мужские гаметы центрических диатомовых одножгутиковые с редуцированной центральной парой фибрилл в жгутике по схеме "9 + 0") эукариотических предков с преобладанием бурых пигментов. Впервые это сходство обосновал А. Пашер, выделив единую филу Chrysophyta с отделами Chrysophyceae, Diatomeae, Heterocontae. M. Шадефо [371] рассматривает диатомовые как один из классов подотдела Chrysophyceae, разделенного на 5 классов: Chrysophycineae, Xanthophycineae, Bacillariophycineae, Silicophycineae, Craspedophycineae (Choanoflagellatae). E. Эттл [405] также выделяет диатомовые в ранге класса из отдела Chrysophyta: Chrysophyceae, Haptophyceae, Xanthophyceae, Eustigmatophyceae, Bacillariophyceae. В отечественной литературе принято рассматривать диатомовые в ранге самостоятельного отдела [73, 239]. Новейшую классификацию диатомовых - см. [105; Т. 2].
Хорошая сохранность панциря диатомовых водорослей в ископаемом состоянии позволяет проследить пути их эволюции на основе палеонтологических данных. Наиболее древние ископаемые диатомовые водоросли известны из отложений раннего мела (мезозойская эра). Находки, относящиеся к юрскому периоду [105], оказались ошибочными или требуют дополнительных исследований.
Существует две точки зрения на древность диатомовых. Согласно первой гипотезе, основанной на возрасте ископаемых остатков, диатомовые считаются относительно молодой группой водорослей, появившейся в мезозое, около 200 млн лет назад. Согласно второй гипотезе, основанной на богатстве и разнообразии меловой флоры, - это древняя группа, существовавшая в палеозое, прошедшая длительное эволюционное развитие до мелового периода. В позднем мелу уже насчитывается около 397 видов и 200 родов центрических, появляются первые бесшовные пеннатные диатомовые водоросли [299]. С конца палеоцена - начала эоцена (палеогеновый период кайнозойской эры) известны шовные пеннатные формы, а бурное их развитие начинается с нижнего миоцена (неогеновый период кайнозойской эры). На территории СССР известно 1580 видов и 189 родов палеогеновых диатомовых водорослей. Совершенствование шва занимает центральное место в эволюции диатомовых, так как именно это обеспечило быстрое развитие пеннатных форм и их преобладание над центрическими.
В пресных водоемах первые диатомовые зарегистрированы в позднепалеогеновых озерных отложениях, в неогене они достигли здесь наибольшего развития. Современная флора диатомовых имеет наибольшее сходство с флорой морей и континентальных водоемов плейстоцена (четвертичный период кайнозойской эры) и характеризуется господством пеннатных форм.
Современная классификация диатомовых водорослей, основанная на морфологии и структуре панциря, оформлена к 30-м годам XX ст. Ф. Хустедтом и базируется на системах Ф. Шютта, Е. Эструпа и Дж. Карстена [189]. Исследования диатомовых с помощью сканирующей и трансмиссионной электронной микроскопии позволили выявить новые элементы структуры панцирей, критически подойти к таксономической значимости признаков, обнаруженных с помощью световой микроскопии. Накопление этих данных привело к необходимости пересмотра существующей системы, предложению ряда новых систем и схем (цит. по: [189]).
В настоящем издании диатомовые водоросли рассматриваются как самостоятельный отдел Bacillariophyta с двумя классами: центрических (Centrophyceae), включающем 5 порядков, и пеннатных (Pennatophyceae), включающем 4 порядка [188].
Диатомовые водоросли имеют широкое распространение и населяют всевозможные биотопы: пресные и соленые, стоячие и текучие водоемы, влажные скалы, почву и даже пахотные земли. Широкое распространение обусловлено пластичностью группы в целом по отношению к различным экологическим факторам и стенобионтностью отдельных видов к тем же самым факторам внешней среды.
В водных экосистемах диатомовые водоросли - круглогодичные доминанты; они участвуют в образовании различных фитоценозов всех типов. В планктоне морей и океанов преобладают центрические, встречаются бесшовные диатомовые: в литоральном - обычно более крупные, толстостенные, лишенные выростов и щетинок, Actinoptychus Ehr., Aliscus Ehr.; в неритическом (прибрежном) - Leptocylindrus Cl., некоторые Chaethoceros Ehr., Rhizosolenia Ehr., Actinocyclus Ehr.; в океаническом (пелагическом) - обычно тонкостенные, с длинными щетинками и выростами Planctoniella Schutt, Sceletonema Grev., Lauderia Cl., Chaetoceros Ehr., Rhizosolenia Ehr., Bacteriastrum Schadb. В открытой части океана диатомовые могут распространяться до глубин 80-350 м, где они образуют глубоководный "теневой планктон". Это обусловлено способностью некоторых видов переходить на гетеротрофное питание при обилии органических веществ в воде. В планктоне пресных водоемов преобладают пеннатные диатомовые; глубина их распространения зависит от прозрачности воды.
Бентосные (донные) фитоценозы отличаются большим разнообразием и количеством диатомовых водорослей, среди которых преобладают пеннатные диатомеи (Navicula Bory, Campylodiscus Ehr. ex Kütz., Surirella Turp., Pinnularia Ehr., Gyrosigma Hass.). Они обычно обитают на глубине не более 50 м.
Наиболее богаты по видовому и количественному составу ценозы обрастаний. Диатомовые господствуют среди обрастаний макроскопических водорослей пресных водоемов (Cocconeis Ehr., Cymbella Ag., Gomphonema Ag.) и морей (Lycmophora Ag., Rhabdonema Kütz., Achnanthes Bory, Mastogloia Thw.). Особенно многообразны диатомовые среди эпифитов. Известны случаи обрастаний диатомовыми водорослями животных, причем иногда наблюдается приуроченность определенных видов диатомовых к определенным видам животных: Synedra cyclopum Brutschy обитает на циклопах, Bennettella ceticola (Nelson et Bennet) R. Holes - на коже китов.
Есть указания на случаи комменсализма: Nitzschia dissipata (Kütz.) Grun. обнаружена в слизи Cymbella lacustris (Ag.) Cl., C. prostrata (Berk.) Cl., C. ventricosa Kütz.; Navicula endophytica Hasle обитает в межклеточном пространстве рецептакулов макрофита Ascophyllum nodosum (L.) Le Jolis. Описаны случаи экзосимбиоза с недиатомовыми: Solenicola setigera обитала на Dactyliosolen mediterraneus Н. Perag., жгутиковая Biocaeca mediterranea на Sceletonema costatum (Grev.) Cl., Chaetoceros anastomosans Grun., Ch. lorensianus Grun., Cerataulina bergonii H. Perag., Nitzschia seriata Cl., Thalassiothrix frauenfeldii Grun. Диатомовые водоросли способны эндосимбиотически существовать в протопласте фораминифер.
Нередко диатомовые становятся хозяевами паразитов, причем наблюдается приуроченность паразита к определенным видам. Известно 17 видов низших грибов, паразитирующих на 41 виде диатомовых водорослей (цит. по: [105, кн. 1]). Перидинея Paulsenella chaetoceratis (Pauls.) Chatton паразитирует на Chaetoceros borealis Bail. и Ch. decipiens Cl.
Среди диатомовых выявлены облигатные гетеротрофы с бесцветными пластидами или вовсе без них: Nitzschia alba Lewin, N. putrida, Synedra hyalina Provasch и N. leucosigma, причем два последних вида способны развиваться в анаэробной среде, усваивать лактат и гидролизованный протеин.
Диатомовые водоросли вегетируют в диапазоне 0-70°С, но способны переносить в состоянии покоя более высокие и более низкие температуры. Оптимальная температура и пределы ее изменения индивидуальны и очень разнообразны для разных видов.
У некоторых диатомовых водорослей температура влияет на форму колоний и структуру панциря: у Asterionella летом образуются только звездчатые колонии, зимой - звездчато-зигзаговидные и зигзаговидные. Настоящий сезонный диморфизм (сезонные изменения структуры панциря) отмечены у Rhizosolenia hebetata (Bail.) Gran, Rh. semispina Hensen, Corethron inerme Karst., Chaetoceros concavicornis Mangin.
По отношению к солености воды выделены четыре группы диатомовых водорослей (по шкале галобности Кольбе): полигалобы (соленость более 40‰), эвгалобы (30-40‰), мезогалобы (5-20‰), олигогалобы (0-5‰) [268].
Наиболее богата диатомовыми флора эвтрофных озер (преимущественно равнинных) с большим содержанием солей. В планктоне распространены Melosira islandica О. Mull., Cyclotella comta (Ehr.) Kütz.; на дне - виды родов Navicula, Nitzschia, Gyrosigma, Pinnularia; в обрастаниях - виды родов Cymbella Ag., Gomphonema Ag., Synedra Ehr., Achnanthes Bory, Cocconeis Ehr. Диатомовая флора водохранилищ, отчасти прудов, близка к флоре эвтрофных озер.
На постоянно увлажняемых субстратах (влажные скалы, камни, подушки мхов и лишайников) диатомовые развиваются в большом количестве. Часто они заключены в выделяемую ими слизь. В почве диатомовые распространены до глубины 1 м, преобладают мелкие подвижные формы (Navicula atomus (Näg.) Grun., Hantzschia amphioxys (Ehr.) Grun., Pinnularia borealis Ehr.). В почвах обнаружено более 50 различных видов, обычно убиквистов. Свыше 30 видов диатомовых способны обитать на снегу и льду, выдерживая понижение температуры до -36°С.
В водах Мирового океана, где каждой широтной зоне свойственна специфическая флора, особенно богат видовой состав диатомовых бореальной зоны, отличающейся оптимальной для их развития температурой (10-20°С).
Космополиты особенно часто встречаются в континентальных водоемах. В некоторых континентальных водоемах (например, в оз. Байкал) есть эндемики. В Черном, Азовском и Каспийском морях известны реликты, сохранившиеся от верхнетретичных морей Черноморского бассейна.
Роль в природе и практическое значение диатомовых водорослей очень велики. В альгофлоре морей и океанов они составляют до 80% и более систематического состава водорослей, создают 50% всей органической массы океана и почти 1/4 глобальной продукции живого вещества Земли, ежегодно поглощая из Мирового океана около 10 млрд т углерода.
Диатомовые водоросли - основное звено трофических цепей водных экосистем. Они играют основополагающую роль в продуктивности водоемов. В 100 г органического вещества диатомовых содержится 40% белков, 30 углеводов, 30% липидов [19]. Их калорийность составляет 525 кал, т. е. превышает калорийность всех других групп водорослей. Высокая калорийность сопровождается легкой усвояемостью органического вещества. В составе белков присутствуют все незаменимые аминокислоты. В составе липидов много ненасыщенных жиров, свободных жирных кислот, в том числе незаменимая линолевая кислота. Диатомовыми водорослями питаются различные животные, низшие ракообразные - веслоногие и ветвистоусые рачки, которые составляют наиболее важную часть зоопланктона по его значению в питании промысловых рыб. При отмирании диатомовые дают массу детрита и растворимых органических веществ, используемых для питания бактериями и простейшими.
Эти одноклеточные водоросли способны интенсивно делиться, давая до четырех поколений в сутки, что нередко вызывает цветение воды, появление неприятных запахов в питьевой воде. Так, массовое развитие Stephanodiscus hantschii Grun. обусловливает рыбный запах, Cyclotella meneghiniana Kütz. - травянисто-тинистый [236].
Ежегодно поглощая из Мирового океана около 3 млрд т кремния, диатомовые водоросли играют основную роль в его круговороте.
Особое значение имеют диатомеи в экологическом мониторинге, являясь индикаторами загрязнения водной среды. В настоящее время известно около 100 таксонов - индикаторов загрязненности воды (например, среди ксенобионтов - Achnanthes flexella (Breb.) Brun., Amphora normanii Rabench., Cymbella cesatii (Rabench.) Grun.; среди мезосапробионтов - Nitzschia angustata (W. Sm.) Grun., N. palea (Kütz.) W. Sm., N. hungarica Grun., Cyclotella meneghiniana Kütz.).
Важнейшая роль принадлежит диатомовым в осадконакоплении. При их отмирании на дне водоемов образуются диатомовые и сапропелевые илы, диатомиты мощностью в несколько сот метров. Твердые кремнеземные оболочки диатомовых сохраняются длительное время в ископаемом состоянии, поэтому диатомовый анализ широко применяют как один из палеонтологических методов. Видовой состав створок диатомеи в осадках позволяет, используя принцип актуализма, судить об условиях осадконакопления и климате прошедших эпох.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'