12. Отдел 3. Криптофитовые водоросли - Cryptophyta (Cryptophycophyta, Cryptophyceae) (З. И. Ветрова)
Криптофитовые представляют собой небольшую своеобразную группу пресноводных и солоноватоводных одноклеточных организмов, в большинстве имеющих монадный тип структуры. Изредка встречаются пальмеллоидные формы. Существование коккоидных форм оспаривается и нуждается в подтверждении. Отдел объединяет около 100 видов, относящихся к 21 роду.
Индивиды криптофитовых водорослей имеют обычно выпуклую спинную и вогнутую брюшную стороны. Они могут быть яйцевидными, обратнояйцевидными, эллипсоидными, бобовидными, клиновидными, линзовидными, веретеновидными, грушевидными, обратноконусовидными, шаровидными и т. п.; обычно уплощенные, часто винтообразно скрученные (рис. 12.1), не метаболичные или метаболичные.
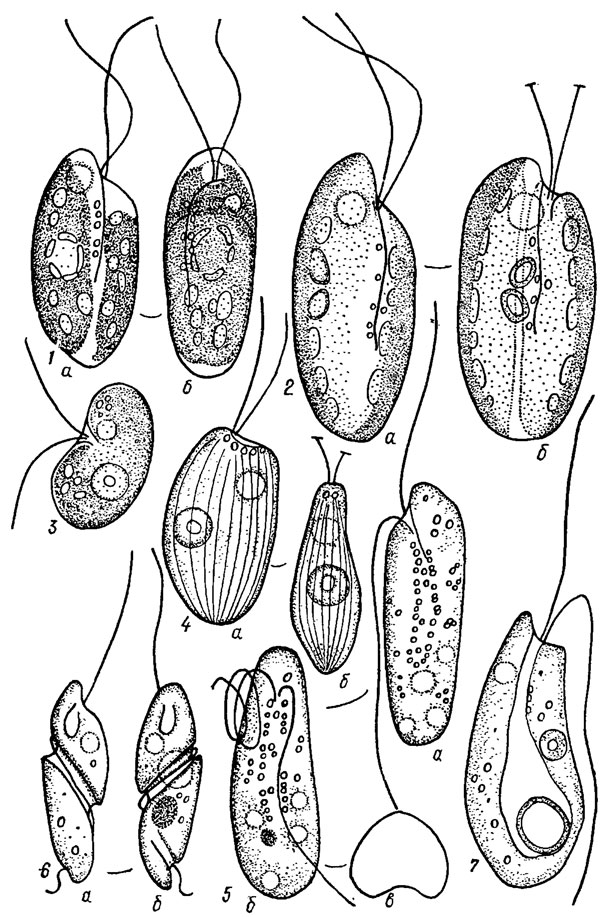
Рис. 12.1. Форма тела криптофитовых водорослей: 1 - Cryptochrysis commutata Pasch. (a - вид сбоку, б - с вентральной стороны); 2 - С. pochmannii Hub.-Pest. (а - вид сбоку, б - с вентральной стороны); 3 - Sennia parvula Skuja; 4 - Cyathomonas truncata (Fres.) From. (a - вид сбоку, б - с брюшной стороны); 5 - Katablepharis hyalurum Skuja (а - вид сбоку, б - с брюшной стороны, в - сверху); 6 - Cryptaulax acroporus (Skuja) Skuja (a - вид сбоку, б - со спинной стороны); 7 - Pnyllomitus amylophagus Klebs
Размеры клеток сильно варьируют у видов разных родов:

Передний конец клетки с углублением - воронкой, он косо срезан, от него проходит более или менее заметная продольная бороздка, не доходящая до заднего конца. Глотка мешковидная, по краю ее и на поверхности находятся трихоцисты (эджектосомы) в виде палочек или зернышек, которые при раздражении выбрасывают тонкие нити.
Клетки криптофитовых имеют покров в виде перипласта, довольно плотного, часто продольно или спирально исчерченного в связи с наличием в наружном слое цитоплазмы своеобразных, выделяющих слизь телец, которые расположены правильными рядами. Глотка преимущественно хорошо развита, с расположенными рядами трихоцистами. Жгутиков 2, почти одинаковой длины, при движении или направленных вперед, или один - вперед, другой - назад. Корешковая система наиболее детально исследована у Cryptomonas ovata Ehr. Вся их сложная система образована ризостилем, тремя микротрубочковыми и двумя фибриллярными корешками. Ризостиль направлен к заднему концу клетки. Он образован лентой из 6-8 микротрубочек. От каждой из них отходит тонкий вырост, связывающий микротрубочки с тонкой пластинкой. Три микротрубочковых корешка расположены латерально. Изогнутая лента из 4 микротрубочек связана с дорсальной митохондрией. Корешок из 2 микротрубочек проходит рядом с сократительной вакуолью. Третий корешок из 4 микротрубочек расположен непосредственно под перипластом. К нему прилегает поперечно исчерченный фибриллярный корешок. Другой корешок из микрофиламентов не имеет исчерченности и связан с вентральной митохондрией. В целом у криптофитовых корешковая система довольно своеобразна: в глубь клетки идет ризостиль, представленный многослойной структурой. Форму его переднему концу придают боковые корешки, из которых 3-4 микротрубочковых и 2 фибриллярных, один из которых ассоциирован с микротрубочковым корешком. Корешковая система довольно однородна в пределах класса, все отклонения можно объяснить вариациями размеров клеток.
Ядро эукариотического типа, одно, расположено в задней части клетки или, реже, посередине, имеет ядрышко (рис. 12.2).
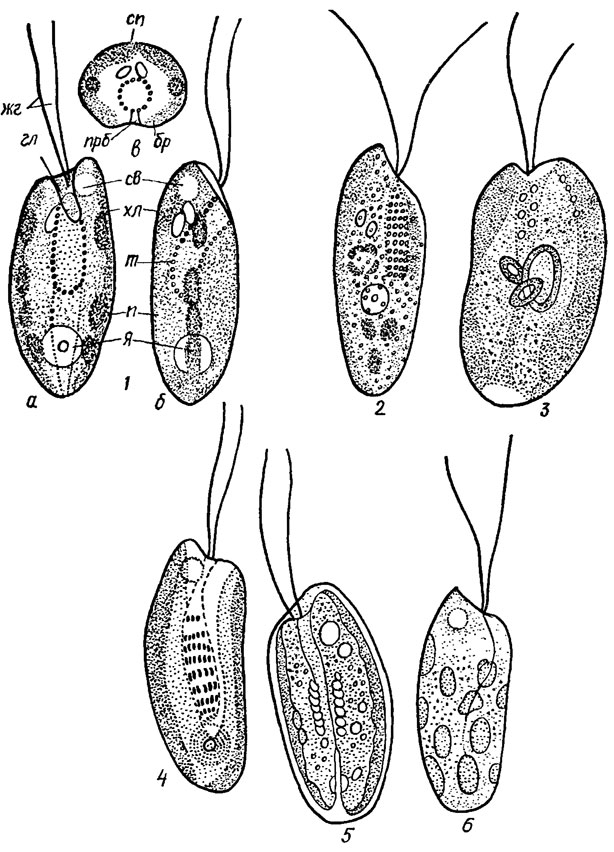
Рис. 12.2. Строение клетки криптомонад: 1 - Cryptomonas sp. (а - вид клетки с брюшной стороны, б - сбоку, в - в поперечном разрезе; сп - спинная сторона клетки, бр - брюшная сторона, прб - продольная бороздка, жг - жгутики, гл - глотка, св - сократительные вакуоли, хл - хлоропласты, т - трихоцисты, п - пиреноиды, я - ядро); 2 - Rhodomonas tenuis Skuja; 3 - Chroomonas nordstedtii Hansg.; 4 - Cryptomonas erosa Ehr.; 5 - Cryptomonas ovata Ehr.; 6 - Cryptochrysis polychrysis Pasch
С помощью электронно-микроскопических исследований у криптофитовых водорослей обнаружена уникальная органелла - нуклеоморф. По своей ультраструктуре она имеет сходство с ядром, находится в перипластидном пространстве, окружена двойной мембраной, пронизанной порами и содержит гранулярный матрикс и несколько плотных гомогенных глобул или палочек [562]. В настоящее время нуклеоморф обнаружен у многих видов криптомонад; у других групп водорослей он не обнаружен.
Предполагают, что наличие нуклеоморфа свидетельствует об эндосимбиотическом происхождении хлоропластов криптофитовых водорослей. Основываясь на присутствии у криптофитовых продукта ассимиляции α-1,4-глюкана, а в составе пигментов фикобилинов (фикоэритрина и фикоциана), некоторые авторы постулируют принадлежность эндосимбионта к красным водорослям [441]. Установлено, что в нуклеоморфе имеется два типа гранул: плотные, которые изредка соединяются в узкие ленты, и более крупные фибриллогранулярные тела, имеющие сходство с ядрышками. Применение специальных красителей помогло обнаружить в нуклеоморфе ядерную кислоту. Ядерноподобные свойства нуклеоморфа, по мнению некоторых авторов [441], подтверждают теорию об эндосимбиотической природе хлоропластов криптомонад. Имеются сведения о том, что при делении ядра нуклеоморф делится в препрофазе вслед за репликацией базального тела, но раньше, чем происходит деление хлоропласта и хлоропласт-эндоплазматического ретикулюма. При делении нуклеоморфа сначала впячивается внутренняя мембрана, затем наружная, а микротрубочки в этот процесс не вовлекаются. Колхицин, который ингибирует ядерное деление, не влияет на деление нуклеоморфа [549].
Хлоропластов 1-2, реже больше; они окрашены в разные цвета от сине-зеленого и оливково-зеленого до желто-бурого, коричневого и темнокрасного. Иногда хлоропласты отсутствуют. В составе пигментов обнаружены хлорофиллы а и с, зеаксантин, каротины, а также фикоциан и фикоэритрин. Пиреноиды окаймленные или голые, чаще они отсутствуют.
Продукт ассимиляции - крахмал, у некоторых представителей масло и хризоламинарин.
Питание фотоавтотрофное или сапрофитное, редко голозойное.
Стигма обнаружена не у всех представителей, она является частью хлоропласта, расположена в передней части под оболочкой и состоит из большого числа пигментных глобул, расположенных параллельными рядами от 1 до 9.
На переднем конце клетки находится одна или несколько сократительных вакуолей, которые изливают свое содержимое в воронку или бороздку.
Размножение у криптофитовых происходит путем продольного деления надвое в подвижном или неподвижном состоянии. При размножении воронка и вакуоли делятся пополам, оба жгутика отходят к одной клетке, а другая образует их заново. Известны цисты и пальмеллевидные состояния. Половой процесс неизвестен.
Цикл развития у криптофитовых происходит двумя основными путями. Обычно эти водоросли вегетируют в подвижном, реже в неподвижном пальмеллевидном состоянии. Пальмеллы округлой формы, состоят из неподвижных клеток, окруженных толстым слоем слизи. В цикле развития некоторых видов (Cyanophora paradoxa Korsch.) известны шаровидные цисты, покрытые толстой слизистой обверткой до 40 мкм в диаметре. Возникают они попарно после деления клеток на 2 дочерние. Перед цистообразованием клетки сбрасывают жгутики, округляются, покрываются первоначально тонкой, бесцветной, гладкой, позже двух контурной окрашенной оболочкой и заполняются многочисленными зернами крахмала. У Cryptomonas frigoris Jav. et Hin. обнаружены сферические цисты с гладкой, бесцветной, бесструктурной оболочкой до 10-15 мкм в диаметре. Возникают они в природных условиях на снегу и существуют длительное время. При наступлении благоприятных условий они прорастают, при этом содержимое каждой цисты делится и образуется 2 дочерние клетки, которые после разрыва оболочки выходят из цисты наружу и покрываются своими оболочками.
Положение криптофитовых в современной классификации растительного мира до сих пор дискутируется. Еще А. Пашер [575] высказывал предположение о их происхождении от хризомонад в качестве боковой ветви, и от криптофитовых выводил перидиниевые. Родство криптофитовых и перидиниевых предполагал и Г. Клебс [508]. Позже А. Пашер [576, 577] включил криптофитовые как самостоятельный отдел в филу Pyrrhophyta. Эту точку зрения поддержали и другие ученые [134, 381, 413]. Однако некоторые авторы считали включение криптофитовых в группу пиррофитовых недостаточно обоснованным [423]. Так, Х. Грехем выделяет криптофитовые из пиррофитовых и рассматривает их как придаток к этому отделу, Ф. Фрич рассматривает криптофитовые как самостоятельный класс, а А. Смит [645] и Б. Фотт относят Cryptophyceae к группе организмов "неопределенного систематического положения". Б. Фотт считает правильным вообще не относить их к растительным филам. Криптофитовые водоросли существенно отличаются от пиррофитовых строением ядра, пигментным составом, что свидетельствует против их близкого филогенетического родства. Учитывая ряд своеобразных и морфологических, и биохимических признаков, считается оправданным выделение криптофитовых в самостоятельный отдел Cryptophyta [120].
Отдел включает 1 класс Cryptophyceae, 1 порядок Cryptomonadales, в состав которого входит 5 семейств: Cryptomonadaceae, Cryptochrysidaceae, Katablepharidaceae, Cyathomonadaceae и Senniaceae.
С экологических позиций криптофитовые водоросли - типичные представители планктона. Изредка они встречаются в иле соленых озер (Cryptomonas salina Wisl., С. stigmatica Wisl.) и в поверхностной пленке ила пресноводных водоемов, среди детрита. Большинство криптофитовых живет в поверхностных, хорошо прогревающихся слоях воды, преимущественно в прибрежной зоне. Они преобладают в небольших стоячих водоемах, лужах, пойменных водоемах, рыбоводных, биологических и технических прудах, бассейнах оранжерей ботанических садов, парков и зоопарков, карт полей фильтрации очистных сооружений различных предприятий. Нередко встречаются в старицах, каналах, болотах, реках, лиманах, озерах, водохранилищах. Виды этих местообитаний обычно вегетируют осенью и даже зимой (Cryptomonas obovata Skuja, Katablepharis ovalis Skuja). Известны виды, живущие и на больших глубинах. Так, Chilomonas thiophila Skuja обнаружен в планктоне озер на глубине 10-14 м. Особый интерес составляют тиофильные виды (Cryptaulaх thiophila Skuja), живущие в озерах на глубине 13-22 м вместе с другими глубоководными видами синезеленых, десмидиевых водорослей и тиофильными бактериями. В сероводородной зоне озер на глубине до 25 м встречается Cryptomonas phaseolus Skuja.
Типичным представителем нейстона является Cryptomonas platyuris Skuja. Очень часто в пробах нейстона встречается Cryptomonas erosa Ehr. Среди криптофитовых встречаются типичные сфагнофилы, живущие в болотах с низкими значениями pH, например, Cryptomonas cylindrica Ehr. (pH 3,5-4,1). Некоторые виды могут развиваться в широком диапазоне pH, например, Chilomonas paramaecium Ehr. (pH 4,1-8,4).
Преимущественное большинство криптофитовых - эвритермные формы. К стенотермным относятся виды рода Rhodomonas Karst. Типичным криофилом является такой вид, как Cryptomonas frigoris Jav. et Hindak, описанный из монокультуры "красного" и "зеленого" снега из Высоких Татр, и Cryptomonas cryophiia из Антарктиды.
Наиболее интересны в экологическом и практическом отношении виды, живущие в сточных водах, загрязненных различными веществами. Они часто достигают здесь массового развития и даже вызывают "цветение" воды. Это главным образом виды рода Cryptomonas: С. caudata Schill., С. compressa Pasch., С. ovata Ehr., C. reflexa (Mars.) Skuja, C. curvata Ehr., C. rufescens Skuja. Особенно часто во всех типах стоков почти в течение всего года в значительном количестве (до 200 млн кл/л) и с большой биомассой (до 600 мг/л) встречается С. ovata. Из других родов в сточных водах зарегистрированы Chilomonas paramaecium, Chroomonas caudata (Geitl.) Skuja. Физико-химические показатели среды, в которой развиваются эти виды, свидетельствуют о их высокой приспособляемости к таким условиям. Некоторые из них могут быть показателями сап ровности. Известны олиго-β-мезосапробы, α-β-мезосапробы и β-мезосапробы из родов Cryptomonas, Chroomonas, Rhodomonas и Cryptochrysis. В биохимическом и физиологическом отношении криптофитовые изучены очень слабо [209].
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'