11. Отдел 2. Динофитовые водоросли - Dinophyta (Dinophyceae, Pyrrhophyta, Pyrrhophycophyta, Peridineae) (З. И. Ветрова)
Динофитовые - преимущественно одноклеточные с монадным типом структуры, но некоторые из них имеют амебоидную, пальмеллоидную, коккоидную или нитчатую структуру. В настоящее время известно около 300 видов, обитающих в пресных и морских водах. Клетки обычно окрашены в темнобурый, красный, иногда в желтый или желто-зеленый цвет, имеются также бесцветные формы.
Индивиды характеризуются разнообразной формой тела, среди которой преобладает шаровидная, эллипсоидная, яйцевидная, реже обратнояйцевидная, грушевидная, обратногрушевидная, некоторые виды имеют форму тела в виде полумесяца или пирамиды. Жгутиковые организмы и стадии отличаются дорсивентральным телом, с бороздками или без них (рис. 11.1). Тело обычно уплощенное, в поперечном сечении эллипсоидное или в виде пластинки. Изредка встречаются виды с неуплощенным округлым телом.
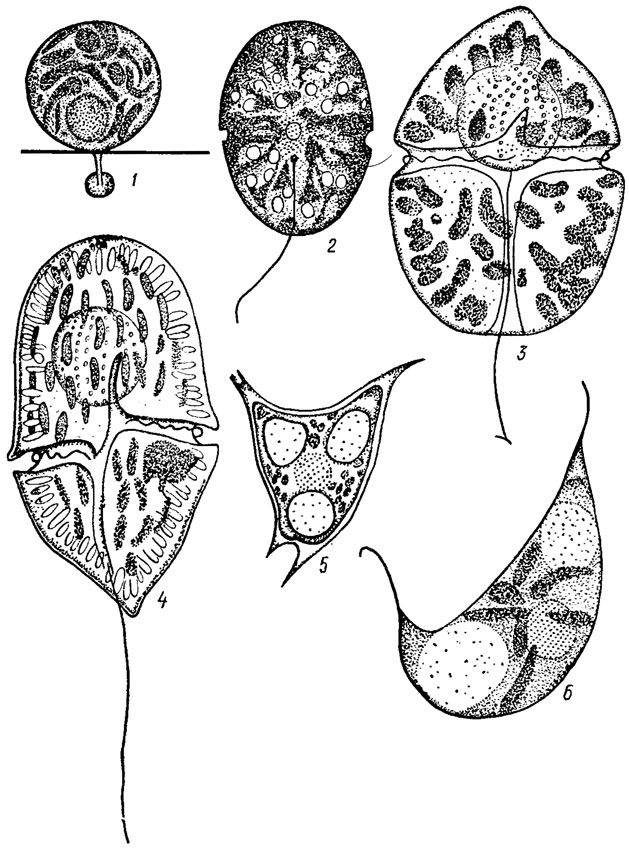
Рис. 11.1. Форма тела динофитовых водорослей: 1 - шаровидная (Stylodinium sphaera Pasch.); 2 - эллипсоидная (Glenodiniun. steinii Lemm.): 3 - яйцевидная (Gymnodinium aeruginosum Stein): 4 - обратно-яйцевидная (G. fusctim (Ehr.) Stein): 5 - тетраэдрическая (Tetradinium javanicum Klebs); 6 - в виде полумесяца (Cystodinium steinii Klebs)
Размеры тела динофитовых колеблются в пределах от 6 (Atelodinium microsporum) до 1500 мкм (Noctiluca miliaris Cienk.). Среди пресноводных представителей наибольших размеров достигают виды рода Ceratium Schrank - до 450 мкм. Максимальных размеров в пределах каждого вида достигают клетки перед делением или образованием цист.
Клеточный покров у наиболее примитивных форм представлен гладким тонким и нежным перипластом (шаровидные формы). Преимущественное большинство динофитовых покрыты текой, часто образующей панцирь (рис. 11.2; 11.3). Он состоит из трех основных частей: верхней - эпивальвы, нижней - гиповальвы и средней - пояска, соединяющего обе части в сплошной панцирь. Эпивальва и гиповальва состоят из пластинок и щитков, соединенных в определенном порядке швами. Верхушка эпивальвы у некоторых видов оканчивается апексом, нижний конец гиповальвы называют антапексом. Некоторые пластинки гиповальвы иногда имеют постоянное число шипов (Peridinium aciculiferum Lemm.; см. рис. 11.2). Внешняя поверхность пластинок гладкая или чаще имеет ноздреватую, штриховатую, точечную скульптуру. Пластинки панциря имеют различную конфигурацию в виде неправильных многоугольников, ромбов, квадратов, треугольников, трапеций и т. д. Число и размещение пластинок, а также наличие шипов относятся к числу важных систематических признаков.
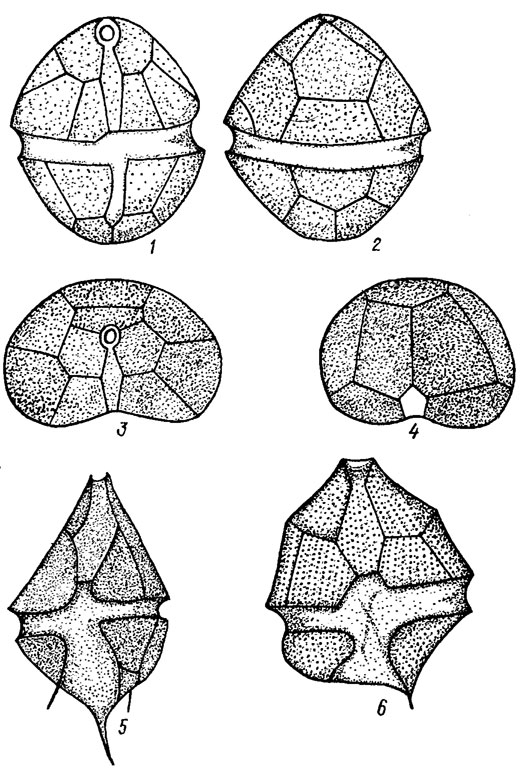
Рис. 11.2. Строение панциря: 1-4 - Peridinium charkowiensis Matv. (1 - со спинной стороны, 2 - с брюшной, 3, 4 - вид сверху); 5 - P. aciculiferum Lemm.; 6 - Р. inconspicuum Lemm
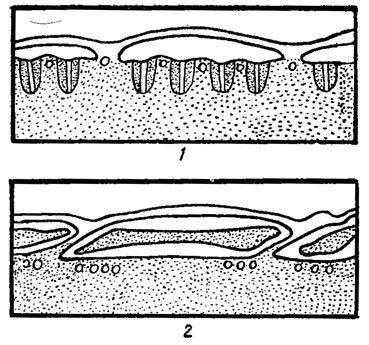
Рис. 11.3. Строение теки у представителей родов Gyrnnodinium Stein (1) и Glenodinium Ehr. (2) (по данным электронной микроскопии)
Швы, соединяющие пластинки панциря, бывают узкие, более или менее широкие, плоские или выпуклые, гладкие или штриховатые. Они представляют собой зону роста панциря, за счет их разрастания увеличивается длина и ширина самого панциря. Молодые клетки имеют простые, узкие швы, старые - широкие и обычно поперечно-штриховатые. На панцире многих видов имеются две бороздки - поперечная и продольная. Поперечная бороздка разделяет панцирь на верхнюю (эпивальву) и нижнюю (гиповальву) части и проходит обычно по экватору, редко она смещена к переднему концу, как у видов рода Amphidinium Clap, et Lachm., или к нижнему, как у видов рода Katodinium (Conr.) Fott. Бороздка полностью описывает дорсальную сторону панциря, соединяясь или не соединяясь концами на вентральной стороне. Поэтому различают кольцевую и спирально завитую поперечную бороздку. Иногда она опоясывает только половину панциря, тогда как вторая часть остается сплошной, не разделенной на верхнюю и нижнюю половины (виды рода Hemidinium Stein). На свободных выступающих краях поперечной бороздки иногда наблюдаются крыловидные поперечно исчерченные перепонки. Продольная бороздка расположена на вентральной стороне панциря, проходит только по гиповальве, редко переходит и на эпивальву, а у видов рода Gonyaulax Dies, доходит до апекса, переходя даже на дорсальную сторону. Левый край продольной бороздки иногда оттянут в крыловидный вырост или оканчивается выступающим на панцире зубцом или шипом.
Монадные динофитовые имеют два неравных по величине жгутика - поперечный и продольный. Поперечный жгутик имеет вид волнистой ленты, выходит из верхней жгутиковой поры и опоясывает клетку влево. Этот жгутик расположен в поперечной бороздке. Продольный - рулевой - жгутик в виде тонкой нити выходит из нижней жгутиковой поры и у большинства видов расположен в продольной бороздке панциря. Он тянется вдоль клетки, выходя за ее пределы, и обеспечивает поступательное движение. Динофитовые с коккоидной, пальмеллоидной и нитчатой структурой имеют жгутиковые зооспоры.
Клетки динофитовых водорослей имеют ядро мезокариотического типа. В ядре может быть одно или несколько ядрышек, которые при делении обычно исчезают. Хромосомы, в отличие от таковых других эукариот, лишены гистонов и состоят из микрофибрилл, содержащих чистую ДНК. Они слабо дифференцированы по длине из-за отсутствии центромеров и вторичных перетяжек, не претерпевают циклических преобразований и постоянно находятся в конденсированном состоянии. Во время митоза ядерная оболочка не исчезает и веретено делении не образуется. У некоторых представителей (Oxyrrhis marina Duj.) ядрышко при делении сохраняется как и у эвгленовых водорослей.
Хлоропласты обычно дисковидные, встречаются удлиненно-эллипсоидные или пластинчатые, лентовидные, вымпеловидные, как правило, многочисленные, редко хлоропластов несколько или даже один, пристенные или радиально расходящиеся от центра к периферии; иногда они расположены в эпивальве или гиповальве (Amphidinium operculatum Clap, et Lachm.). Хлоропласты содержат хлорофиллы а и с, β- и γ-каротин, из ксантофиллов - фукоксантин, диадиноксантин, диатоксантин, диноксантин, перидинин, неоперидинин, пирроксантин.
Пиреноиды наблюдаются очень редко, преимущественно у коккоидных и панцирных форм, хлоропласты расположены радиально. Обычно пиреноид находится в центре хлоропласта и окаймлен крахмальными скорлупками (Amphidinium rhynchocephalum Anissim.)
Стигма расположена за пределами хлоропласта, в основании жгутиков. Состоит из двух рядов пигментных глобул, окруженных двухмембранной оболочкой, и пластинчатого тела, связанного с эндоплазматическим ретикулюмом. Некоторые динофитовые имеют крупную стигму (до 20 мкм длиной и 15 мкм шириной), которая состоит из пигментных глобул, линзовидного тела и ретиноида. Линзовидное тело занимает почти 2/3 стигмы и состоит из группы строго упорядоченных пузырьков. В основании линзы расположен ретиноид. Он представляет чашеподобное образование, состоящее из определенным образом ориентированных фибрилл и зернистого вещества, а также расположенных на самом дне ряда глобул. Благодаря такой организации стигмы свет попадает на линзу, фиксируется и концентрированным пучком посылается через ретиноид на глобулы, т. е. приходит туда в преобразованном виде.
У пресноводных видов имеются мелкие, рассеянные в протопласте вакуоли и крупные. Характерной особенностью динофитовых являются своеобразные вакуоли, называемые пузулами, открывающимися узким каналом у места прикрепления жгутиков. У некоторых представителей обнаружены трихоцисты - палочковидные или булавовидные бесцветные образования, радиально расположенные на периферии протопласта. У некоторых морских видов наблюдаются сложные капсулы, в середине которых содержатся спирально сложенные жалящие нити (Polykrikos Biitschli). Продуктами ассимиляции динофитовых являются крахмал и масло, реже хризоламинарин, гликоген и др.
Размножение у динофитовых водорослей осуществляется в основном делением клеток в подвижном состоянии или зооспорами и апланоспорами. У некоторых видов описан половой процесс, происходящий путем слияния взрослых клеток или жгутиковых клеток и апланоспор. При неблагоприятных условиях у многих панцирных видов образуются толстостенные цисты, по форме напоминающие вегетативные клетки (Ceratium cornutum (Ehr.) Clap. et Lachm. Вначале оболочка у них тонкая, однослойная позже она утолщается, нередко покрывается выростами в виде шипов (Woloszynskia leopoliensis (Wolosz.) Thomp.).
Циклы развития динофитовых водорослей с участием полового процесса считаются наиболее примитивными. Подавляющее большинство динофитовых размножается делением клеток на две части. Клетки паразитических видов делятся на большее число частей. Известно также размножение мелкими амебоидами. Половое размножение наблюдается редко. Половой процесс - изогамия или слабо выраженная анизогамия. В качестве гамет могут функционировать вегетативные клетки или в результате их деления в каждой клетке образуется несколько гамет, часто внешне подобных вегетативным клеткам. Мужские гаметы мельче женских. Перед слиянием гаметы соединяются брюшными сторонами, мужская гамета при этом сбрасывает щитки панциря и ее цитоплазма вливается в цитоплазму женской гаметы. Образовавшаяся зигота остается подвижной (планозигота). Внешне она подобна вегетативной клетке. Таким образом возникает диплоидный организм, который в отличие от гаплоидных индивидов не размножается бесполым путем. Планозигота растет несколько дней и превращается в большую клетку с длинными выростами. При первом делении мейоза возникают ядра с половинным набором хромосом и образуются две жгутиковые клетки с антапикальными и апикальными выростами. Через 2-3 дня происходит второе мейотическое деление, в результате которого образуются гаплоидные вегетативные клетки. Таков цикл развития у Ceratium horridum Gran. Обычно же у динофитовых водорослей наблюдаются циклы развития с преобладанием гаплоидной или диплоидной формы развития. В случае преобладания гаплоидной фазы диплоидная форма развития существует относительно короткий период. Переход зиготы в гипнозиготу (гипноспору) существует не у всех представителей динофитовых. Гипнозиготы возникли у динофитовых не в связи с переходом от гаплоидного состояния к диплоидному, а как форма приспособления к переживанию неблагоприятных условий. Гипнозигота в ряде случаев составляет один из этапов существования диплоидной формы развития [253].
Существуют разные мнения о филогенетических связях динофитовых. Так, Г. Клебс [508] указывал на некоторые черты сходства динофитовых с криптофитовыми. Ф. Шютт [624] предполагал их родство с диатомовыми, а Р. Веттштейн [693] объединял динофитовые, диатомовые и конъюгаты в один отдел (ствол) Zygophyta. А. Пашер [577] считал динофитовые водоросли рода Peridinium и криптофитовые родственными группами и объединил их в один ствол Pyrrhophyta, разделив его на три отдела: Cryptophyceae, Desmocontae, Dinophyceae [575, 579]. M. Шадефо [371] включал в подотдел Pyrrophyta классы Cryptophyceae, Dinophyceae, Raphidophyceae и Euglenophyceae, основываясь на их сходстве по некоторым признакам. Как показал анализ литературных данных, наиболее распространен взгляд на родство динофитовых с криптофитовыми. Основанием для этого является сходная окраска хлоропластов, наличие в клетках крахмала как запасного продукта и строение монадных форм, имеющих два неодинаковых жгутика и продольный брюшной желобок. Однако, что касается окраски хлоропластов, то, как отмечал Д. К. Зеров [120], криптофитовые резко отличаются от динофитовых наличием фикобилинов, иным составом каротинов и ксантофиллов. Монадные формы криптофитовых имеют ряд особенностей в строении, отличающих их от подвижных клеток динофитовых. Это дало основание ряду авторов исключить криптофитовые из состава динофитовых и рассматривать последние в качестве самостоятельного обособленного отдела Dinophyta.
В принятой нами классификации отдел Dinophyta делится на два класса: Desmophyceae и Dinophyceae [120, 306, 414, 645]. Класс Desmophyceae включает 4 порядка: Desmomastigales, Prorocentrales, Dinophysidiales, Desmocapsales. Спорное положение в системе Desmophyceae занимает порядок Dinophysidiales, который некоторые авторы переносят в класс Dinophyceae, поскольку у представителей этого порядка имеются поперечные и продольные бороздки, оболочка состоит из отдельных пластинок, место прикрепления жгутиков несколько сдвинуто на брюшную сторону. Но наряду с этими признаками у Dinophysidiales имеется характерный для панцирных Desmophyceae продольный шов, разделяющий оболочку в сагиттальном направлении на две половины. Д. К. Зеров полагает, что развитие сходных черт у Dinophysidiales и Peridiniales является конвергентным приспособлением к сходным условиям существования и Dinophysidiales развились из панцирных форм типа Prorocentrales, имеющих такой же сагиттальный шов. По Д. К. Зерову [120], классы Desmophyceae и Dinophyceae представляют две параллельные ветви эволюции.
Динофитовые происходят от голых монадных форм со своеобразным ядром, двумя жгутиками и наличием определенных пигментов. В классе Desmophyceae наиболее примитивный порядок Desmomastigales, а в классе Dinophyceae - Gymnodiniales. Из форм, сходных с современными Gymnodiniales, развились покрытые оболочкой, подвижные в вегетативном состоянии Peridiniales, амебоидные Dinamoebidiales, пальмеллоидные Gloeodiniales и паразитные Blastodiniales. Нитчатые Dinotrichales, очевидно, развились из коккоидных форм. Д. К. Зеров считает, что основным для такого построения филогенетических отношений в классе Dinophyceae является тот факт, что представители всех порядков этого класса, как бы они не отличались в вегетативной стадии, имеют зооспоры, сходные по строению с Gymnodinium.
Экологические особенности, распространение и роль в природе динофитовых изучены еще недостаточно. Большинство представителей с монадной структурой относятся к планктонным организмам, обитающим обычно в прибрежье водоемов, в зарослях макрофитов, среди скоплений нитчатых водорослей. Только некоторые виды достигают массового развития в поверхностных слоях ила (Amphidinium rhynchocephalum). Значительно меньше динофитовых среди эпифитов и типичных бентосных форм. К широко распространенным относятся немногие виды - это Peridinium cinctum (О. F. М.) Ehr., Р. bipes Stein, P. willei Huitf-Kaas, Ceratium hirundinella (O. F. M.) Schrank, C. cornutum (Ehr.) Clap, et Lachm. Известны пресноводные, солоноватоводные и морские динофитовые. Значительную роль в распределении динофитовых играют температурный и световой факторы. Максимальное развитие большинства видов связано с летним периодом, однако есть и такие холодолюбивые представители (Реridinium aciculiferum Lemm., Gymnodinium macronucleum Litv., G. helveticum Penard и др.), массовое развитие которых наблюдается лишь в зимний период. Встречаются и типичные криофилы (Gyrodinium pascheri (Such.) Schil.), вегетация которых происходит в снегу. Большое значение в распределении динофитовых имеют хлор, кальций, органические вещества и pH среды. Некоторые виды предпочитают щелочные воды (Glenodinium beroiinense (Lemm.) Lind., G. pygmaeum (Lind.) Schill., Peridinium aciculiferum, Ceratium cornutum). Большинство видов не выносят загрязненных вод (олигосапробы) [209].
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'