9a. 4. Класс 3. Гормогониевые - Hormogoniophyceae (Nostochineae hormogoneae, Hormogoneae) (Н. В. Кондратьева)
(Nostochineae hormogoneae [678], Hormogoneae (см. [357, 445]).)
Класс Hormogoniophyceae [651a] является самым крупным классом Сyаnophyta, включающим большую часть его видов, характеризующихся нитчатой (трихомальной) формой строения тела особей и способностью к образованию гормогониев, т. е. специальных фрагментов трихомов, способных к активному (произвольному) движению и прорастанию в новые особи.
Индивиды многоклеточные, "простые" или колониальные (с многоклеточными цианоидами). "Простые" индивиды содержат один трихом (см. рис. 1.1) и наиболее соответствуют термину "индивид" в смысле "особь", "организм". Колониальные индивиды обычно возникают из одного "простого" и имеют признаки, характеризующие их как целое (наличие общего "скелета" в виде общих влагалищ, т. н. перидерма (см. рис. 1.9), особых способов размножения, например почкования, и по сути соответствуют эвкалониям (настоящим колониям), т. е. колониям в узком понимании (рис. 9.32, 1; см. рис. 1.9; 3.1, 10, 11, 14). К колониям в широком смысле слова наряду с колониальными индивидами (колониальными организмами) относятся также колонии индивидов, например сплетения нитей осциллаториевых водорослей. В отличие от колониальных индивидов колонии индивидов - образования менее целостные. Они нередко возникают из многих, ранее обособленных особей и часто соответствуют поселениям индивидов. Образуются, например, на агаризованной среде в месте внесения в нее исходного инокулята.
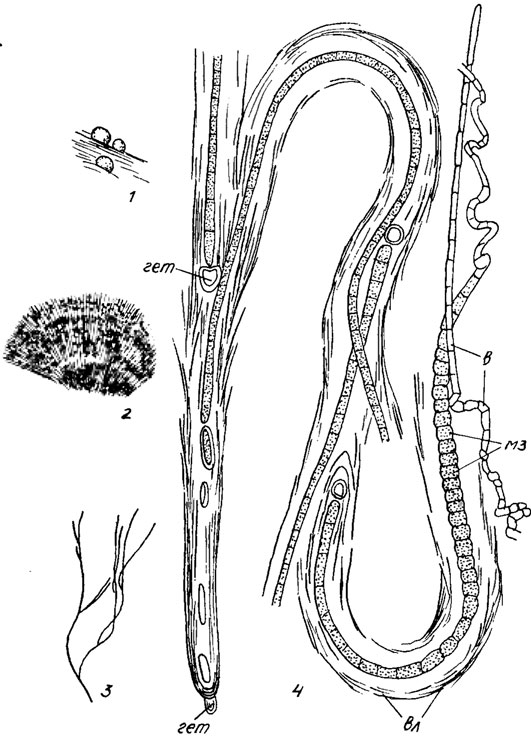
Рис. 9.32. Rivularia haematites (D. C.) Ag.: 1 - общий вид трех 'сложных' колоний; 2 - часть среза через 'сложную' колонию; 3 - общий вид 'сложного' цианоида; 4 - часть 'сложного' цианоида, отдельные участки которого соответствуют 'простым' многоклеточным цианоидам, сходным с 'простыми' индивидами; гет - гетероцисты; в - волосик; мз - меристемальная зона; вл - влагалище
Нити гормогониевых водорослей могут соответствовать "простым" и колониальным индивидам, быть разветвленными или неразветвленными. Ветвление нитей бывает настоящим и ложным. Настоящим ветвлением, называют такое, при котором наблюдается ветвление трихомов (рис. 9.33). При ложном ветвлении нитей ветвятся только влагалища (см. рис. 1. 5, 5, 10 и 9.32)*.
* (Влагалища, окружающие один трихом или ряд последовательно расположенных трихомов, называют индивидуальными влагалищами. Влагалища вокруг группы относительно параллельных трихомов именуют общими влагалищами (см. рис. 9.2).)
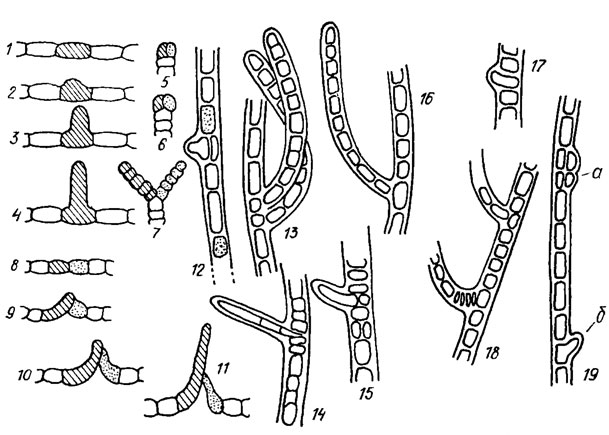
Рис. 9.33. Типы ветвления нитей гормогониевых водорослей: 1-4 - настоящее боковое ветвление; 5-7 - дихотомические ветвления; 8-11 - У-образное ветвление у Mastigocladus laminosus Cohn; 12-19 - настоящее боковое ветвление у Hapalosiphon fontinalis (Ag.) Born. em. Elenk. (12, 15, 19, a - ветвлению предшествует продольное деление клетки, в результате которого в трихомах местами наблюдается двурядное расположение клеток; 19, б - ветвь возникает в результате недоведенного до конца почкования вегетативной клетки)
Существует несколько типов ложного ветвления. У форм с индивидуальными влагалищами наблюдается плектонемоидное ветвление нитей (см. рис. 1.5, 10), происходящее в результате фрагментации трихомов и дальнейшего прорастания фрагментов in situ (на месте). Концы прорастающих фрагментов прорывают старые влагалища, окружаются новыми и превращаются в боковые ветви. Если прорастает и превращается в ветвь только один из смежных концов расположенных рядом фрагментов трихома, возникает одинарное ветвление. Если в ветви преобразуются оба смежных конца двух соседних фрагментов, возникает двойное ветвление. У форм с общими влагалищами наблюдается схизотриксоидное ветвление, при его развитии трихомы или их фрагменты выдвигаются из концов влагалищ, отклоняются друг от друга и окружаются новыми влагалищами. Иногда на концах нитей с общими влагалищами образуется целый пучок ветвей. При схизотриксоидном ложном ветвлении ветви по своему происхождению обычно терминальные (боковыми они бывают вторично). При плектонемоидном - первично боковые.
Трихомы бывают одно-, дву- и многорядными, неразветвленными или разветвленными, гомоцитными (состоящими из относительно сходных, не дифференцированных по форме и функциям клеток) и гетероцитными - состоящими из клеток, неодинаковых по форме, функциям, а иногда и локализации. О форме клеток трихомальных особей см. раздел 1.2.1 (см. рис. 1.2; 1.4). Напомним только, что те клетки гетероцитных трихомов, которые внешне сходны с клетками гомоцитных трихомов, называются вегетативными, резко отличающиеся от них - особыми.
К числу особых клеток принадлежат гетероцисты и акинеты. И те и другие возникают из вегетативных клеток. При образовании гетероцист содержимое вегетативной клетки становится гомогенным, более бледным (до бесцветного) или желтоватым. Оболочка исходной для них клетки утолщается, становится двух контурной; на полюсах появляются поры и другие полярные структуры (пробки, полярные зернышки). Двупоровые гетероцисты, возникшие из срединных вегетативных клеток, называются интеркалярными. Однопоровые, образовавшиеся из конечных вегетативных клеток трихома, - терминальными. Терминальные гетероцисты, расположенные в основе трихома, носят название базальных, находящиеся на концах боковых ветвей трихома - латеральных (см. рис. 1.8).
Акинеты (споры, точнее покоящиеся споры, покоящиеся клетки) - это особые одноклеточные образования, выполняющие функцию сохранения жизни в неблагоприятных условиях, а в тех случаях, когда они образуются на особях в количестве большем, чем одна - то и функцию размножения. Оболочка акинет утолщенная, нередко с украшениями (см. рис. 1.7).
Рост особей происходит за счет деления вегетативных клеток (или сходных с ними клеток гомоцитных трихомов), осуществляющегося обычно в плоскости, поперечной к продольной оси трихома, что ведет к линейному расположению клеток. В случае продольного деления могут образоваться многорядные нити и нити с настоящим ветвлением (рис. 9.34; см. рис. 1.14, 4). Деление клеток трихома бывает диффузным или локальным, в определенных (интеркалярных, апикальных или базальных) зонах трихома (рис. 1.14). Тогда говорят о наличии меристемальных зон.
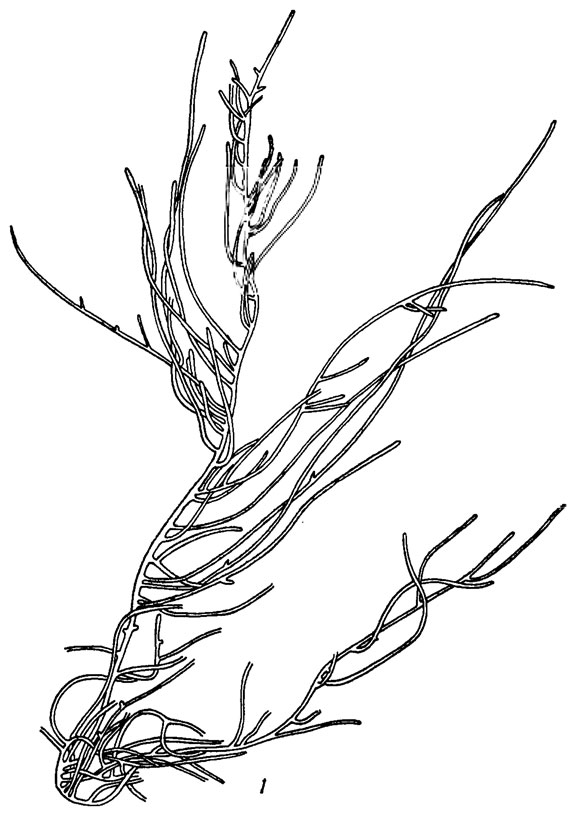
Рис. 9.34. Hapalosiphon fontinalis (Ag.) Born. em. Elenk.: 1 - общий вид нити
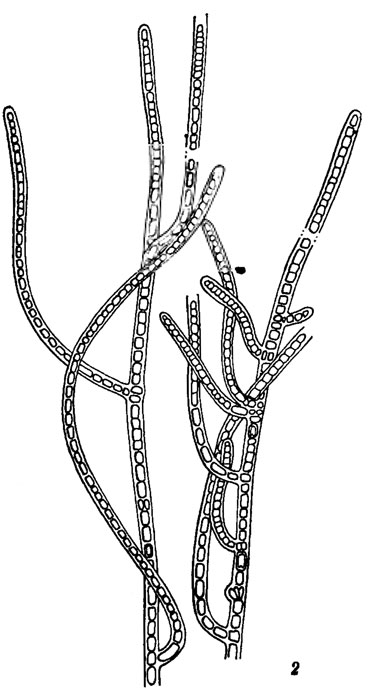
Рис. 9.34. Hapalosiphon fontinalis (Ag.) Born. em. Elenk.: 2 - части нитей при большем увеличении
Размножение у гормогониевых водорослей, как правило, вегетативное (случайной фрагментацией, гормогониями, акинетами и др.). Собственно бесполое размножение не характерно. Эндоспоры обнаружены только у отдельных представителей. Экзоспоры не найдены. Половое размножение и жгутиковые стадии отсутствуют.
Наиболее характерно для гормогониевых водорослей размножение с помощью гормогониев, которые у многих форм образуются по всей длине трихома путем распада его на внешне невидоизмененные части. Однако нередко гормогонии образуются в определенных частях нити (например, на верхушке ветвей; рис. 9.35) и представляют собою относительно дифференцированные образования. Их возникновению нередко предшествует деление клеток материнского трихома. У гормогониевых водорослей с влагалищами гормогонии могут освобождаться путем активного выползания. Различают первичные гормогонии, образующиеся в результате прорастания акинет (см. рис. 3.1, 7) и вторичные гормогонии, возникающие путем фрагментации трихомов.
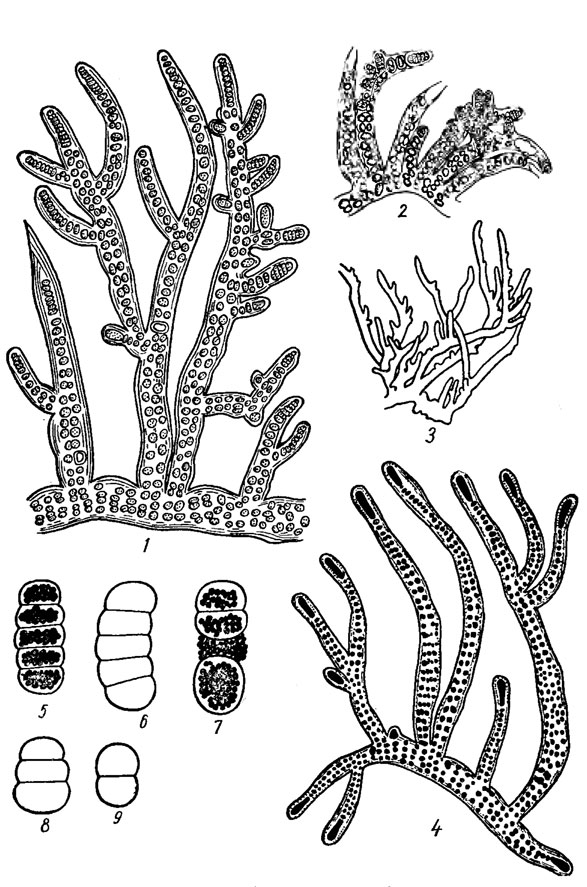
Рис. 9.35. Stigonema minutum (Ag.) Hass. em. Elenk.: 1-3, 5-9 - f. minutum (1, 2 - части нитей с гормогониями; 3 - контурное изображение части разветвленной нити-кустика при меньшем увеличении; 5-9 - гормогонии при большем увеличении); 4 - f. turfaceum (Cooke) Elenk
Сведения о циклах развития гормогонневых водорослей содержатся в разделе 3.1; данные о внутривидовой изменчивости - в разделе 8.2. Здесь напомним только, что представители гормогониевых водорослей нередко имеют довольно сложный цикл развития, в процессе которого наблюдается ряд морфологически неодинаковых стадий, на которых особи могут быть сходными с представителями других родов, и что для Hormogoniophyceae свойственна поливариантность (многовариантность) развития, связанная с их способностью размножаться на промежуточных этапах онтогенеза и зависящая от условий окружающей среды (см. рис. 3.1). В жизненном цикле гормогониевых водорослей нередко наблюдается смена уровня индивидуальности особей. Классификация жизненных циклов Hormogoniophyceae еще не разработана. Их можно группировать по разным признакам: полноте прохождения этапов (полные или неполные), их числу, темпам прохождения, разнообразию и характеру размножения, его повторности и др.
Основным типом онтоморфогенеза гормогониевых водорослей (из одноклеточных или малоклеточных зачатков) является тот, при котором водоросль на начальных этапах развития закономерно проходит осциллаторие-подобное (т. е. напоминающее особи рода Oscillatoria Vauch.), нередко активно подвижное состояние (status oscillatorioideus; см. рис. 3.1, 1, 2). В таком состоянии пребывают, например, первичные и вторичные гормогонии.
Судя по данным сравнительно-онтоморфологического анализа, проведенного с учетом других сведений [141], первичные трихомальные синезеленые водоросли (произошедшие, вероятно, из свободных первичных одноклеточных форм в результате их незавершенного деления), по-видимому, были мало клеточными, однорядными, более или менее цилиндрическими, симметричными по отношению к поперечной плоскости, не имели четко выраженных влагалищ и, не исключено, с самого начала были способны к скользящему движению вдоль субстрата. От таких исходных многоклеточных форм могло возникнуть все разнообразие современных гормогониевых водорослей. Один из вероятных путей эволюции (в результате сравнительно равномерного совершенствования всех клеток первичного трихома и вследствие повышения его целостности, осуществлявшегося за счет изменений на субклеточном уровне) привел к становлению современного порядка осциллаториевых (Oscillatoriales) водорослей. Два других основных пути были направлены главным образом в сторону разделения функций между клетками первичного трихома и привели к современным потоковым (Nostocales) и стигонемовым (Stigonematales) водорослям. Вероятны и другие варианты путей эволюционного развития гормогониевых. Многие авторы, например, полагают, что порядки Oscillatoriales и Nostocales возникли в результате регрессивного развития каких-то представителей порядка Stigonematales, в свою очередь произошедших от тех или иных таксонов класса Chamaesiphonophyceae. В связи с этим еще и сейчас существуют принципиально различающиеся классификации гормогониевых [141].
Ниже описаны только три основных порядка класса Hormogoniophyceae: Oscillatoriales, Nostocales и Stigonematales.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'