9а. 1. Общая характеристика Cyanophyta (Н. В. Кондратьева)
Отдел Cyanophyta Schussnig [622] (включает около 2 тыс. видов, широко распространенных в разнообразных водных и вневодных биотопах, их индивиды одноклеточные или многоклеточные, "простые" или колониальные (с одноклеточными или многоклеточными* цианоидами; см. рис. 9.2 и 9.3). Они могут быть прикрепленными и неприкрепленными к субстрату, неподвижными или способными к скользящему движению, однако жгутиков и ресничек никогда не образуют. Особи Cyanophyta обычно микроскопические, но колониальные индивиды у ряда видов (например, у Nostoc pruniforme Ag.) могут измеряться сантиметрами. Индивиды очень часто образуют разнообразные соединения - колонии индивидов, иногда занимающие большие пространства, и продуцируют значительное количество слизи, нередко заметно влияющей на форму и общий габитус колоний.
* (Термин "многоклеточный" по отношению к Cyanophyta означает, что тело особей состоит из нескольких или многих клеток, между которыми существует плазматическая связь.)
Индивиды и колонии индивидов синезеленых водорослей обычно имеют специфический синезеленый цвет. Однако их окраска может сильно варьировать в зависимости от комбинации пигментов - быть почти зеленой, оливковой, желтовато-зеленой и др. Для одноклеточных Cyanophyta характерна коккоидная форма строения тела. Для многоклеточных - нитчатая (трихомальная), реже разнонитчатая [141]*. Очень редко наблюдается определенная тенденция к пластинчатому или объемному расположению клеток. Жгутиковая (монадная) форма строения тела отсутствует. Обязательной составной частью тела особей гормогониевых водорослей является трихом - нитевидное (неразветвленное или разветвленное) образование, состоящее из одного, реже двух - многих рядов физиологически взаимосвязанных клеток. Нитевидными у Cyanophyta бывают и колонии одноклеточных форм. Плазматическая взаимосвязь между клетками в этом случае отсутствует (рис. 9.1, 2). Понятие "нить" (у представителей крупнейшего класса отдела Суаnоphyta - гормогониевых водорослей) шире понятия "трихом". Только в простейшем случае нить гормогониевых представлена одним трихомом (см. рис. 1.1). Чаще (рис. 9.2, см. рис. 1.5) в ее состав входит помимо трихома (или группы трихомов), также влагалище (т. е. слизистый чехол вокруг трихома), а нередко и другие образования (гетероцисты, акинеты и т. п.). В связи с этим нитчатую форму строения тела гормогониевых водорослей лучше называть трихомальной. У одноклеточных Cyanophyta описанных выше нитей, естественно, нет, но нитчатую форму нередко имеют колониальные индивиды, которые в этом случае часто тоже называют нитями.
* (В разделе 1.3 вместо термина "форма строения тела" употреблен термин "тип структуры", имеющий несколько иной смысл, хотя бы потому, что определяется на основании изучения строения тела только взрослых особей. Формы строения тела синезеленых водорослей устанавливаются безотносительно возраста индивидов, в связи с чем одна и та же особь на разных этапах своего развития (или при разных условиях существования) может характеризоваться разной формой строения тела [42, вып. 1, ч. 1; 141]. По сути "форма строения тела" у синезеленых водорослей рассматривается как особый интегральный признак (свойство) индивида. Основные формы строения тела взрослых особей Cyanophyta соответствуют свойствам "основных типов структуры" в смысле, принятом автором раздела 1.3.)
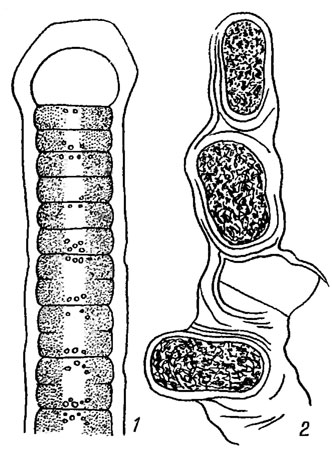
Рис. 9.1. Части нитей гормогониевой водоросли Tolypothrix Kütz. (1) и хамесифоновой Hyella Born, et Flah. (2)
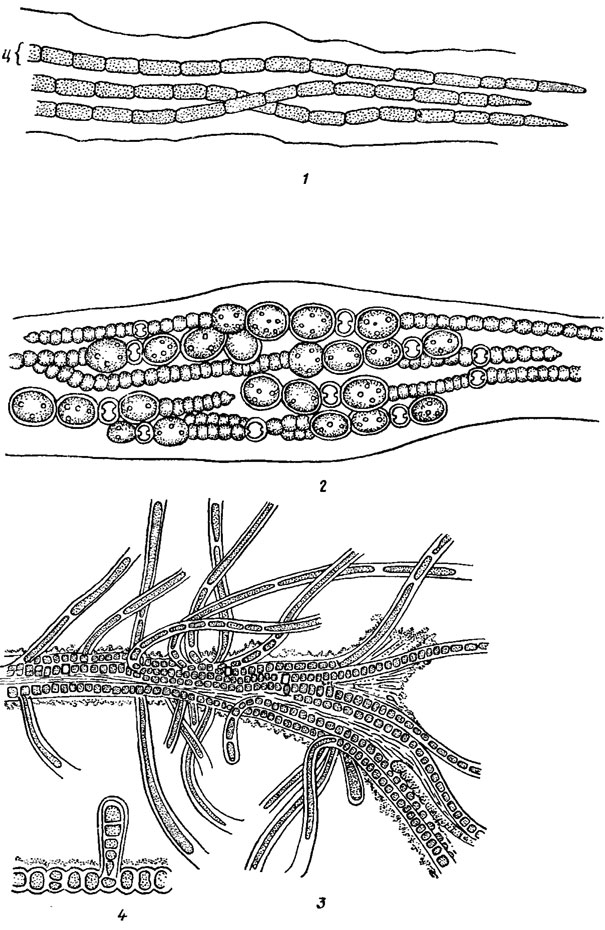
Рис. 9.2. Параллельные формы строения тела (нити с общими влагалищами) у представителей разных порядков: 1 - Microcoleus tenerrimus Gom. (Oscillatoriales; ц - цианоид); 2 - Anabaena solicola Kondrat. (Nostocales); 3, 4 - Fischerellopsis harrissii Fritsch (Stigonematales)
В пределах упомянутых выше основных форм строения тела особей синезеленых водорослей можно выделить подформы. Например, в пределах трихомальной формы строения тела можно различать подформу гомоцитную (клетки не дифференцированы по форме и функции), гетероцитную с неразветвленными трихомами и гетероцитную с разветвленными трихомами (см. рис. 1.5). У представителей разных таксонов Cyanophyta нередко наблюдаются параллельные формы строения тела (см. рис. 1.5; 9.2). Морфологически наиболее прогрессивной формой строения тела особей синезеленых водорослей является та, которая характеризуется разветвленными многорядными гетероцитными трихомами, нарастающими с помощью специальной верхушечной клетки (см. рис. 1.14, 4) и имеющие тенденцию к объемному (или пластинчатому) расположению клеток.
Форма клеток синезеленых водорослей описана в разделах 1.2.1, 9а.2 и 9а.3. Тонкое строение клеток (и их размеры) - в разделе 1.2.1. Здесь напомним только, что настоящих ядер, хлоропластов и вакуолей, выполненных клеточным соком, в клетках Cyanophyta нет. Нередко встречаются газовые вакуоли. Оболочка клеток многослойная, с внутренним пептидогликановым (муреиаовым) слоем. Из хлорофиллов имеется только хлорофилл a. Хлорофилла b и бактериохлорофилла нет. Наличны фикобилипротеиды и каротиноиды. Продуктом ассимиляции является гликогеноподобный полисахарид. Крахмал не образуется.
Клетки и одноклеточные особи Cyanophyta размножаются равным, реже неравным делением. При этом внутренние слои оболочки клетки вростают вовнутрь клетки (подобно закрывающейся ирисовой диафрагме; см. рис. 1.6; 1.12). В отдельных случаях наблюдается множественное деление содержимого. Митоза и мейоза нет. Размножение особей вегетативное, реже собственно бесполое (экзо- и эндоспорами). У ряда представителей образуются покоящиеся споры (акинеты). Типичного полового процесса нет, но обнаружены парасексуальные процессы [42, вып. 1, ч. 1, с. 56], т. е. явления, при которых происходит только частичное объединение геномов разных клеток [96, с. 127], например, трансформация.
Циклы развития синезеленых водорослей кратко охарактеризованы в разделе 3.1.
Внутривидовая изменчивость наблюдается на всех уровнях организации (в том числе на популяционном; см. разделы 1.1 и 8.2) и касается не только зрелых особей, но и особей, находящихся на промежуточных возрастных этапах (что следует учитывать при идентификации синезеленых водорослей). С позиций уровневого подхода уже разработана и апробирована первая классификация внутривидового фенотипического, разнообразия Cyanophyta [53].
Что же касается сходства синезеленых водорослей с другими организмами, то наиболее близки к ним прокариотические зеленые водоросли, которые, подобно Cyanophyta, характеризуются оксигенным фотосинтезом при прокариотическом строении клетки, но отличаются от них наличием хлорофилла b и отсутствием фикобилипротеидов), а также фотосинтезирующие бактерии (фотобактерии - Photobacteriobionta) и группа бесцветных прокариотических микроорганизмов спорного систематического положения. От фотобактерий синезеленые водоросли принципиально отличаются оксигенным фотосинтезом, осуществляющимся с помощью иного комплекса пигментов* и протекающего в аэробных условиях. Показано, однако, что при определенных условиях некоторые Cyanophyta способны переходить к фотосинтезу, сходному с бактериальным. Бесцветные прокариотические микроорганизмы спорного систематического положения (в частности представители семейств Beggiatoaceae Mig. и Vitreoscillaceae Pringsh.; см., например [42, вып. 1, ч. 2; 651a]) внешне очень сходны с представителями некоторых таксонов Cyanophyta, но не имеют ассимиляционных пигментов. Часть из них принадлежит к царству бактерий (Bacteria), но некоторые могут оказаться вторично бесцветными синезелеными водорослями.
* (У фотобактерий типичного хлорофилла нет, но имеется бактериохлорофилл и бактериовиридин [139].)
Вопрос о сходстве синезеленых водорослей с бактериями издавна привлекал внимание исследователей. В настоящее время известны как общие для них признаки, так и признаки их отличающие. Однако вряд ли правильно решать вопрос об уровне обособленности Cyanophyta от Bacteria путем формального подсчета признаков сходства и различия. Правильнее обсуждать его с позиций определенных методологических подходов и, прежде всего, взглядов о ходе развития жизни на Земле и значимости появления конкретных свойств у организмов для развития всей ее биосферы. С этой точки зрения уже только одно наличие у Cyanophyta оксигенного фотосинтеза дает основания для обособления их от бактерий, тогда как все известные признаки сходства этих организмов с бактериями, в том числе прокариотическое строение клетки, может рассматриваться как результат их принадлежности к одному надцарству (Procaryota). В связи с этим предложенное бактериологами переименование синезеленых водорослей (Cyanophyta) в цианобактерии (Cyanobacteriales) нельзя считать оправданным и целесообразным [142, 511], тем более, что термин "водоросли" (как и термин "микроорганизмы") не является таксономическим. И совсем уж неправильным, и даже вредным для развития науки, следует считать предложение подчинить номенклатуру Cyanophyta правилам "Международного кодекса номенклатуры бактерий" [211]*, представляющего собой по сути кодекс номенклатуры культивируемых бактерий, тогда как Cyanophyta уже более столетия изучают главным образом по материалам, собранным непосредственно в природе. Принятие этого предложения внесет в систематику Cyanophyta дополнительную путаницу, усложнит и без того трудную идентификацию их таксонов. Правильнее общими усилиями ботаников-альгологов и бактериологов разработать, как это сделано в отношении высших растений (см. [212]) Международный кодекс номенклатуры культивируемых микроскопических, и в том числе синезеленых, водорослей, который давал бы дополнительные рекомендации относительно наименования их культивируемых форм. Но в целом, по крайней мере при современном уровне знаний, номенклатуру Cyanophyta следует по-прежнему подчинять действию "Международного кодекса ботанической номенклатуры" [42, вып. 1, ч. 1]. Содержание сущности дискуссии по вопросам номенклатуры Cyanophyta освещено в опубликованных нами ранее статьях [142, 511], а также в работах других авторов.
* (Сейчас номенклатура Cyanophyta подчиняется правилам "Международного кодекса ботанической номенклатуры" [493].)
Среди Eucaryota определенные черты сходства с Cyanophyta имеют мезокариотические водоросли (отсутствие связи ДНК с гистонами, отсутствие экваториальной пластинки и др.), к которым принадлежат динофитовые. Однако у них, в отличие от прокариот, имеется ядро, окруженное оболочкой. Синезеленые водоросли сравнивают также с хлоропластами растений, которые, согласно симбиогенетической (экзогенной, эндосимбиотической) теории могли возникнуть в результате внедрения одноклеточных прокариотических водорослей в клетку гетеротрофного организма-хозяина (см. раздел 8.5). В связи с этим Cyanophyta являются ценным модельным объектом для решения вопроса о путях становления растений, тем более, что они были первыми в истории Земли организмами, перешедшими к оксигенному фотосинтезу.
Вопрос о происхождении отдела Cyanophyta до сих пор не решен. Наиболее вероятным является взгляд о дивергентном происхождении бактерий и синезеленых водорослей от неизвестных нам первичных прокариотических форм и более позднем историческом становлении отдела Cyanophyta по сравнению с бактериями. Считают, что первыми (еще в архее) возникли одноклеточные формы синезеленых водорослей, из которых (вероятно в результате их незавершенного деления) образовались трихомальные формы. Общепринят взгляд, согласно которому синезеленые водоросли достигли современного уровня морфологической дифференцированности еще в далекие геологические времена и дошли до наших дней, не претерпев принципиальных морфологических преобразований.
В связи с тем что возникновение многих таксонов Cyanophyta произошло еще в глубине архея, т. е. в ранний период становления жизни, не оставивший достаточного количества ископаемых остатков, для установления родственных связей в пределах отдела Cyanophyta палеонтологические методы почти не применимы. Данные, полученные с помощью физиолого-биохимических методов, для филогенетических обобщений еще недостаточны. Наиболее перспективно в смысле познания вероятных путей эволюционного развития Cyanophyta на данном этапе остается использование сравнительно-морфологического анализа с учетом особенностей онтоморфогенеза и условий их существования в далекие времена. С этой точки зрения можно принять что исходные одноклеточные Cyanophyta, не имеющие утолщенных клеточных покровов, дали начало как современным одноклеточным (прикрепленным и не прикрепленным к субстрату), так и нитчатым (трихомальным) формам синезеленых водорослей.
Все современные формы отдела синезеленых водорослей теоретически (исходя главным образом из существующих представлений об основных направлениях их морфологического прогресса) могут быть сгруппированы в два, три или четыре класса*. Если принять представление о трех основных путях эволюционного развития синезеленых водорослей от исходных коккоидных одноклеточных форм, то можно согласиться в выделением в пределах Cyanophyta трех классов: Chroococcophyceae - хроококковые водоросли, Chamaesiphonophyceae - хамесифоновые водоросли и Hormogoniophyceae - гормогониевые водоросли. Характеристика этих классов приведена в разделах 9а.2 - 9а.4.
* (Многие авторы, однако, рассматривают синезеленые водоросли не как отдел, но как таксон более низкого ранга, и в связи с этим делят его не на классы, но непосредственно на порядки или семейства.)
Представители Cyanophyta играют существенную роль в природе и немаловажны в хозяйственном отношении (см. раздел 6). Благодаря способности к усвоению свободного азота атмосферы при оксигенном фотосинтезе и значительной устойчивости к воздействию неблагоприятных факторов многие из них вегетируют в условиях, непригодных для развития эукариот. Они селятся на бесплодных субстратах (голых скалах, продуктах извержения вулканов и пр.) и принимают участие в процессе формирования первичных почв. Нередки случаи массового развития этих организмов (на такырах, в щелочных термальных источниках, толще воды многих водоемов и др.). В результате их жизнедеятельности заметно изменяется химизм окружающей среды, что сказывается на функционировании экосистем. Биомасса, образованная планктонными Cyanophyta в период интенсивного "цветения" воды (см. раздел 6.5), может составлять 80-90% биомассы всех водорослей. Однако интенсивное развитие синезеленых водорослей наблюдается далеко не во всех современных биотопах. В развитых фитоценозах они уступают высшим растениям, а при определенных условиях даже отсутствуют или почти отсутствуют (при pH менее 4-5, в толще воды быстрых горных рек и др.).
Известны синезеленые водоросли, живущие в симбиозе с другими организмами. В частности они являются компонентами слоевища многих лишайников, сожительствуют с высшими растениями, например, с водным папоротником Azolla и др.
Синезеленые водоросли принадлежат к числу древнейших организмов, их ископаемые остатки и продукты жизнедеятельности обнаружены в породах, образовавшихся 2, 3 (а может быть 2,7-3,2) млрд лет назад, т. е. еще в архейскую эру. Полагают, что первые на Земле (докембрийские) экосистемы состояли только из прокариотических организмов, в том числе Cyanophyta. Говорят даже о "веке синезеленых водорослей". Интенсивное развитие синезеленых водорослей имело громадное значение для развития жизни на Земле, и не только из-за накопления ими органического вещества, но и в связи с обогащением первичной атмосферы кислородом. Существенную роль синезеленые водоросли сыграли и в создании известковых пород. Способность отлагать известь свойственна также ряду современных видов синезеленых водорослей. Говорят о "современных аналогах строматолитов". Некоторые Cyanophyta, напротив, способны к разрушению горных пород.
Хозяйственное значение Cyanophyta связано со съедобностью некоторых их представителей (Nostoc pruniforme Ag., Spirulina piatensis (Nordst.) Geitl. и др.), возможностью использования азотфиксирующих форм для повышения плодородия почв (по крайней мере в районах орошаемого земледелия)*, необходимостью предотвращения чрезмерного "цветения" воды в водоемах, нередко обусловливаемого токсическими формами Cyanophyta, такими как Microcystis aeruginosa Kütz. em. Elenk. (рис. 9.3), Anabaena flos-aquae Breb. (рис. 9.4), Aphanizomenon flos-aquae (L.) Ralfs (рис. 9.5) и др. (см. раздел 6.5). Разрабатываются способы массового (промышленного) культивирования некоторых видов Cyanophyta, например Spirulina platensis [602].
* (В Японии, например, с целью получения "живого удобрения" в полупроизводственных условиях выращивают Tolypothrix tenuis Kütz. Во Вьетнаме для удобрения полей издавна используют водный папоротник Azolla, в симбиозе с которым живет Anabaena azollae Strasb.)
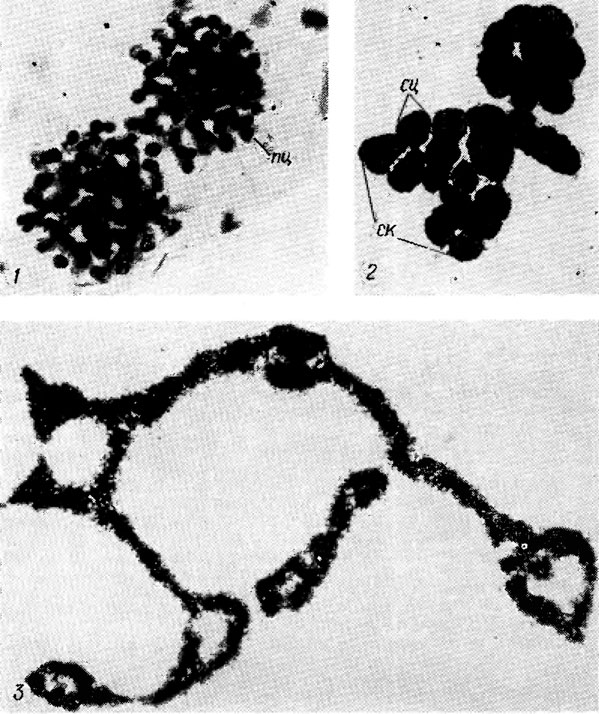
Рис. 9.3. Microcystis aeruginosa Kütz. em. Elenk. f. aeruginosa: 1 - st. simplex; 2 - st. viridis; 3 - st. seriptus (пц - 'простой' одноклеточный цианоид, ск - сложная колония; сц - сложный цианоид)

Рис. 9.4. Anabaena flos-aquae Brob.: 1 - общий вид колонии (клубочка); 2 - отдельная нить (вк - вегетативная клетка; г - гетероциста; мс - молодая спора; с - спора (акинета))
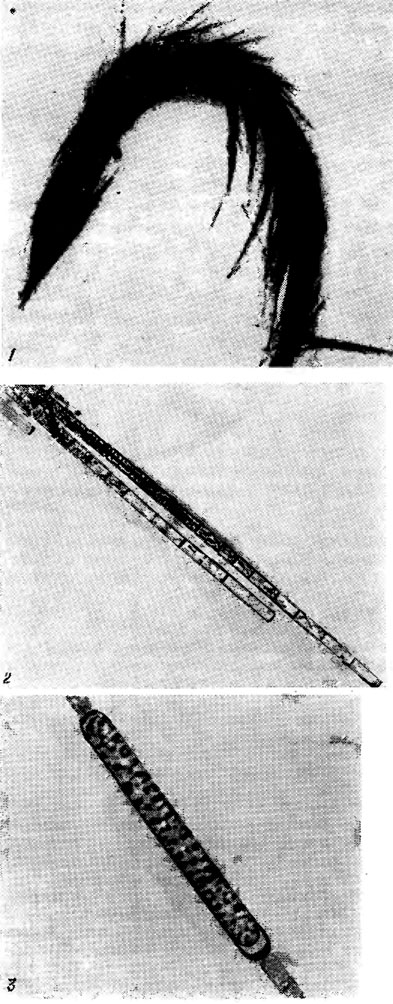
Рис. 9.5. Aphanizomenon flos-aquae (L.) Ralfs: 1 - общий вид колонии (пучка нитей); 2 - концы трихомов; 3 - акинета
В заключение подчеркнем, что в связи с теоретической и практической значимостью исследования синезеленых водорослей, а также их существенной ролью в жизни естественных экосистем, рассматриваемая группа организмов издавна привлекала внимание исследователей. Уже опубликованы многие обобщающие работы, определительные пособия, сборники и монографии, специально посвященные Cyanophyta, причем не только флористикосистематические и морфологические [42, вып. 1; 112; 162; 239, вып. 2; 388; 430-432; 651а и др.], но также (главным образом во второй половине XX ст.) посвященные другим, как теоретическим, так и прикладным вопросам [26, 89, 91, 97, 169, 289, 290, 319, 412, 673, 674 и др.].
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'