Глава V. Харовые водоросли (Charophyta) (И. М. Шайкин)
Методы изучения, принципы классификации, основные черты палеоэкологии, филогения и стратиграфическое значение
Харовые водоросли (харофиты, лучницы) - своеобразные растения, резко отличающиеся от остальных водорослей обликом слоевища и органов размножения. Они известны как из современных пресноводных и засолоненных водоемов, так и в ископаемом состоянии от силурийского до четвертичного периодов.
Морфологии, систематике, палеоэкологии, распространению харофитов посвящены сотни публикаций ботаников и палеонтологов многих стран (СССР, Великобритании, Франции, Швеции, США, КНР, Румынии, Польши, Италии, ФРГ, Аргентины и др.). Описаны 6 родов и 300 видов современных харовых водорослей со всех континентов, свыше 110 родов и 600 видов ископаемых представителей всех трех порядков типа Charophyta.
Современные харовые водоросли - сложные многоклеточные слоевища (талломы). Это кустистоветвящиеся ните- или стеблевидные зеленые побеги членисто-мутовчатого строения, укореняющиеся на дне водоемов с помощью многочисленных тонких бесцветных ризоидов (рис. V. 1). Слоевище очень похоже на некоторые высшие растения (хвощи, роголистник), поэтому основные ветвящиеся побеги условно названы "стеблями", а расположенные мутовками боковые побеги - "листьями". Места ответвления листьев называются узлами, а участки стебля между ними - междоузлиями /Голлербах, 1977; Голлербах, Красавина, 1983/.
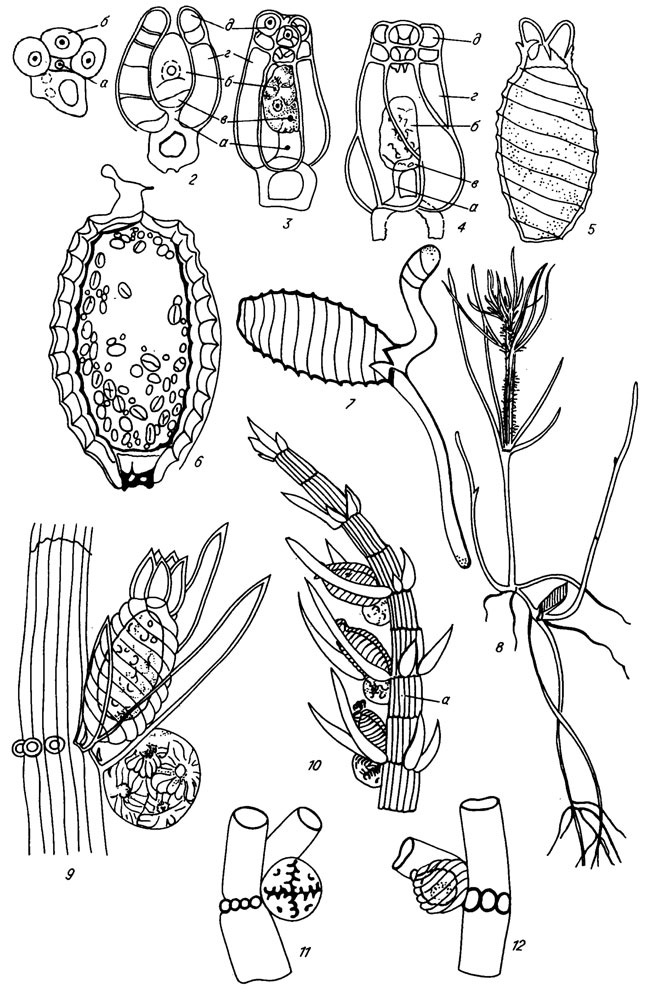
Рис. V. 1. Современные харовые водоросли. 1-8 - последовательные стадии образования водоросли: 1-4 - развитие спорофидия (а - клетка листового узла, б - яйцеклетка, в - поворотные клетки, г - обволакивающие клетки, д - клетки коронки), 5, 7, 8 - прорастание ооспорангия, 6 - строение ооспорангия; 9, 10 - расположение спорангия и антеридия у однодомных видов (а - кора таллома (по М. М. Голлербаху /1977/; М. М. Голлербаху, Л. K. Красавиной /1983/; В. П. Маслову /1963 а, б/; В. Крофту /Croft, 1952/); 11-12 - расположение антеридия и спорофидия у двудомных видов
Размножение у харофитов вегетативное и половое. Органы полового размножения (женские - спорофидии, мужские - антеридии) развиваются на листовых узлах (см. рис. V. 1) либо вместе на одном талломе у однодомных видов, либо раздельно на разных талломах у двудомных видов.
Зрелый спорофидии состоит из оогония, оболочки (споростегия) и одной-двух базальных клеток (см. рис. V. 1). В шаровидном теле антеридия сложным путем развиваются сперматозоиды, которые благодаря водной среде попадают через отверстие в центре спорофидия в оогоний. После оплодотворения яйцеклетки оогония образуется ооспорангий. Он состоит из ооспоры и оболочки-спорангиодерма /Horn af Rantzien, 1959 а; Маслов, 1963 а/. Ооспора - преобразованная яйцеклетка, стенка которой имеет двухслойные оболочки - спорины (мембраны); спорангиодерм - отвердевшие спиральные клетки, внешний слой которых обызвествляется, образуя кальцитовую оболочку (кальцину). В основании ооспорангия бывшие базальные клетки, обызвестившись, превращаются в базальную пластинку (см. рис. V. 1).
После периода покоя оосорангии прорастают. Прорастание начинается с исчезновения зерен крахмала и жира, затем происходят замена их зернистой протоплазмой, деление ядра и центральной клетки, от которой берет начало новое растение. Одновременно с внутренними изменениями оболочки ооспорангия на вершине расщепляются на пять зубцов, давая выход проростку, возникающему из центральной клетки (см. рис. V. 1). До этого путем деления центральной клетки происходит образование двух функционально различных клеток: более крупной, дающей начало предростку, из которого вырастает стеблевой побег, и меньшей - дающей начало первому ризоиду (см. рис. V. 1). В результате ряда клеточных делений возникают две однорядные нити, которые дифференцируются на узлы и междоузлия. Дальнейший рост - верхушечный. Из узлов предростка образуются вторичные предростки, мутовки листьев и боковые ветви стебля, из узлов первого ризоида - вторичные ризоиды и их мутовчатые волоски. Таким путем формируется слоевище современной харовой водоросли, состоящей из нескольких стеблевых побегов в верхней части и нескольких сложных ризоидов в нижней /Голлербах, 1977/.
Обызвествляются отдельные органы у современных и, очевидно, ископаемых харофитов по-разному /Horn af Rantzien, 1959 b; Маслов, 1963 а/. У вегетативных частей происходит так называемое "физиологическое отложение извести", т. е. выпадение извести на поверхности клеток в результате фотосинтетической деятельности растения в водной среде. В спиральных и узловой (базальной) клетках женского органа размножения осуществляется так называемое "органическое отложение извести", т. е. выделение извести из клеточного сока, в котором кальций находится в виде органического соединения, У современных харофитов развитие Известковой оболочки начинается только с момента оплодотворения яйцеклетки и продолжается в течение последующей жизни растения, но этот процесс протекает довольно быстро.
В ископаемом состоянии целые слоевища сохраняются редко. Чаще встречаются их обломки. Главным образом ископаемые харофиты представлены в виде обызвествленных остатков ооспорангия - кальцины, которую принято называть гирогонитом. В основании гирогонита имеется отверстие, закрытое изнутри базальной пластинкой (или пластинками).
У некоторых семейств харофитов гирогониты покрыты вторичной известковой оболочкой - утрикулом, образовавшимся за счет срастания обызвествившихся "веточек" и "листьев" таллома (рис. V. 2, V. 3).

Рис. V. 2. Утрикулы и гирогониты харофитов. 1, 2 - хованелла сбоку и сверху; 3, 4 - сицидиум сбоку и сверху; 5 - трохилиск; 6, 7 - эохара сбоку и сверху; 8, 9 - стомохара сбоку и сверху; 10, 11 - палеохара сбоку и сверху; 12-14 - рашкиелла сбоку и сверху с 'крышечкой', сверху без 'крышечки'; 15 - харацея сверху после выхода плода, 16 - харацея, вершина (по В. П. Маслову /1963/, К. Гуревич /1968/, Р. Пек / Реск,1963 а/)

Рис. V. 3. Утрикулы и гирогониты. 1, 2 - утрикулы умбелл в узлах таллома; 3, 4 - утрикулы Pinnoputamen; 5-9 - гирогониты харацей лампротамноидного типа: 5, 6 - Lamprothamnium; 7-9 - схемы вершин сбоку; 10-13 - гирогониты харацей псилохароидного типа; 10, 11 - Saidakovskyella, 12 - Hornichara, 13 - Grambastichara; 14, 15 - Tectochara - гирогонит харацей нителлопсидоидного типа; 16-18 - Tolypella (современная) - гирогонит толипеллоидного типа (по Б. Й. Чувашову /1973/, Ц. Ванг, Дж, Лу/ Wang, Lu, 1980/, Хорн аф Рантциен /Horn af Rantzien, 1959 a, b/)
Вегетативные части ископаемых харофитов еще недостаточно освещены в литературе, методика их изучения не разработана, а имеющиеся описания основаны на методике описания современных харофитов /Harris, 1939; Peck, 1937, 1957; Madler, 1952/. Как известно, современные харофиты относятся лишь к одному семейству Characeae, появившемуся в середине мезозойской эры, а некоторые находки свидетельствуют о том, что остатки слоевищ происходят из более древних эпох и принадлежат к другим семействам /Ищенко, Сайдаковский, 1975; Т. А. и А. А. Ищенко, 1982/.
Отсюда следует, что на данном этапе изучения ископаемых харофитов необходимо накапливать фактический материал, опираясь на имеющиеся знания о современных и ископаемых харовых водорослях. Поэтому при находках вегетативных частей ископаемых харофитов необходимо приводить тщательные описания и изображения как внешних признаков, так и внутренней структуры по изломам и шлифам.
Описание вегетативных частей харофитов включает морфологию растения, его размеры (длина общая и отдельных частей), характер строения узлов и междоузлий, количество и размеры стеблей и боковых побегов - листьев, на изломах - количество и диаметр центральной и кортикальных трубок. В шлифах описывается микроструктура стенок вегетативных частей (зернистость, цвет). Устанавливается наличие на талломе гирогонитов, утрикулов.
Общепринятая методика изучения гирогонитов и утрикулов базируется на классических работах /Lyell, 1832; Unger, 1850; Карпинский, 1906; Reid, Groves, 1916; Harris, 1939; Peck, 1934 a, b, 1937,1938, 1946, 1953, 1957; Rasky, 1945; Маслов, 1947, 1950, 1956 a, б, 1960 a, б, 1961 a, б, 1962, 1963 a, б, 1965, 1966; Mädler, 1952, 1953, 1955, 1957; Horn af Rantzien, 1953, 1959 a, b; Grambast, 1956, 1962, 1966, 1973, 1975/. Советские и многие зарубежные палеонтологи придерживаются номенклатуры и методики изучения гирогонитов и утрикулов, изложенных В. П. Масловым в 1963-1966 гг. Гирогониты и утрикулы изучаются по внешним признакам наружной поверхности и внутренним, видимым в шлифах на разрушенных экземплярах (рис. V. 4).
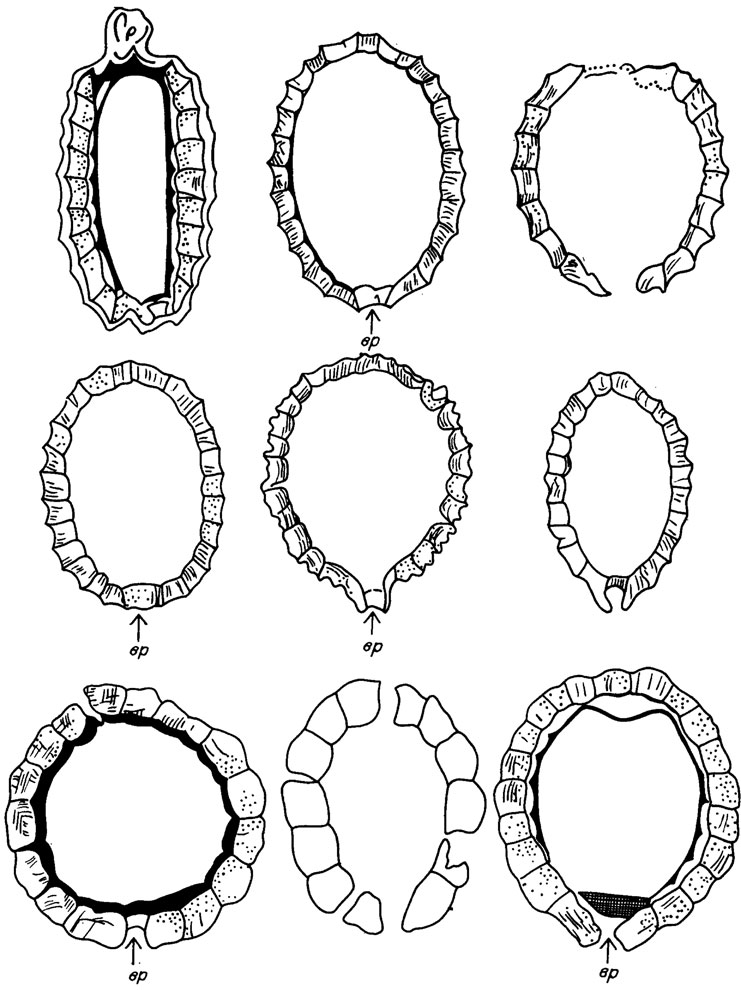
Рис. V. 4. Осевые сечения гирогонитов, вр - базальная пластинка (по В. П. Маслову /1963 а, б/)
На гирогонитах замеряются длина и ширина, количество оборотов видимых сбоку партекальцин, их ширина, экваториальный угол, диаметр апикальной и базальной поры, высота и диаметр апикального выступа (шейки). По данным замеров делается расчет отношения длины гирогонита к его ширине, изополярного и анизополярного индекса /Horn af Rantzien, 1959 а/.
Внутренние признаки гирогонитов в шлифах: микроструктура оболочки (зернистость, цвет), ее толщина, строение (слоистость) вершины, основания полости гирогонита, наличие ооспоровой мембраны.
Аналогично гирогонитам изучаются внешние и внутренние признаки утрикулов. При этом особое внимание уделяется характеру скульптуры поверхности утрикула (орнаментации), наличию дополнительных отверстий, узелков и т. д.
Приведенные морфологические и микроструктурные признаки положены в основу современной систематики ископаемых харофитов. Харофиты диагностируются по следующим систематическим признакам.
1. Семейство и более высокие таксоны: по наличию или отсутствию утрикула, у утрикулов - по форме (симметрии) и характеру орнаментации; у гирогонитов - по направлению завивания партекальцин (правое или левое) и их количеству, строению вершины.
2. Род: у утрикулов - по форме (симметрии), характеру орнаментации, дополнительным отверстиям, у гирогонитов - по строению вершины, форме гирогонита, количеству партекальцин.
3. Вид: у утрикулов - по форме, размерам, характеру орнаментации, строению вершины и основания, у гирогонитов - по форме и размерам, форме вершины и основания, количеству видимых сбоку партекальцин, их форме, размерам и форме вершинной поры и мыса, размерам и форме базальной поры, отношению длины к ширине, изополярному и анизополярному индексам, микроструктуре оболочки в шлифе, наличию и форме базальной пластинки.
В случае нахождения целых слоевищ или их частей вместе с гирогонитами или утрикулами последние являются определяющими при диагностике ископаемых харофитов.
Систематика современных харофитов основывается главным образом на морфологии слоевища и лишь незначительно учитывает особенности женского органа размножения /Голлербах, 1977; Голлербах, Красавина, 1983/. Как показал Дж. Хорн аф Рантциен /Horn af Rantzien, 1959/ в специальном исследовании современных харофитов, она возможна и на основе изучения ооспорангия и его известковой оболочки. Поскольку ископаемые вегетативные органы харофитов встречаются редко либо вовсе не сохраняются, формально классификацию этих водорослей по остаткам женского органа размножения нельзя было бы проводить. Однако можно утверждать, что наличие вегетативных частей водорослей вряд ли изменило бы что-нибудь. Почему? Потому, что, по данным многолетних исследований, наиболее изменчивыми решающими систематическими признаками у харофитов обладает женский Орган размножения (либо его остатки у ископаемых). Это и определяет необходимость устранения формальных причин для создания систематики ископаемых харофитов не по вегетативным органам, а по обызвествленным оболочкам женского органа размножения. Такая классификация в значительной степени искусственна, но она оправдала себя в практике геологического изучения осадочных толщ земной коры.
Направление завивания партекальцин, их количество и характер смыкания на вершине гирогонита, а также наличие утрикула уверенно служат для определения порядков и семейств ископаемых харофитов. Наиболее выдающимся критерием для выделения таксонов самого высокого ранга является направление завивания основных элементов известковой оболочки, главным образом у гирогонитов. По этому критерию выделены три порядка: Sycidiales, с вертикально расположенными основными элементами оболочки; Trochiliscales, с гирогонитами, имеющими правое завивание партекальцин (по часовой стрелке); Charales, с гирогонитами, имеющими левое завивание партекальцин (против часовой стрелки),
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'