Глава III. Зеленые водоросли (Chlqrophyta) (В. П. Шуйский)
Принципы систематики и методы изучения зеленых водорослей
Отдел Chlorophyta по числу ныне живущих видов (более 13 тысяч) занимает первое место среди водорослей, но это, вероятно, всего лишь реликты былого разнообразия, поскольку, по единодушному мнению альгологов, пик своего расцвета зеленые водоросли прошли еще в позднем палеозое и мезозое /Жизнь растений, 1977; Виноградова и др., 1980; Курс..., 1981/.
Название отдела связано с наличием в клетках современных его представителей травяно-зеленых хлоропластов, в которых хлорофилл преобладает над каротиноидами. Окраска водорослей не всегда чисто зеленая" Она нередко затушевывается дополнительными пигментами, в особенности красным гематохромом.
Питание зеленых водорослей обычно фотоавтотрофное, но встречаются миксотрофия и гетеротрофия. Среди зеленых водорослей известны симбионты беспозвоночных животных и паразиты. В качестве запасных питательных веществ накапливаются крахмал, а также масла. По наличию хлорофилла и крахмала зеленые водоросли сходны с высшими растениями и поэтому считаются наиболее вероятными их родоначальниками. Репродуктивные системы зеленых водорослей отличаются большой динамичностью и реактивностью к внешним условиям среды. Одинаково широко распространено вегетативное, бесполое и половое размножение.
Вегетативное размножение в простом случае осуществляется путем фрагментации слоевища и последующего дорастания отдельных кусков до размеров материнского растения, более сложный способ - посредством акинет (модифицированных клеток с толстой оболочкой и запасом питательных веществ). Еще один способ заключается в образовании так называемых выводковых почек - особых выростов слоевища, которые отделяются от основного вегетативного тела перегородкой, а после отпадения от него прорастают в новое растение.
Бесполое размножение происходит с помощью подвижных, снабженных жгутиками зооспор или неподвижных апланоспор. Споры созревают в клетках слоевища или в специальных органах бесполого размножения - спорангиях. По месту образования спор различаются эндо-, кладо- и хористоспоровый способы размножения /Кордэ, Максимова, 1980; Основы палеонтологии. 1963; Elliot, 1972/. Наиболее развитым считается последний из перечисленных способов, но все они несомненно существуют с палеозойского времени.
Наибольшим разнообразием отличаются способы полового размножения. Различают изо-, анизо- и оогамию. При изогамии копулируют с последующим образованием зиготы равновеликие подвижные изогаметы, сходные с зооспорами. Слияние гамет различной величины называется анизогамией, или гетерогамией. Иногда имеет место типичная оогамия - слияние крупной женской яйцеклетки и мужской гаметы. Формирование гамет, как и спор, происходит в осевой части слоевища (эндоспоровый тип), в ответвлениях (кладоспоровый тип) или в особых органах - гаметангиях, сходных со спорангиями (хористоспоровый тип) (рис. III. 1). В результате слияния двух гамет образуется зигота (ооспора) - особая клетка с толстой оболочкой и запасом питательных веществ. После ряда промежуточных превращений зигота прорастает во взрослое растение.
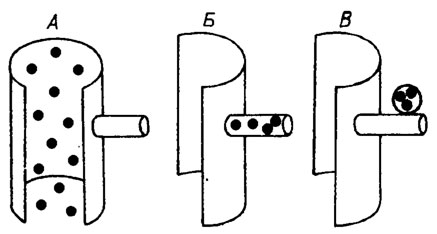
Рис. III. 1. Схематическое изображение эндоспорового (А), кладоспорового (Б) и хористоспорового (В) способов полового и бесполого размножения
У многих зеленых водорослей наблюдается чередование полового и бесполого поколений. В первом случае спора бесполого размножения прорастает в гаметафит - растение, на котором формируются гаметангии, а зигота - в спорофит. Если гаметафит и спорофит морфологически не различаются, то такая смена поколений называется изоморфной. При гетероморфной смене поколений гаметофит и спорофит могут сильно отличаться друг от друга. Их нередко принимают за различные растения и описывают под разными родовыми названиями. Например, установлено, что современный род Chlorochytrium - спорофитная стадия рода Acrosiphoma, а род Codiolum - спорофит родов Urospora и Spongomorpha /Зинова, 1967, с. 66/.
Способность водорослей сохраняться в осадочных породах самым существенным образом зависит от механических и химических особенностей клеточных оболочек. Естественно, что так называемые "голые" вегетативные клетки, а также споры и гаметы, окруженные только цитоплазматической мембраной - плазмалеммой, имеют мало шансов перейти в ископаемое состояние с сохранением черт первичной морфологии. Но, как показывают электронно-микроскопические исследования, оболочки имеют сложную слоистую структуру и обладают достаточно большой механической прочностью /Жизнь растений, 1977; Курс..., 1981/.
Каждый отдельный слой в разрезе оболочки имеет двухкомпонентное строение: он состоит из аморфного матрикса - стромы, образованной гемицеллюлозами и пектином, а также закономерно ориентированных целлюлозных фибрилл, которые играют роль главного опорного элемента. Отсутствие или слабое развитие целлюлозных фибрилл делает оболочку гомогенной, состоящей преимущественно из пектиновых веществ. В этом случае ее механическая прочность увеличивается, как, например, у сифоновых водорослей (за счет пропитывания известью). В альгологии этот процесс именуется импрегнацией, или инкрустацией.
Биохимия обызвествления слоевища, к сожалению, практически не изучена. Все большее внимание в последнее время привлекает к себе явление биоминерализации, но оно рассматривается пока только в самых общих чертах /Барсков, 1984; Лоуэнстам, 1984/. Выявлены два принципиально различных механизма минерализации, свойственные как животным, так и растениям. Это механизмы биологически индуцируемой и матрицируемой минерализации /Лоуэнстам, 1984/.
При индуцируемой биоминерализации происходят простые химические реакции между конечными продуктами метаболизма и катионами, заимствованными из окружающей водной среды. В результате этих реакций в клеточных оболочках образуются карбонатные минералы в виде кристаллических индивидов различного размера и формы без признаков упорядоченности в агрегатах. Морфологически эти минеральные новообразования мало отличаются от компонентов осадка или цементирующей массы. По этому типу, вероятно, развивается обызвествление большинства сифоновых водорослей, для которых характерны равномерно- или неравномерно-зернистое строение оболочек и относительно небольшая устойчивость при перекристаллизации.
При минерализации с помощью органических матриц зарождение и последующий рост минералов происходят как бы по регламентированной программе под постоянным биологическим контролем и в известной степени независимо от условий окружающей среды. Элементы органических матриц, которые предопределяют состав и кристаллографическую форму минералов, формируются в особом клеточном органе - аппарате Гольджи. Матрицируемый способ биоминерализации особенно широко развит у животных, но он отмечается у самых различных по уровню организации представителей органического мира, в частности даже у прокариот - цианобактерий. Не исключено, что он встречается и у зеленых водорослей.
Матрицируемая биоминерализация может быть простой клеточной и более сложной" - тканевой /Барсков, 1975, 1984/. В любом случае для матрицированных минералов характерен специфический облик и небольшой диапазон размеров кристаллических индивидов. При клеточной минерализации образуются агрегаты изометричных кристаллов или минеральных прутьев - спикул. Возможно, к этому типу относятся ультрамикроскопические структуры кальцита некоторых обызвествленных водорослей, например Calcipholium, Litanaella, а также Renalcis, Girvanella, афросальпингат, археоциат и др. Интересно, что скелетные минеральные образования данной разновидности обнаруживают повышенную устойчивость при перекристаллизации. Это может быть связано с "консервирующим" влиянием остаточного материала органических матриц. Тканевый тип биоминерализации для зеленых водорослей, по-видимому, не характерен и может проявляться только в редких случаях, в частности у таких проблематичных форм, как Fasciella Ivanova (= Shartymophycus Kulik).
Кроме минеральной импрегнации клеточных стенок, известна так называемая адкрустация - выделение из протопласта через поры на поверхность клетки особых веществ, например кутина, а также разного рода слизей пектинового или целлюлозного состава, выполняющих защитную функцию. Ослизнение слоевища у зеленых водорослей может быть постоянным (у примитивных форм это пальмеллоидный тип структуры), временным или вообще отсутствовать, В ослизняющейся части слоевища, по-видимому, могут происходить процессы индуцированной биоминерализации, но на ископаемом материале этот факт однозначно подтвердить довольно трудно, поскольку образовавшийся таким способом, кальцит не отличается от диагенетического. Отложение извести не в оболочках, а на поверхности слоевища отмечается даже у современных пресноводных зеленых водорослей рода Vaucheria /Виноградова и др., 1980, с. 99/.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'